1. Introduction
Thermophilic bacterial proteins and enzymes are attractive to the academic sector and for industrial applications [
1,
2]. Thermophilic proteins and enzymes display high thermal stability and significant activity at elevated temperatures. The high stability and activity of thermophilic proteins at elevated temperatures enable conducting experiments and observing reactions that do not occur at room temperature (e.g., polymerase chain reaction). The molecular mechanism of thermostability has been intensively investigated [
3,
4,
5,
6,
7]. In quantitative terms, it is expressed by a protein stability curve that describes how the free energy of the denaturation, Δ
G, is dependent on temperature and how the denaturation enthalpy and entropy change over an extensive temperature range. In the case of proteins, denaturation enthalpy is strongly temperature-dependent due to the high heat capacity change in the process. A significant positive change in heat capacity has an interesting consequence for the depiction of protein stability as a curve, which adopts a parabolic shape. Hence, two different denaturation processes can be observed: one on the hot side and one on the cold side—so-called cold denaturation. The molecular origins of cold denaturation lie in the temperature-dependent hydration of hydrophobic groups, which becomes more favorable at low temperatures [
8]. Cold denaturation is not often observed, mainly due to two interrelated reasons. First, cold denaturation usually occurs at temperatures well below the experimental window of spectroscopic/calorimetric techniques. To observe cold denaturation at ambient temperatures, the protein stability curve, Δ
G vs.
T, should be shifted horizontally to higher temperatures, indicating a higher hot denaturation transition temperature. Thus, thermophilic proteins can be good candidates for observing cold denaturation.
However, the presence of residual structure in the unfolded state may decrease heat capacity [
9,
10] and, hence, reduce the curvature of the protein stability curve, which then shifts the threshold for cold denaturation to temperatures that are not readily achievable. Second, the changes in heat capacity are often not sufficiently high to allow observation of cold denaturation. As heat capacity changes are correlated with the number of residues [
11] or, more precisely, with the size of the cooperative unit, cold denaturation should be reversible and readily detectable for larger proteins. On the other hand, larger proteins are often arranged in domains (beads on a string or discontinuous domains) that may independently fold/unfold, diminishing the size of the cooperative unit and, hence, the magnitude of the heat capacity change. Taken together, there are only limited cases under which cold denaturation can be observed.
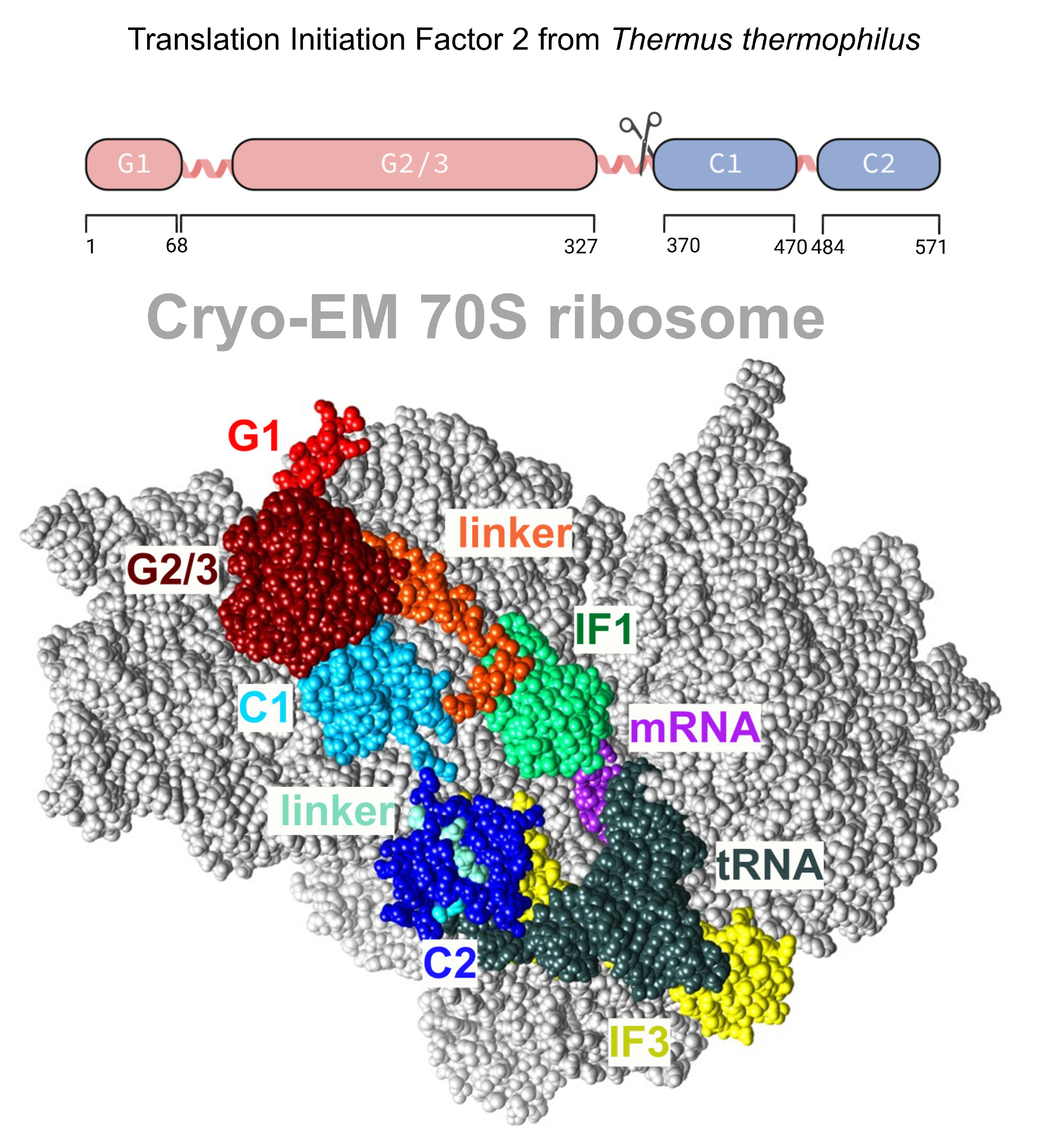
Translation initiation factor 2 (IF2,
Figure 1) is among the largest nonribosomal GTPase involved in initiating protein biosynthesis in the cell. It belongs to the family of structurally related enzymes catalyzing GTP hydrolysis during the early processes of mRNA translation. IF2 plays a central role in translation initiation via the specific selection of fMet-tRNA
fMet. IF2 specific selection is based on recognition of the N-formyl-methionine and aminoacyl acceptor stem of fMet-tRNA
fMet [
12,
13] and significantly faster association of the 50S subunit and correctly assembled 30S complex, which contains all functional components for successful translation initiation. IF2 also participates in other cellular processes, such as transcription [
14], the protein export reaction [
15], and the prevention of protein aggregation [
16], and it functions as a cellular metabolic sensor that toggles between an active, GTP-bound form under normal conditions and an inactive form with bound ppGpp—a nucleotide messenger produced in bacteria during their starvation [
17].
