
Rhythm of the First Language: Dynamics of Extracellular Vesicle-Based Embryo–Maternal Communication in the Pre-Implantation Microenvironment


Abstract
:1. Introduction
2. Results
2.1. Trophoblast-Derived EVs Altered Endometrial Transcriptome Significantly in Each Tested Time Point
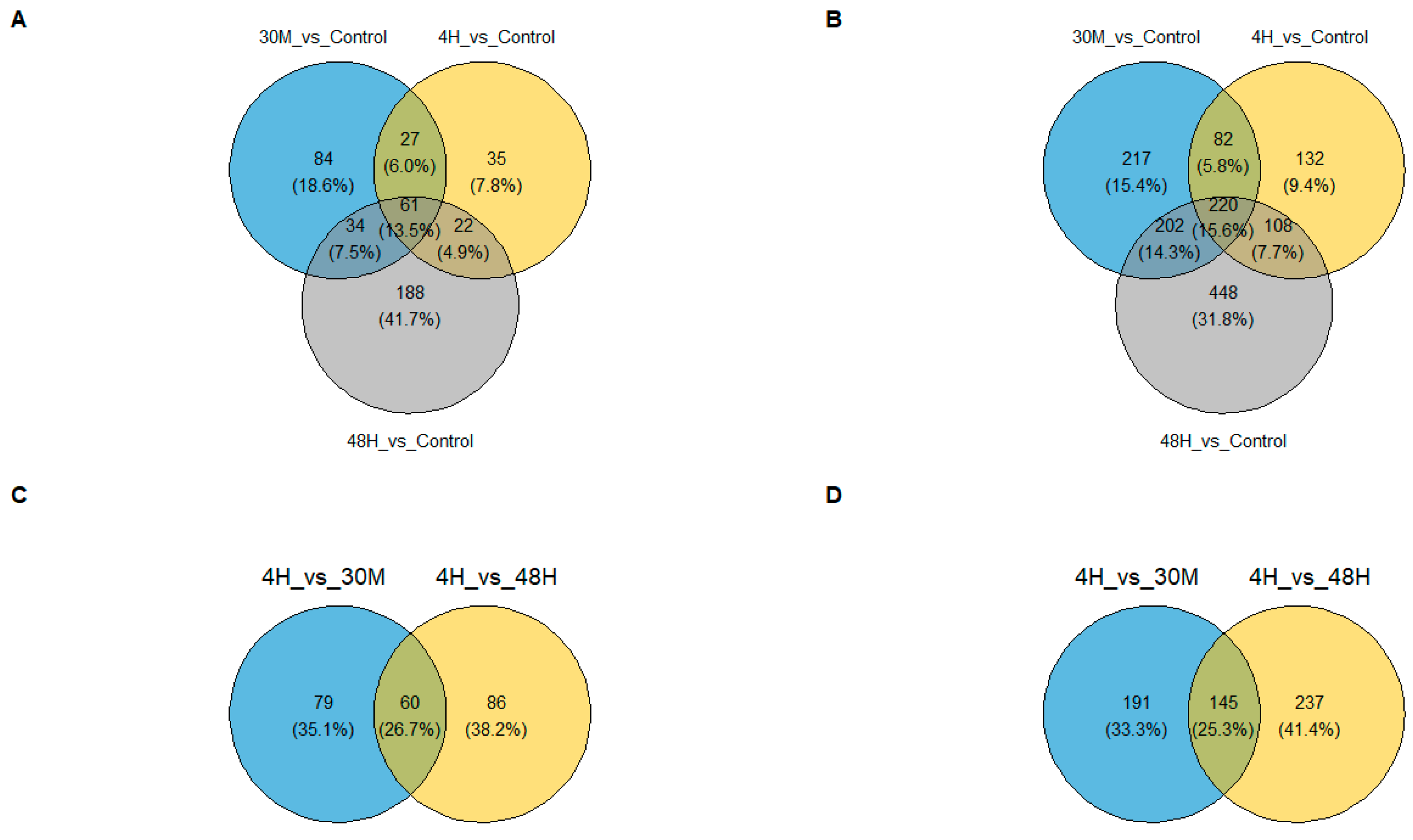
2.2. Different Sets of Genes Showed Altered Expression at Each Observed Time Point
2.3. Pathways Crucial in Embryo Implantation Were Enriched in a Specific Pattern along the Three Observed Time Points
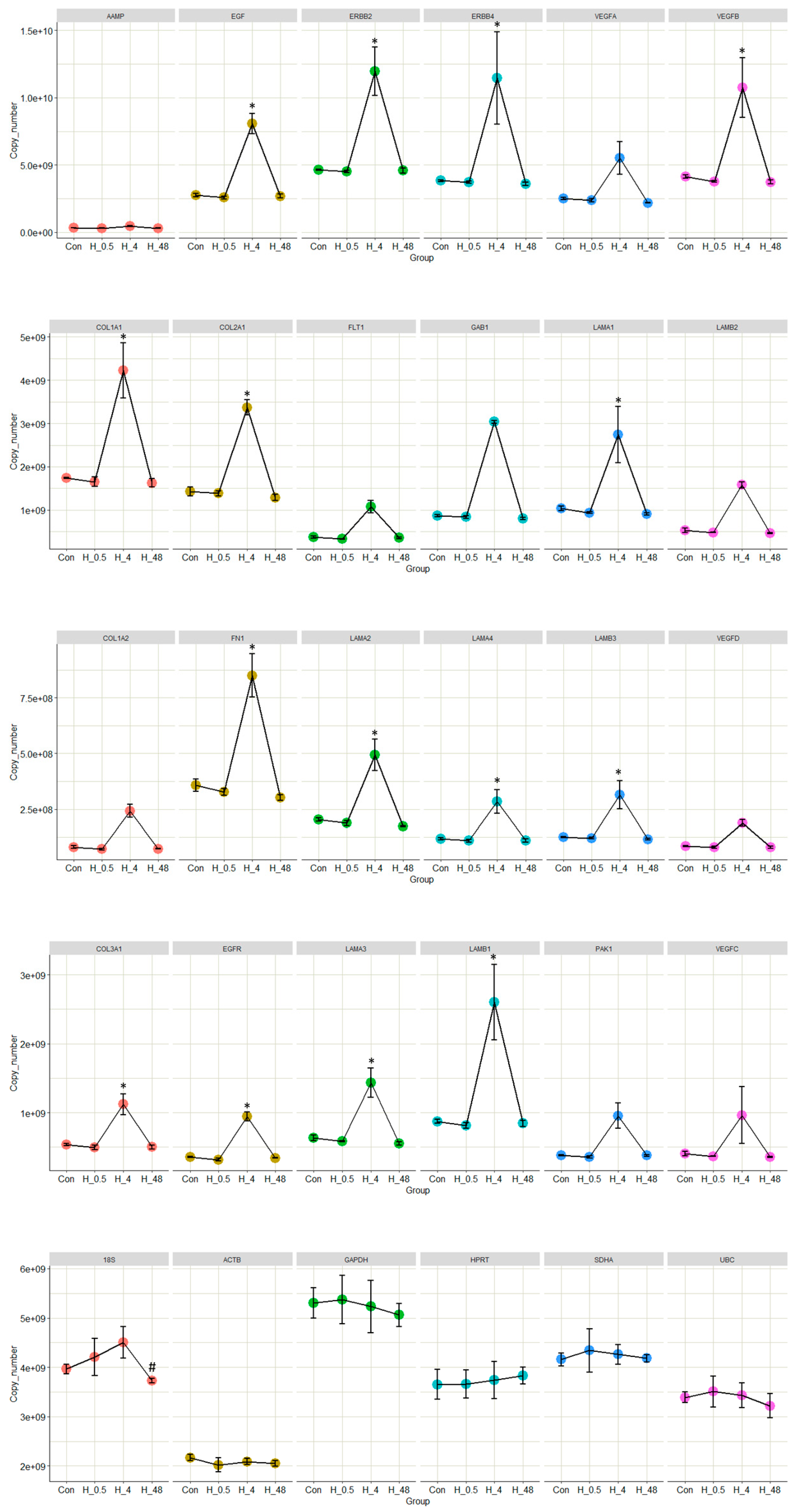
2.4. The Expressions of Key Genes in the Selected Pathways Were Significantly Altered Due to the Influence of EVs
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Trophoblast Spheroid Preparation
4.2. Depleting Extracellular Vesicles from FBS Using Ultrafiltration
4.3. Size Exclusion Chromatography for EV Enrichment
4.4. RNA Extraction and Quality Control
4.5. Library Preparation and RNAseq
4.6. Differential Gene Expression Analysis and Network Analysis
4.7. Quantitative Real-Time PCR with Absolute Quantitation
4.8. Experimental Plan
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jarvis, G.E. Early embryo mortality in natural human reproduction: What the data say. F1000Research 2016, 5, 2765. [Google Scholar] [CrossRef] [PubMed]
- Apter, S.; Balaban, B.; Campbell, A.; Catt, J.; Coticchio, G.; de los Santos, M.J.; Debrock, S.; Ebner, T. The Vienna consensus: Report of an expert meeting on the development of ART laboratory performance indicators. Reprod. Biomed. Online 2017, 35, 494–510. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, A.J.; Baird, D.D.; Weinberg, C.R. Time of Implantation of the Conceptus and Loss of Pregnancy. N. Engl. J. Med. 1999, 340, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Ashary, N.; Tiwari, A.; Modi, D. Embryo Implantation: War in times of love. Endocrinology 2018, 159, 1188–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modi, D.; Bhartiya, P. Physiology of embryo-endometrial cross talk. Biomed. Res. J. 2015, 2, 83–104. [Google Scholar] [CrossRef]
- Bhagwat, S.R.; Chandrashekar, D.S.; Kakar, R.; Davuluri, S.; Bajpai, A.K.; Nayak, S.; Bhutada, S.; Acharya, K.; Sachdeva, G. Endometrial Receptivity: A Revisit to Functional Genomics Studies on Human Endometrium and Creation of HGEx-ERdb. PLoS ONE 2013, 8, e58419. [Google Scholar] [CrossRef]
- McEwan, M.; Lins, R.J.; Munro, S.K.; Vincent, Z.L.; Ponnampalam, A.P.; Mitchell, M.D. Cytokine regulation during the formation of the fetal-maternal interface: Focus on cell-cell adhesion and remodelling of the extra-cellular matrix. Cytokine Growth Factor Rev. 2009, 20, 241–249. [Google Scholar] [CrossRef]
