
IRAPs in Combination with Highly Informative ISSRs Confer Effective Potentials for Genetic Diversity and Fidelity Assessment in Rhododendron


Abstract
:1. Introduction
2. Results
2.1. Identification of Polymorphic IRAP Markers
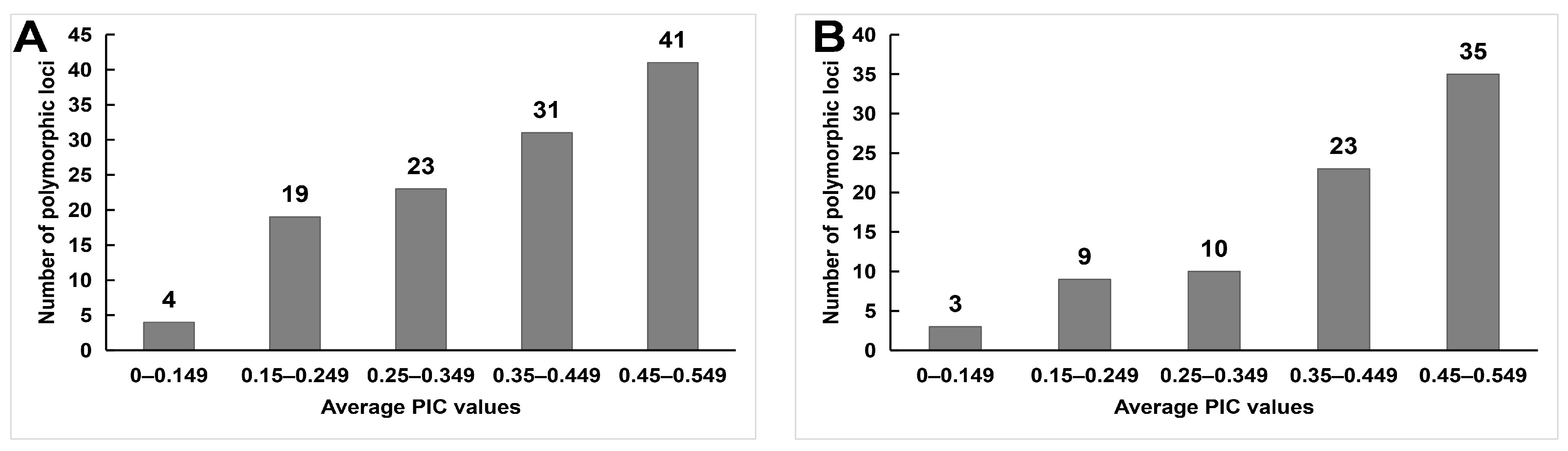
2.2. Informative Identification of IRAP and ISSR Markers
2.3. Detection of Genetic Diversity among Rhododendron Accessions
2.4. Genetic Fidelity Assessment of In Vitro Cultures of the Endangered R. bailiense
3. Discussion
3.1. IRAPs and Its Effectiveness in Genetic Divergence Detection for Rhododendron Germplasm
3.2. A Combination of IRAPs and ISSRs Shows More Effectiveness in Genetic Diversity Assessment in Rhododendrons
3.3. Somaclonal Variation as Detected by IRAPs and ISSRs
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction
4.3. LTR Retrotransposon Cloning and Sequencing
4.4. Design of IRAP Primer
4.5. Polymorphism Detection of Wild Rhododendron Accessions
4.6. Genetic Fidelity Assessment for In Vitro Cultures
4.7. Data Analysis
4.8. Cluster Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ma, H.; Liu, Y.; Liu, D.; Sun, W.; Liu, X.; Wan, Y.; Zhang, X.; Zhang, R.; Yun, Q.; Wang, J.; et al. Chromosome-level genome assembly and population genetic analysis of a critically endangered rhododendron provide insights into its conservation. Plant J. 2021, 107, 1533–1545. [Google Scholar] [CrossRef]
- Tian, X.L.; Chang, Y.H.; Neilsen, J.; Wang, S.H.; Ma, Y.P. A new species of Rhododendron (Ericaceae) from northeastern Yunnan, China. Phytotaxa 2019, 395, 66–70. [Google Scholar] [CrossRef]
- Yang, B.; Huang, M.; Wang, L.; Wu, X.; Xiaoyong, D. Fruit morphology of Ericaceaeand seed characteristics of 21 species of Rhododendron in Guizhou. Guizhou For. Sci. Technol. 2020, 48, 8–14. [Google Scholar] [CrossRef]
- Chen, X.; Huang, J.; Xie, H.; Chen, X. A New Species and a New Variety of Rhododendron (Ericaceae) in Guizhou. Seed 2010, 29, 65–67+72. [Google Scholar] [CrossRef]
- Ma, Y.; Chamberlain, D.; Sun, W.; Zhang, C. A new species of Rhododendron (Ericaceae) from Baili Rhododendron nature reserve, NW Guizhou, China. Phytotaxa 2015, 195. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Xia, G.; Zhang, M.; Huang, C. Research on the priority conservation order of rhododendron in Baili Rhododendron Provincial Nature Reserve, Guizhou. Xiang Cun Ke ji 2022, 13, 81–85. [Google Scholar] [CrossRef]