For successful translation initiation and recovery, IF2 interacts with many biological partners and undergoes functional cycling, including GTP hydrolysis, GDP/GTP exchange, initiator tRNA binding, and binding to the 30S ribosome subunit. While these activities are facilitated by different domains of IF2, they all act in an orchestrated, sequential manner that is indicative of allosteric-like domain–domain communication at individual interaction stages. Our understanding of the function–structure relationship has been largely improved by structural data. First, the full-length three-dimensional structure of IF2 was resolved for IF2/eIF5B from
M. thermoautotrophicum [
18]. This remarkable structure displays a novel and unusual chalice-like protein fold with two domains separated by a 4 nm long α-helix. This fold possesses some common properties with the domain architectures of EF-Tu and EF-G, suggesting that all three proteins may interact similarly.
Interestingly, the structure of IF2 does not explain how the signal of GTP hydrolysis in the G-domain is propagated to the distal C2 domain. In 2005, a cryo-EM model of ribosome-bound IF2 from
E. coli was published [
19], in which IF2 undergoes conformational rearrangement upon binding to the ribosome. In particular, the authors observed that the portion attributable to the GTP-binding domain (G domain) of IF2 was significantly larger than the G domain of the reoriented crystal structure [
19]. Hence, IF2 undergoes large conformational changes during translation initiation. In 2016, 11 cryo-EM structures of the
Thermus thermophilus translation preinitiation complex were published [
12]. The complex contains 30S–IF1–IF2–IF3–mRNA–tRNA. Again, the authors observed that the conformation of ribosome-bound IF2 differs from the crystal structure of
Thermus IF2 [
20]. Similar conformational rearrangements of IF2 were also reported in recent cryo-EM structures of the 70S initiation complex from
Pseudomonas aeruginosa, with a bound compact form of IF2–GDP and initiator tRNA [
21]. NMR and circular dichroism of
E. coli IF2 revealed a flexible linker connecting the N-terminal and central segments of the protein with the C-terminal domain [
22]. Thus, multidomain IF2 may be considered flexible, with the capacity to undergo dynamic conformational transitions. Interestingly, the structure of release factor 2, another translation factor, also exhibits differences between its free form in solution compared with the conformation of its ribosome-bound form [
23,
24,
25,
26].
The X-ray crystal structures of full-length
Thermus thermophilus apo IF2 and its complex with GDP have been obtained at 3.1 Å resolution [
20,
27]. The structure of IF2 bears a resemblance to beads on a string, rather than exhibiting a chalice shape similar to eIF5B. IF2 adopts a stable and ordered conformation when interacting with the ribosome, which is facilitated by the predetermined inherent flexibility and expandability of the protein (
Figure 1). Possible elements of flexibility are long helices. The overall dimensions of IF2 depend on the nucleotide status of IF2. The apoform of
T. thermophilus IF2 is more extended, while the GDP-bound IF2 appears more compact. However, IF2 is significantly larger when in a functionally active state, bound to the 30S ribosomal subunit. Unlike all other translational GTPases, IF2 does not have an effecter domain that stably contacts with the switch II region of the GTPase domain. In summary, cryo-EM reconstructions, NMR experiments, and the 3D structure in solution have shown that IF2 transitions from being flexible in solution to having an extended conformation with disrupted interdomain contacts when interacting with ribosomal complexes.
In our previous work, we analyzed the thermodynamic properties of the reversible denaturation of IF2 along with its isolated G and C fragments [
28]. For the purpose of clarity, we used the term “fragments” for proteins obtained upon proteolytical cleavage of IF2. Previously, these fragments were called domains, which is inaccurate based on the structural information that is now available. Circular dichroism in the far-UV region has shown that IF2 and its fragments are well folded. Sedimentation velocity experiments reveal that full-length IF2, the G fragment, and most of the C fragment population are fully monomeric.
Thermal denaturation of multidomain IF2 wt occurs at a high melting temperature of 94.5 °C and displays near-complete reversibility. IF2 can be stabilized by GDP and its analog GTP, as indicated by the higher transition temperatures. The high reversibility of thermal denaturation of such a large (63 kDa) protein is remarkable. This might be a possible consequence of the highly modular multidomain structure of IF2. Thermal denaturation is also found to be highly reversible for the G fragment but not the C fragment. The C fragment of IF2 vividly and irreversibly aggregates upon heating at temperatures above the melting temperature. This adverse aggregation can be suppressed using a denaturing agent, which also restores the high reversibility of thermal denaturation. A similar effect of GndHCl on the reversibility of thermal transition was observed for the
B. stearothermophilus C fragment [
29]. The isothermal unfolding experiment revealed an unusually small m-value (m = d ΔG/d[denaturant]), possibly due to unfolding intermediate(s) and because of independent, spectroscopically overlapping unfolding transitions of the G and C fragments. The observed effects follow the Hofmeister series of salts or others. Here, we evaluate the effect of salts on the cold denaturation of thermophilic multidomain initiation factor 2 in the hope of determining the dominant contributions to cold denaturation and to the stability of IF2 and its domains. A possible major contributor could be the screening of electrostatic interactions of IF2 domains by salts (lower dependence on salt type) or nonelectrostatic components and dispersion forces (strongly dependent on salt type). By comparing the effect of chloride salts of the cations Li
+, Na
+, NH
4+, and Cs
+ and sodium perchlorate on thermodynamic values, we found that the suppression of cold denaturation is highly salt-specific.