- Massimiani, M.; Lacconi, V.; La Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular signaling regulating endometrium–blastocyst crosstalk. Int. J. Mol. Sci. 2020, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Hantak, A.M.; Bagchi, I.C.; Bagchi, M.K. Role of uterine stromal-epithelial crosstalk in embryo implantation. Int. J. Dev. Biol. 2014, 58, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Cha, J.; Sun, X.; Dey, S.K. Mechanisms of implantation: Strategies for successful pregnancy. Nat. Med. 2012, 18, 1754–1767. [Google Scholar] [CrossRef]
- Raab, G.; Kover, K.; Paria, B.C.; Dey, S.K.; Ezzell, R.M.; Klagsbrun, M. Mouse preimplantation blastocysts adhere to cells expressing the transmembrane form of heparin-binding EGF-like growth factor. Development 1996, 122, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Stavreus-Evers, A.; Aghajanova, L.; Brismar, H.; Eriksson, H.; Landgren, B.M.; Hovatta, O. Co-existence of heparin-binding epidermal growth factor-like growth factor and pinopodes in human endometrium at the time of implantation. Mol. Hum. Reprod. 2002, 8, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Paria, B.C.; Elenius, K.; Klagsbrun, M.; Dey, S.K. Heparin-binding EGF-like growth factor interacts with mouse blastocysts independently of ErbB1: A possible role for heparan sulfate proteoglycans and ErbB4 in blastocyst implantation. Development 1999, 126, 1997–2005. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Février, B.; Raposo, G. Exosomes: Endosomal-derived vesicles shipping extracellular messages. Curr. Opin. Cell Biol. 2004, 16, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.H.; Rome, S.; Jalabert, A.; Forterre, A.; Singh, H.; Hincks, C.L.; Salamonsen, L.A. Endometrial exosomes/microvesicles in the uterine microenvironment: A new paradigm for embryo-endometrial cross talk at implantation. PLoS ONE 2013, 8, e58502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smijs, T.G.; Pavel, S.; O’Farrell, N.; Houlton, A.; Horrocks, B.R.; Yasui, T.; Yanagida, T.; Ito, S.; Konakade, Y.; Takeshita, D.; et al. Unveiling massive numbers of cancer-related urinary-microRNA candidates via nanowires. Sci. Adv. 2017, 1, 95–112. [Google Scholar] [CrossRef]
- Aleksejeva, E.; Zarovni, N.; Dissanayake, K.; Godakumara, K.; Vigano, P.; Fazeli, A.; Jaakma, Ü.; Salumets, A. Extracellular vesicle research in reproductive science— Paving the way for clinical achievements. Biol. Reprod. 2022, 106, 408–424. [Google Scholar] [CrossRef]