2. Results
2.1. Cold and Hot Denaturation Processes of IF2 Are Sensitive to Salts
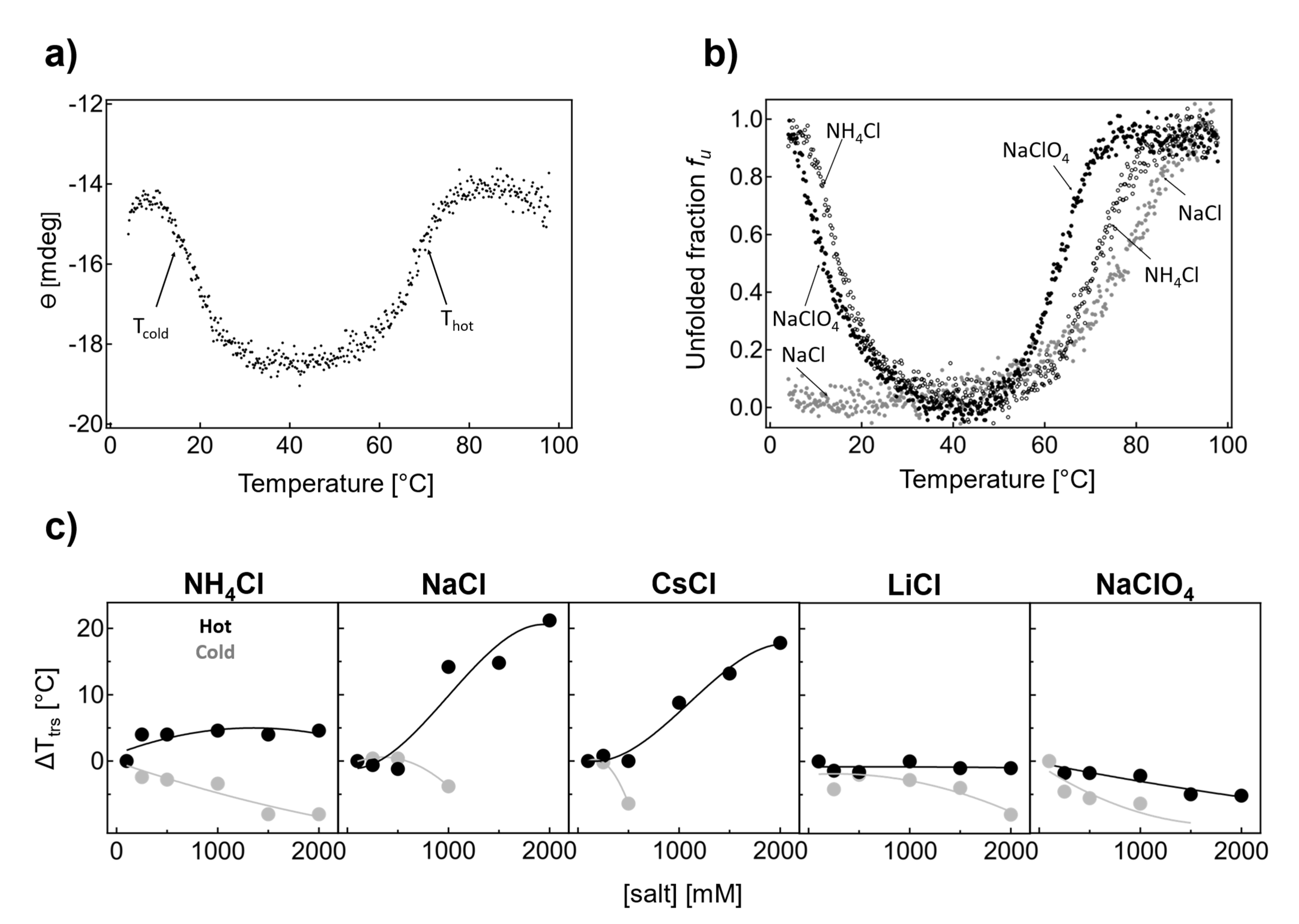
Circular dichroism in the far-UV region is a highly sensitive technique for studying the conformational stability of proteins. This is due to the strong absorption of the peptide group and the high sensitivity of electron transitions n→π* (at 222 nm) and π→π * (at 208 nm) to the protein secondary structure [
30]. Circular dichroism at 222 nm was used to monitor the thermal unfolding of IF2 in the presence of 0.1 M NaCl and a phosphate buffer (
Figure 2a). At the initial stages of thermal denaturation at 4 °C, the ellipticity was observed to have a sigmoidal dependence on temperature. Then, at 30 °C, the ellipticity remained constant until the temperature reached 60 °C and further increased; again, a sigmoidal-like unfolding transition could be observed. We attributed both sigmoidal processes to cold and hot denaturation (see labels
Ttrs, cold and
Ttrs, hot in
Figure 2a). Hence, at ambient temperatures (ca. 20 °C), the thermophilic multidomain protein IF2 was largely unfolded (ca. 50%). To obtain the transition midpoint temperatures in a model-independent way, we applied an approach based on the first derivative of the experimental curves. Upon derivation, two extrema were observed: a minimum, which corresponds to the midpoint of cold denaturation; and a maximum, which corresponds to the midpoint of hot denaturation. Both denaturing transitions were cooperative, and the temperatures corresponding to cold and hot denaturation were
Ttrs, cold 18 ± 0.4 °C and
Ttrs, hot 67.4 ± 0.2 °C.