- Hart, A.R.; Khan, N.L.A.; Godakumara, K.; Dissanayake, K.; Piibor, J.; Muhandiram, S.; Eapen, S.; Heath, P.R.; Fazeli, A. The role of extracellular vesicles in endometrial receptivity and their potential in reproductive therapeutics and diagnosis. Reprod. Biol. 2022, 22, 100645. [Google Scholar] [CrossRef]
- Godakumara, K.; Dissanayake, K.; Hasan, M.M.; Kodithuwakku, S.P.; Fazeli, A. Role of extracellular vesicles in intercellular communication during reproduction. Reprod. Domest. Anim. 2022, 57, 14–21. [Google Scholar] [CrossRef]
- Hasan, M.M.; Reshi, Q.U.A.; Lättekivi, F.; Viil, J.; Godakumara, K.; Dissanayake, K.; Andronowska, A.; Jaakma, Ü.; Fazeli, A. Bovine Follicular Fluid Derived Extracellular Vesicles Modulate the Viability, Capacitation and Acrosome Reaction of Bull Spermatozoa. Biology 2021, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Es-Haghi, M.; Godakumara, K.; Häling, A.; Lättekivi, F.; Lavrits, A.; Viil, J.; Andronowska, A.; Nafee, T.; James, V.; Jaakma, Ü.; et al. Specific trophoblast transcripts transferred by extracellular vesicles affect gene expression in endometrial epithelial cells and may have a role in embryo-maternal crosstalk. Cell Commun. Signal. 2019, 17, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godakumara, K.; Ord, J.; Lättekivi, F.; Dissanayake, K.; Viil, J.; Boggavarapu, N.R.; Faridani, O.R.; Jääger, K.; Velthut-Meikas, A.; Jaakmaet, Ü.; et al. Trophoblast derived extracellular vesicles specifically alter the transcriptome of endometrial cells and may constitute a critical component of embryo-maternal communication. Reprod. Biol. Endocrinol. 2021, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Reshi, Q.U.A.; Viil, J.; Ord, J.; Lättekivi, F.; Godakumara, K.; Hasan, M.M.; Nõmm, M.; Jääger, K.; Velthut-Meikas, A.; Jaakmaet, Ü.; et al. Spermatozoa induce transcriptomic alterations in bovine oviductal epithelial cells prior to initial contact. J. Cell Commun. Signal 2020, 14, 439–451. [Google Scholar] [CrossRef]
- Dissanayake, K.; Nõmm, M.; Lättekivi, F.; Ord, J.; Ressaissi, Y.; Godakumara, K.; Reshi, Q.U.A.; Viil, J.; Jääger, K.; Velthut-Meikas, A.; et al. Oviduct as a sensor of embryo quality: Deciphering the extracellular vesicle (EV)-mediated embryo-maternal dialogue. J. Mol. Med. 2021, 99, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, K.; Nõmm, M.; Lättekivi, F.; Ressaissi, Y.; Godakumara, K.; Lavrits, A.; Midekessa, G.; Vill, J.; Bæk, R.; Jørgensen, M.M.; et al. Individually cultured bovine embryos produce extracellular vesicles that have the potential to be used as non-invasive embryo quality markers. Theriogenology 2020, 149, 104–116. [Google Scholar] [CrossRef]
- Gangoda, L.; Boukouris, S.; Liem, M.; Kalra, H.; Mathivanan, S. Extracellular vesicles including exosomes are mediators of signal transduction: Are they protective or pathogenic? Proteomics 2015, 15, 260. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol. 2002, 2, 569–579. [Google Scholar] [CrossRef]
- Mathieu, M.; Martin-Jaular, L.; Lavieu, G.; Théry, C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat. Cell Biol. 2019, 21, 9–17. [Google Scholar] [CrossRef]
- Bonsergent, E.; Grisard, E.; Buchrieser, J.; Schwartz, O.; Théry, C.; Lavieu, G. Quantitative characterization of extracellular vesicle uptake and content delivery within mammalian cells. Nat. Commun. 2021, 12, 1864. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Zhao, W.L.; Ye, Y.Y.; Bai, X.C.; Liu, R.Q.; Chang, L.F.; Zhou, Q.; Sui, S.-F. Cellular internalization of exosomes occurs through phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl. Acad. Sci. USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonsergent, E.; Lavieu, G. Content release of extracellular vesicles in a cell-free extract. FEBS Lett. 2019, 593, 1983–1992. [Google Scholar] [CrossRef] [PubMed]
- Chevillet, J.R.; Kang, Q.; Ruf, I.K.; Briggs, H.A.; Vojtech, L.N.; Hughes, S.M.; Arroyo, J.D.; Cheng, H.H.; Meredith, E.K.; Gallichotte, E.N.; et al. Quantitative and stoichiometric analysis of the microRNA content of exosomes. Proc. Natl. Acad. Sci. USA 2014, 111, 14888–14893. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [Green Version]
- Andreu, Z.; Yáñez-Mó, M. Tetraspanins in extracellular vesicle formation and function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef] [Green Version]
- Simeone, P.; Bologna, G.; Lanuti, P.; Pierdomenico, L.; Guagnano, M.T.; Pieragostino, D.; Del Boccio, P.; Vergara, D.; Marchisio, M.; Miscia, S.; et al. Extracellular Vesicles as Signaling Mediators and Disease Biomarkers across Biological Barriers. Int. J. Mol. Sci. 2020, 21, 2514. [Google Scholar] [CrossRef] [Green Version]
- Zhdanov, V.P. Ligand-receptor-mediated attachment of lipid vesicles to a supported lipid bilayer. Eur. Biophys. J. 2020, 49, 395. [Google Scholar] [CrossRef]
- Staufer, O.; Hernandez Bücher, J.E.; Fichtler, J.; Schröter, M.; Platzman, I.; Spatz, J.P. Vesicle Induced Receptor Sequestration: Mechanisms behind Extracellular Vesicle-Based Protein Signaling. Adv. Sci. 2022, 9, 2200201. [Google Scholar] [CrossRef]
- Torres, V.I.; Barrera, D.P.; Varas-Godoy, M.; Arancibia, D.; Inestrosa, N.C. Selective Surface and Intraluminal Localization of Wnt Ligands on Small Extracellular Vesicles Released by HT-22 Hippocampal Neurons. Front. Cell Dev. Biol. 2021, 9, 2887. [Google Scholar] [CrossRef]