One of the possible mechanisms underlying the thermal adaptation of thermophilic proteins is based on the optimization of electrostatic interactions on the protein surface. The role of electrostatic interactions in determining protein stability can be studied by examining the dependence of protein stability on the concentration different salt types. To understand the role of electrostatic interactions in the stability of thermophilic IF2, we first conducted experiments using selected salts at a very high ionic strength: 1.5 M (
Figure 2b).
Figure 2b shows the thermal denaturation of IF2 in the presence of 1.5 M NaCl, NH
4Cl, and NaClO
4, plotted as unfolded fraction (
fu) vs. temperature (
T). This combination allows us to compare the role of cations of different ranks in the Hofmeister series (Na
+ versus NH
4+, both chlorides) and different anions (Cl
− versus ClO
4−, Na
+). In the presence of NH
4Cl, only minor shifts in
Thot toward higher temperatures at higher salt concentrations and a slight shift in
Tcold to lower values were observed. In the presence of NaCl, we found that thermal denaturation on the hot side shifted toward higher temperatures at higher salt concentrations. Interestingly, at 1.5 M NaCl, cold denaturation of IF2 was not observed due to a significant shift in the threshold toward lower temperatures that were not achievable under the conditions of the study. The difference between Na
+ and NH
4+ chlorides indicates that the nature of the cation and its rank in the Hofmeister series affect the midpoint of cold denaturation.
NH4+ is a chaotropic cation, while sodium is a mild kosmotropic cation that can stabilize the native conformation of IF2. Such stabilization can be offset by a chaotropic anion. Therefore, in the following unfolding experiment, we used a combination of sodium and chaotropic anion-perchlorate. In the presence of 1.5 M NaClO4, cold denaturation was again observed with Tcold = 11.6 ± 0.2 °C. The temperature for thermal denaturation on the hot side decreased as expected for chaotropic anion, with Ttrs, hot = 63.6 ± 0.4 °C. One can conclude that cold and hot denaturation processes are dependent on the salt type and ionic strength.
We systematically examined the thermal denaturation of IF2 in NH
4Cl, NaCl, CsCl, LiCl, and NaClO
4 in the concentration range of 0.1–2 M (
Figure 2c gray
Ttrs, cold and black
Ttrs, hot). In the presence of NH
4Cl (
Figure 2c), the thermal transition temperature
Ttrs, hot showed a hyperbolic increase. On the other hand,
Ttrs, cold displayed a linear decrease, indicating that IF2 becomes more stable and, hence, more folded at ambient temperature. Here, even though NH
4+ is a chaotropic cation, higher transition midpoint temperatures resulted in the stabilization of the protein at elevated temperatures.
A similar increase in thermal denaturation on the hot side is visible in the presence of NaCl. Here,
Ttrs, hot increased dramatically;
Ttrs, hot increased by ca. 15 °C in the presence of 1 M salts and by 20
°C in 2 M NaCl. In the case of cold denaturation,
Ttrs, cold decreased to a small extent, by 3.8 °C, with 1 M NaCl, as can be seen from
Figure 2c. However, cold denaturation was not observed because the CD signal was approximately halved compared with at 0.1 M NaCl. The possible reasons for this are discussed later in this paper.
Thermal denaturation in the presence of CsCl mirrors the dependence found for NaCl (
Figure 2c). Increasing the CsCl concentration significantly increased the transition temperature. At the same time, the threshold for cold denaturation shifted toward lower temperatures, and the cold denaturation process was not visible anymore above 0.5 M CsCl.
LiCl and NaClO4 salts were found to share similarities in terms of the concentration dependences of Ttrs, hot and Ttrs, cold. Here, Ttrs, hot decreased by 1.0 ± 0.2 °C (2 M LiCl) or 5.2 ± 0.2 °C (2 M NaClO4), which indicates a mild destabilization of native IF2 on the hot side. On the other hand, Ttrs, cold decreased by 8.0 ± 0.4 °C (at 2 M LiCl) and 9.8 ± 0.2 °C (at 1.5 M NaClO4), indicating IF2 becomes more stable at ambient temperatures.
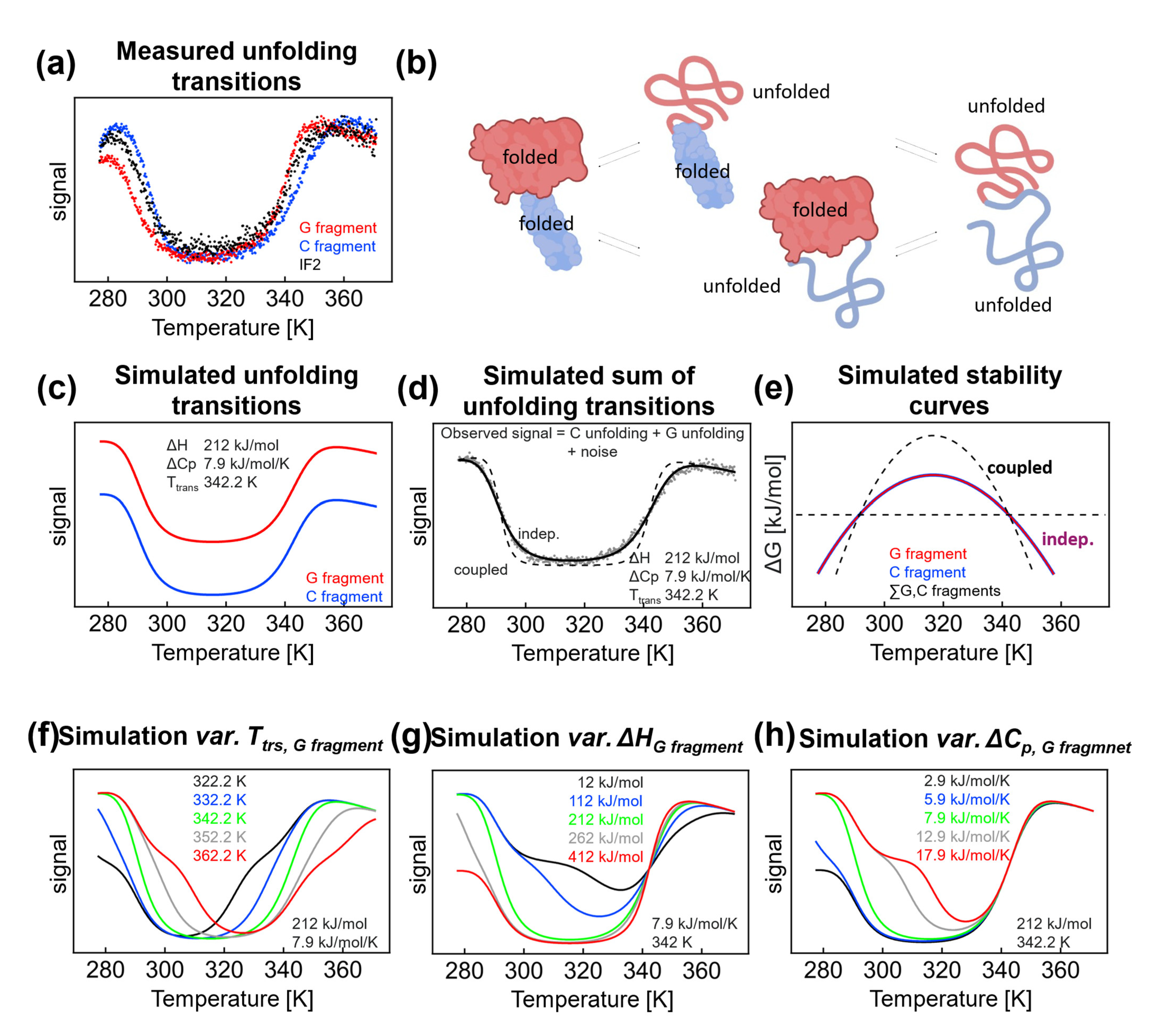
2.2. Limitations to the Analysis of Thermal Denaturation of Two Overlapping Independent Fragments
Thermus thermophilus IF2 consists of two parts that can be obtained after proteolytical cleavage [
31]: a G fragment and a C fragment. Although the fragments have similar stabilities, two different denaturation pathways are represented. On the other hand, when the denaturation of domains is coupled, it can be described as a two-state process. For fragments displaying similar signals (as in our case,
Figure 3a, similar secondary structure content) and independent denaturation, analysis of the thermal denaturation profiles can be misleading. We previously showed [
28] that both fragments display similar thermal denaturation profiles; in the presence of 9 M urea,
Ttrs, hot for the G fragment is 67.6 °C and
Ttrs, hot for the C fragment is 71.6 °C. Hence, we assumed there are two denaturation pathways (
Figure 3b). In one pathway, the denaturation starts from the C fragment, and in the second pathway, the unfolding starts from the G fragment. The observed denaturation process would correspond to the average signal, and the enthalpy and heat capacity would be much lower compared with the coupled all-or-nothing denaturation, where enthalpies and heat capacities add up. We conducted a thermodynamic analysis of IF2 in the presence of 0.1 M NaCl and found values that were too low, as was expected for such a large multidomain protein. To highlight the observed inconsistencies, the thermal profile of the CD signal of IF2 in 0.1 M NaCl was fitted by the Gibbs–Helmholtz equation with the parameters Δ
cp 7.9 kJ/mol/K, Δ
H 212 kJ/mol, and
Ttrs, hot 69.1 °C, i.e., with a heat capacity value that is significantly smaller than expected for a 563 aa protein (7.9 vs. expected 33 kJ/K/mol) [
11]. We assumed that the experimental curve could be decomposed according to independent unfolding of the G and C fragments, in which transitions are described by the same thermodynamic parameters (
Figure 3c). Hence, the observed thermal profile reflects the sum of the unfolding of the independent fragments, with limited possibility to resolve individual fragment unfolding profiles and their corresponding thermodynamic parameters (
Figure 3d,e).
Mathematical Modeling—Effect of Individual Parameters and Transition Temperature (
Figure 3f): We examined how the thermal profiles of independently unfolding fragments can be resolved and how they affect the apparent two-state character of thermal curves. The denaturation parameters for the C fragment were held constant (Δ
H 212 kJ/mol/K, Δ
cp 7.9 kJ/mol, and
Ttrs, hot 342.2 K) while the parameters for the G fragment were changed as indicated in
Figure 3f (variable
Ttrs, hot, Δ
H 212 kJ/mol/K, Δ
cp 7.9 kJ/mol). A difference of 10 °C in
Ttrs, hot between fragments did not allow resolving individual unfolding transitions. Under the parameters of the study, a difference of at least 20 °C was needed to resolve transitions in both the hot and cold sides. From a technical viewpoint, multistate behavior on the hot side was more evident due to the presence of post- and pretransition baselines. This situation was reversed when
Ttrs, hot shifted by at least 20 °C toward a higher temperature; here, the cold denaturation profile exhibited clear post- and pretransition baselines and, hence, fragment unfolding became visible. To conclude, even with a 15 °C difference between
Ttrs, hot of the fragments was not sufficient to allow distinguishing between individual transitions.
Enthalpy (
Figure 3g): A decrease in the enthalpy of the G fragment would result in an evident fingerprint of multistate unfolding. Importantly, the simulated enthalpies (112 and 12 kJ/mol) were quite low and on the small side of the usual enthalpy range. Moreover, post- and pretransition baselines were not reached due to the low stability of the native form; under any conditions, the native state is not 100% present, which may bias the analysis [
32]. The increasing denaturation enthalpy of the G fragment and high Van ’t Hoff enthalpies do not allow for a clear resolution between the individual denaturation curves.