- Kaloglu, C.; Onarlioglu, B. Extracellular matrix remodelling in rat endometrium during early pregnancy: The role of fibronectin and laminin. Tissue Cell 2010, 42, 301–306. [Google Scholar] [CrossRef]
- Brown, J.J.G.; Papaioannou, V.E. Distribution of hyaluronan in the mouse endometrium during the periimplantation period of pregnancy. Differentiation 1992, 52, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.A.; Burghardt, R.C.; Bazer, F.W.; Spencer, T.E. Osteopontin: Roles in Implantation and Placentation1. Biol. Reprod. 2003, 69, 1458–1471. [Google Scholar] [CrossRef]
- Miner, J.H.; Li, C.; Mudd, J.L.; Go, G.; Sutherland, A.E. Compositional and structural requirements for laminin and basement membranes during mouse embryo implantation and gastrulation. Development 2004, 131, 2247–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Dossary, A.A.; Bathala, P.; Caplan, J.L.; Martin-DeLeon, P.A. Oviductosome-Sperm Membrane Interaction in Cargo Delivery: Detection of fusion and underlying molecular players using three-dimensional super-resolution structured illumination microscopy (SR-SIM). J. Biol. Chem. 2015, 290, 17710–17723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, K.V.R.; Mangale, S.S. Integrin receptors: The dynamic modulators of endometrial function. Tissue Cell 2003, 35, 260–273. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.D. Adhesion Molecules and Implantation. In Implantation and Early Development; Cambridge University Press: Cambridge, UK, 2005; pp. 49–60. [Google Scholar] [CrossRef]
- Kirn-Safran, C.B.; D’Souza, S.S.; Carson, D.D. Heparan sulfate proteoglycans and their binding proteins in embryo implantation and placentation. Semin. Cell Dev. Biol. 2008, 19, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Large, M.J.; Wetendorf, M.; Lanz, R.B.; Hartig, S.M.; Creighton, C.J.; Mancini, M.A.; Kovanci, E.; Lee, K.-F.; Threadgill, D.W.; Lydon, J.P.; et al. The Epidermal Growth Factor Receptor Critically Regulates Endometrial Function during Early Pregnancy. PLoS Genet. 2014, 10, 1004451. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, R.; Alsat, E.; Evian-Brion, D. Alteration of epidermal growth factor receptor in placental membranes of smokers: Relationship with intrauterine growth retardation. Am. J. Obstet. Gynecol. 1994, 170, 1238–1243. [Google Scholar] [CrossRef]
- Fujita, Y.; Kurachi, H.; Morishige, K.; Amemiya, K.; Terakawa, N.; Miyake, A.; Tanizawa, O. Decrease in epidermal growth factor receptor and its messenger ribonucleic acid levels in intrauterine growth-retarded and diabetes mellitus-complicated pregnancies. J. Clin. Endocrinol. Metab. 1991, 72, 1340–1345. [Google Scholar] [CrossRef]