Heat capacity change (
Figure 3h): In the last simulation set, we changed the values of the heat capacity of the G fragment while keeping all other parameters constant, including the parameters for the C fragment (
Figure 3h). A decrease in the heat capacity from 5.9 to 2.9 kJ/mol/K leads to changes in the cold denaturation profile; however, the changes in the profiles are difficult to analyze using the three-state model. The thermal denaturation was not affected at high temperatures because,
Ttrs, hot on the hot side was taken as constant in our simulation. An increase in the unfolding heat capacity from 12.9 to 17.9 kJ/mol/K results in a very clear thermal profile with visible multistep unfolding processes. However, at very high values of Δ
Cp, the native baseline is not reached (i.e., a temperature range where the native state is population is effectively 100 % [
32]), which hampers the analysis of the midpoints.
In our initial effort to analyze the thermal denaturation of IF2, we used the first derivative of the experimental curve. In such an approach, issues associated with the resolvability of thermal denaturation transitions may be encountered. Based on the same data spacing used in our experiments, we compared outcomes from the first derivative with the expected values for the melting temperatures (see
Table S1,
Supplementary Materials). We found that the temperature difference between the melting temperatures of the fragments must be at least ±20 °C to detect the second melting temperature. The transition temperatures of the fragments did not change in our simulations; hence, the unfolding transitions differed only due to changes in the enthalpy change or heat capacity, and we did not observe the second melting temperature on the hot side of the thermal denaturation curve. Only on the cold side and under a special set of conditions were two cold denaturation steps visible, as highlighted by the presence of two minima of the derived curve.
In summary, simulations of independent thermal denaturation transitions were useful for achieving resolution based on our experimental method. Our simulations indicate the highly limited resolution of the denaturation curves and thermodynamic parameters for the full-length protein because of the significant overlap in the denaturation curves for both fragments. Hence, instead of studying the full-length protein, we focused on the individual fragments: the G and C fragments.
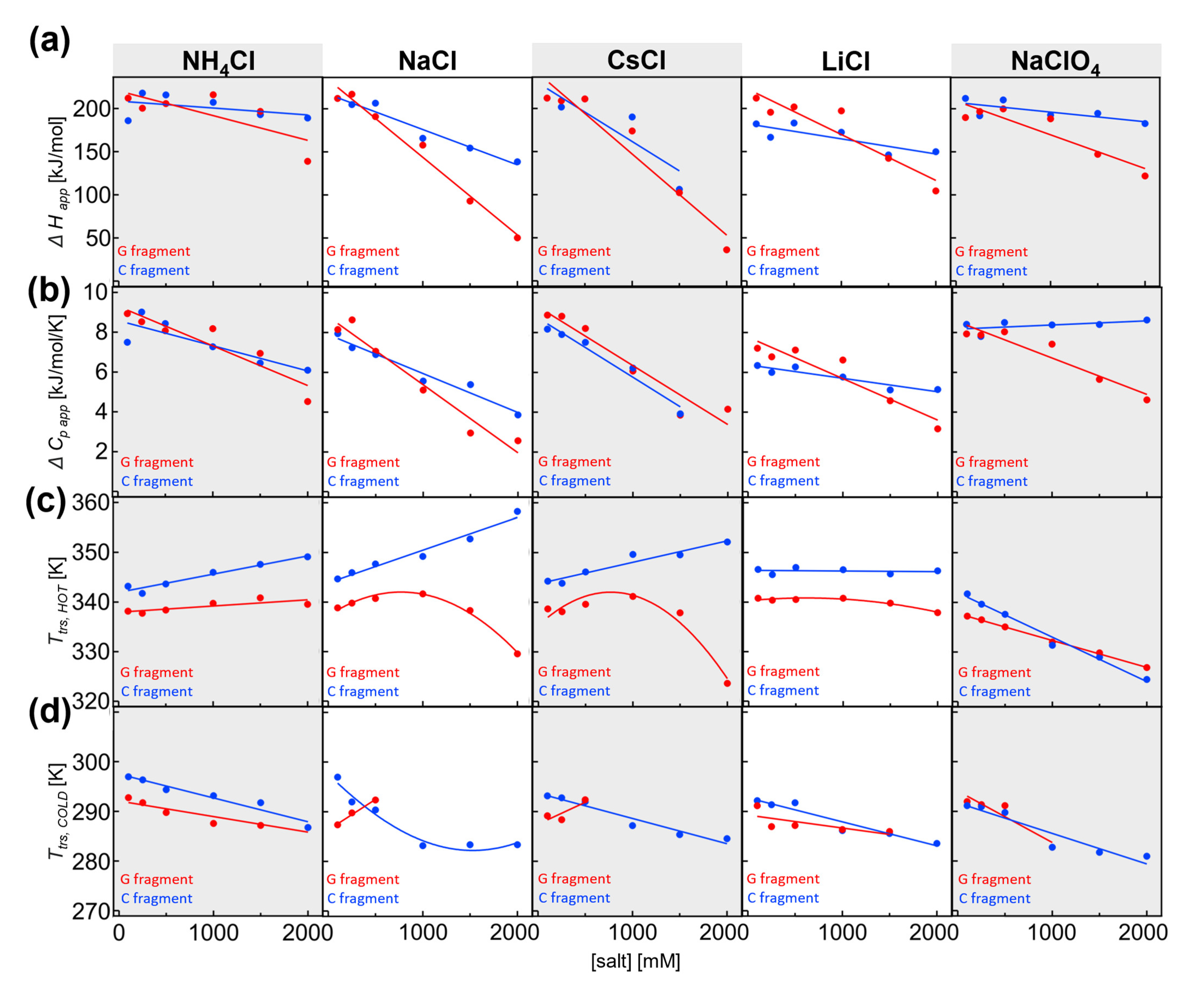
2.3. Salt-Dependent Thermal Denaturation of Individual Fragments Reveals Differences in Salt-Screenable Electrostatic Interactions
As indicated by our simulations, it is important to dissect multidomain proteins and separately analyze their fragments. To this end, we prepared and isolated individual fragments of IF2 (see [
28]). Thermal denaturation using circular dichroism in the far-UV region showed that both fragments undergo cold and hot denaturation (
Figure 3a). Using nonlinear regression analysis of the two-state model, we were able to obtain the thermodynamic values Δ
H, Δ
cp, and
Ttrs, hot/cold for the individual transitions. Due to cold and hot denaturation, all parameters could be obtained with reasonable accuracy. We found that Δ
Happ and Δ
cp, app of the individual fragments are similar regarding their full-length IF2. We wondered whether the stability of individual fragments would also display the same salt and ionic strength concentration dependence. To this end, the thermal denaturation experiments were conducted for both fragments and analyzed. The thermodynamic data are shown in
Figure 4a–d ac (G fragment in red and C fragment in blue) and are in agreement with the values found for other proteins [
11].