- Dissanayake, V.H.W.; Tower, C.; Broderick, A.; Stocker, L.J.; Seneviratne, H.R.; Jayasekara, R.W.; Kalsheker, N.; Pipkin, F.B.; Morgan, L. Polymorphism in the epidermal growth factor gene is associated with birthweight in Sinhalese and white Western Europeans. Mol. Hum. Reprod. 2007, 13, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Fondacci, C.; Alsat, E.; Gabriel, R.; Blot, P.; Nessmann, C.; Evain-Brion, D. Alterations of human placental epidermal growth factor receptor in intrauterine growth retardation. J. Clin. Investig. 1994, 93, 1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Li, M.; Cox, S.; Davis, M.K.; Tawfik, O.; Paria, B.C.; Das, S.K. HB-EGF directs stromal cell polyploidy and decidualization via cyclin D3 during implantation. Dev. Biol. 2004, 265, 181–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshihara, M.; Mizutani, S.; Kato, Y.; Matsumoto, K.; Mizutani, E.; Mizutani, H.; Fujimoto, H.; Osuka, S.; Kajiyama, H. Recent Insights into Human Endometrial Peptidases in Blastocyst Implantation via Shedding of Microvesicles. Int. J. Mol. Sci. 2021, 22, 13479. [Google Scholar] [CrossRef]
- Shi, J.W.; Lai, Z.Z.; Yang, H.L.; Yang, S.L.; Wang, C.J.; Ao, D.; Ruan, L.Y.; Shen, H.H.; Zhou, W.J.; Mei, J. Collagen at the maternal-fetal interface in human pregnancy. Int. J. Biol. Sci. 2020, 16, 2220. [Google Scholar] [CrossRef]
- Li, T.; Greenblatt, E.M.; Shin, M.E.J.; Brown, T.J.; Chan, C. Endometrial laminin subunit beta-3 expression associates with reproductive outcome in patients with repeated implantation failure. J. Assist. Reprod. Genet. 2021, 38, 1835–1842. [Google Scholar] [CrossRef]
- Guo, X.; Yi, H.; Li, T.C.; Wang, Y.; Wang, H.; Chen, X. Role of Vascular Endothelial Growth Factor (VEGF) in Human Embryo Implantation: Clinical Implications. Biomolecules 2021, 11, 253. [Google Scholar] [CrossRef]
- Klonisch, T.; Wolf, P.; Hombach-Klonisch, S.; Vogt, S.; Kuechenhoff, A.; Tetens, F.; Fischer, B. Epidermal growth factor-like ligands and erbB genes in the peri-implantation rabbit uterus and blastocyst. Biol. Reprod. 2001, 64, 1835–1844. [Google Scholar] [CrossRef] [Green Version]
- Gazor, R.; Eskandari, M.; Sharafshah, A.; Bahadori, M.H.; Golmohammadi, M.G.; Keshavarz, P. Assessment of EGFR Gene Expression Following Vitrification of 2-cell and Blastocyst Mouse Embryos. Avicenna J. Med. Biotechnol 2018, 10, 120. [Google Scholar]
- Wang, W.; Taylor, R.N.; Bagchi, I.C.; Bagchi, M.K. Regulation of human endometrial stromal proliferation and differentiation by C/EBPβ involves cyclin E-cdk2 and STAT3. Mol. Endocrinol. 2012, 26, 2016–2030. [Google Scholar] [CrossRef] [Green Version]
- Kornilov, R.; Puhka, M.; Mannerström, B.; Hiidenmaa, H.; Peltoniemi, H.; Siljander, P.; Seppänen-Kaijansinkko, R.; Kaur, S. Efficient ultrafiltration-based protocol to deplete extracellular vesicles from fetal bovine serum. J. Extracell. Vesicles 2018, 7, 1422674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. Feature Counts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kolde, R. Pheatmap: Pretty heatmaps. R Package Version 2012, 1, 726. [Google Scholar]
- Yu, G.; He, Q.Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]
- Wang, J.H.; Jiang, D.; Rao, H.Y.; Zhao, J.M.; Wang, Y.; Wei, L. Absolute quantification of serum microRNA-122 and its correlation with liver inflammation grade and serum alanine aminotransferase in chronic hepatitis C patients. Int. J. Infect. Dis. 2015, 30, e52–e56. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Total Alignments | Successfully Assigned Alignments | Percentage |
|---|---|---|---|
| 30A | 737,886 | 197,435 | 26.75 |
| 30B | 724,495 | 248,661 | 34.32 |
| 30C | 832,026 | 221,035 | 26.56 |
| 4A | 803,439 | 214,825 | 26.73 |
| 4B | 635,143 | 202,754 | 31.92 |
| 4C | 790,801 | 242,175 | 30.62 |
| 48A | 628,615 | 187,983 | 29.9 |
| 48B | 747,331 | 242,578 | 32.45 |
| 48C | 758,514 | 195,382 | 25.75 |
| ControlA | 809,428 | 224,117 | 27.68 |
| ControlB | 664,114 | 176,640 | 26.59 |
| ControlC | 641,522 | 184,343 | 28.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godakumara, K.; Heath, P.R.; Fazeli, A. Rhythm of the First Language: Dynamics of Extracellular Vesicle-Based Embryo–Maternal Communication in the Pre-Implantation Microenvironment. Int. J. Mol. Sci. 2023, 24, 6811. https://doi.org/10.3390/ijms24076811
Godakumara K, Heath PR, Fazeli A. Rhythm of the First Language: Dynamics of Extracellular Vesicle-Based Embryo–Maternal Communication in the Pre-Implantation Microenvironment. International Journal of Molecular Sciences. 2023; 24(7):6811. https://doi.org/10.3390/ijms24076811
Chicago/Turabian StyleGodakumara, Kasun, Paul R. Heath, and Alireza Fazeli. 2023. "Rhythm of the First Language: Dynamics of Extracellular Vesicle-Based Embryo–Maternal Communication in the Pre-Implantation Microenvironment" International Journal of Molecular Sciences 24, no. 7: 6811. https://doi.org/10.3390/ijms24076811
APA StyleGodakumara, K., Heath, P. R., & Fazeli, A. (2023). Rhythm of the First Language: Dynamics of Extracellular Vesicle-Based Embryo–Maternal Communication in the Pre-Implantation Microenvironment. International Journal of Molecular Sciences, 24(7), 6811. https://doi.org/10.3390/ijms24076811