In the presence of NH4Cl, the unfolding enthalpy for both G and C fragments decreased. However, the salt dependence of the G fragment, was slightly higher. The same was true for Δcp, app, which decreases significantly at high NH4Cl concentrations, from 8.9 to 4.5 kJ/mol/K at 2 M. Again, the Δcp, app was slightly more salt-sensitive for the G fragment. In 0.1 M NH4Cl, the transition temperature was 65.0 °C for the G fragment and 70.0 °C for the C fragment. As part of the full-length IF2, the Ttrs, hot fragments were separated by less than 5 °C; hence, we could not resolve the individual transitions. Ttrs, hot for both fragments were dependent on NH4Cl, and we observed that Ttrs, hot was more salt-dependent for the C fragment than the G fragment.
A much larger difference and high salt sensitivity were found for NaCl; again, the unfolding enthalpy and heat capacity change decreased at high salt concentrations of NaCl. This change was even more pronounced for the G fragment when Δ
Happ reduced from 212 to 50 kJ/mol at 2 M NaCl. At the same time, the C fragment Δ
Happ decreased from 212 kJ/mol at 0.1 M NaCl to 138.5 kJ/mol at 2 M NaCl. The salt-induced change in heat capacity is clearly visible for both fragments, with Δ
cp, app reduced to 4 kJ/mol/K for the C fragment and 2 kJ/mol for the G fragment. Again, the slope is higher for the G fragment, indicating higher salt sensitivity. The transition temperature in 0.1 M NaCl is 65.7 °C for the G fragment and 71.5 °C for the C fragment. The difference between
Ttrs, hot for both fragments increases dramatically due to a significant drop in
Ttrs, hot of the G fragment at a high salt concentration, highlighting the high salt sensitivity of the G fragment. At 2 M NaCl,
Ttrs, hot is 56.4 °C for the G fragment and 85.1 °C for the C fragment, which are substantially different and could, therefore, be potentially observable in full-length IF2. Our mathematical simulations (see
Section 2.2) were performed with one variable parameter (e.g.,
Ttrs, hot), while the other thermodynamic parameters remained fixed (e.g., Δ
H, Δ
cp). Our analysis of experimental data clearly shows that all parameters are salt-dependent. Hence, they may compromise the ability to resolve individual transitions even if the difference in
Ttrs, hot is significant enough for a certain combination of the thermodynamic parameters. Using experimentally obtained thermodynamic parameters for individual domains at 2 M NaCl, we calculated the profiles of the individual denaturation curves of G and C fragments. Again, we used temperature steps and extracted the noise from experiments. After the simple addition of the calculated denaturation curves of fragments, we obtained a hypothetical denaturation curve for full IF2. Then, the calculated full IF2 denaturation curve was analyzed and subjected to visual assessment. We found that even a 28 °C difference in transition temperatures for the G and C fragments is insufficient when the denaturation enthalpies and heat capacities are very low, as was observed with 2 M NaCl. In summary, in the presence of NaCl and CsCl, the G fragment displays nonlinear salt dependence of transition temperatures, while the C fragment displays linear dependences. Individual denaturation processes for G/C domains could not be observed in the full-length protein because the small magnitude of enthalpy and heat capacity changes further decreased the resolution.
We found that in the presence of CsCl, the differences between G and C fragments were often very subtle and with similar values for ΔHapp and Δcp, app. Decreases in ΔHapp were observed at high salt concentration, with only a slight difference in the salt dependence slope for G and C fragments. As was found for other salts, Δcp, app decreases as the salt concentration is increased. Here, Δcp, app for G and C fragments demonstrated nearly equal salt sensitivity and slope compared with variations in salt. In 0.1 M CsCl, the transition temperature was 65.5 °C for the G fragment and 71.1 °C for the C fragment. Similarly, as for NaCl dependence, the difference between Ttrs, hot for both fragments increased dramatically due to the significant nonlinear drop in Ttrs, hot of the G fragment at high salt concentration, hence highlighting the high salt sensitivity of the G fragment.
At 2 M CsCl, Ttrs, hot was 50.5 °C for the G fragment and 79.0 °C for the C fragment, which represents a substantial difference that could be potentially observable in full-length IF2. Again, as in the case of 2 M NaCl, we simulated the individual unfolding transitions for the G and C fragments. Here, similar to our previous observations, the resolution was compromised due to a large decrease in enthalpy and heat capacity. Hence, we were not able to resolve different denaturation curves for the fragments.
In summary, the G fragment displayed a higher and nonlinear salt dependence than the C fragment. In 0.1 M NaClO4, the transition temperature was 58.1 °C for the G fragment and 68.6 °C for the C fragment. As a part of the full-length IF2, the Ttrs, hot of fragments differed by less than 10.5 °C; hence, we could not resolve the individual transitions. Ttrs, hot for both fragments decreased at high perchlorate concentrations. Here, the G fragment showed less salt sensitivity than the C fragment. The difference between Ttrs, hot decreased, and G and C fragments had similar melting temperatures at 1 M NaClO4, causing them to be indistinguishable in thermal denaturation experiments of the full-length protein. At higher concentrations, the melting temperatures differed only slightly for both fragments.
2.4. Salt-Dependent Thermodynamic Parameters for Denaturation of the G and C Fragments
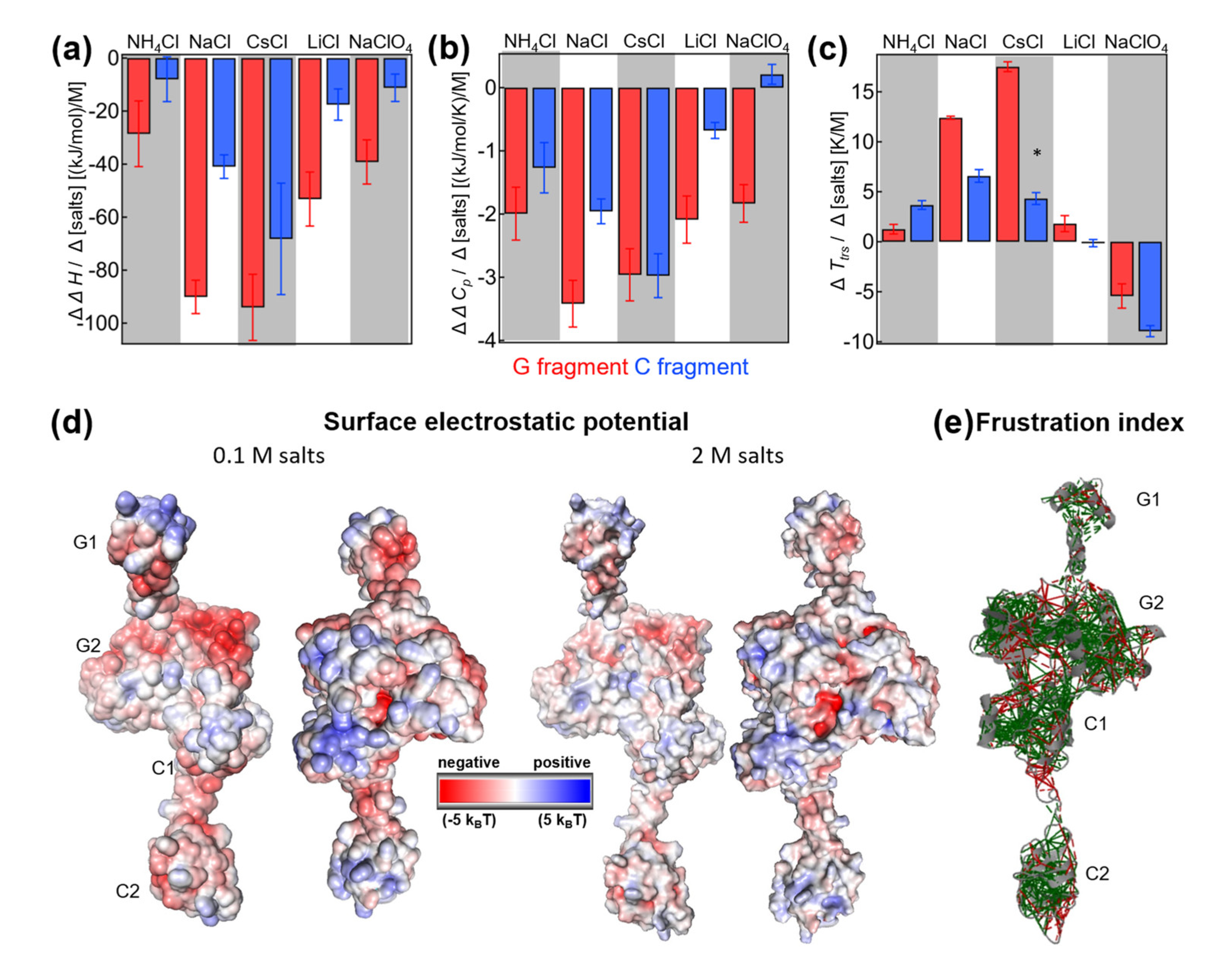
From the dependence of thermodynamic parameters on salt concentrations, we quantified how strongly these parameters depend on salt. In particular, we defined the slope of the salt dependencies (e.g., ΔΔ
H/Δ[salt]) as a suitable measure to determine the extent to which salt influences the denaturation process. By comparing different salt types, one can potentially learn about the molecular mechanism behind the salt-specific effects on cold denaturation (
Figure 5a–c).
Salt-specific enthalpy change: Overall, the G domain showed much higher sensitivity to salts than the C fragment. In absolute values, the G fragment had a higher slope (red versus blue in
Figure 5a). Under all conditions, the slopes were negative and dependent on the salt type. The largest negative slope was observed for CsCl, while NH
4Cl and NaClO
4 had the lowest slope values. The profile of salt-specific slopes was similar for G and C fragments. In summary, the analysis showed that the change in denaturation enthalpy was dependent on the type of cation and anion, and denaturation enthalpy was more highly dependent on salts for the G fragment than the C fragment.
Salt-specific heat capacity change: Again, the concentration dependence of the heat capacity was found to be higher for the G domain than the C fragment. The only exception is CsCl, where both fragments had similar slopes. The highest negative slope was observed for NaCl and the G fragment, and the lowest negative slope for LiCl and the C fragment in the case of chloride ions. The slope became slightly positive in the presence of NaClO4 for the heat capacity of the C fragment, while the G domain heat capacity had a significantly negative slope. As previously observed, the slopes were dependent on the type of salt, and the profiles observed for the G and C fragments were similar. In addition, the C fragment heat capacity showed high anion dependence when comparing the slopes of sodium chloride with those of sodium perchloride anion. The change in the slopes was much larger compared with that observed for the G fragment.
Salt-specific transition temperature changes Ttrs, hot: Transition temperatures have only been discussed on the hot side, where we have enough data points at different salt concentrations. For the cold transition, we were not able to reliably estimate the slope due to technical limitations. The G fragment transition temperatures showed high positive slopes for CsCl, while the C fragment transition temperatures showed the highest slope in NaCl. The profiles of the slopes for different salts were similar, even though differences in magnitude were present. In the case of perchlorate, both slopes were negative; however, the C fragment transition temperatures had a larger negative slope than those of the G fragment.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