- Wei, X.Y.; Chen, J.J.; Zhang, C.Y.; Wang, Z.H. In Vitro shoot culture of Rhododendron fortunei: An important plant for bioactive phytochemicals. Ind. Crops Prod. 2018, 126, 459–465. [Google Scholar] [CrossRef]
- Nowakowska, K.; Pinkowska, A.; Siedlecka, E.; Pacholczak, A. The effect of cytokinins on shoot proliferation, biochemical changes and genetic stability of Rhododendron ‘Kazimierz Odnowiciel’ in the in vitro cultures. Plant Cell Tissue Organ Cult. 2022, 149, 675–684. [Google Scholar] [CrossRef]
- Lee, S.I.; Kim, J.H.; Park, K.C.; Kim, N.S. LTR-retrotransposons and inter-retrotransposon amplified polymorphism (IRAP) analysis in Lilium species. Genetica 2015, 143, 343–352. [Google Scholar] [CrossRef]
- Biswas, M.K.; Xu, Q.; Deng, X.X. Utility of RAPD, ISSR, IRAP and REMAP markers for the genetic analysis of Citrus spp. Sci. Hortic. 2010, 124, 254–261. [Google Scholar] [CrossRef]
- Kalendar, R.; Flavell, A.J.; Ellis, T.H.N.; Sjakste, T.; Moisy, C. Analysis of plant diversity with retrotransposon-based molecular markers. Heredity 2011, 106, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukich, M.; Schulman, A.H.; Giordani, T.; Natali, L.; Kalendar, R.; Cavallini, A. Genetic variability in sunflower (Helianthus annuus L.) and in the Helianthus genus as assessed by retrotransposon-based molecular markers. Theor. Appl. Genet. 2009, 119, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ramakrishnan, M.; Vinod, K.K.; Kalendar, R.; Yrjala, K.; Zhou, M. Development and Deployment of High-Throughput Retrotransposon-Based Markers Reveal Genetic Diversity and Population Structure of Asian Bamboo. Forests 2020, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Wen, X. Establishment and optimization of in vitro micropropagation of Rhododendron delavayi Franch. J. Southwest Univ. Nat. Sci. Ed. 2012, 34, 61–66. [Google Scholar] [CrossRef]
- He, G.-Q.; Jin, H.-Y.; Cheng, Y.-Z.; Yu, Y.-H.; Guo, D.-L. Characterization of genome-wide long terminal repeat retrotransposons provide insights into trait evolution of four grapevine species. J. Syst. Evol. 2022, 61, 414–427. [Google Scholar] [CrossRef]
- Guo, Y.; Zhai, L.N.; Long, H.; Chen, N.P.; Gao, C.X.; Ding, Z.S.; Jin, B. Genetic diversity of Bletilla striata assessed by SCoT and IRAP markers. Hereditas 2018, 155, 35. [Google Scholar] [CrossRef]
- Shingote, P.R.; Mithra, S.V.A.; Sharma, P.; Devanna, N.B.; Arora, K.; Holkar, S.K.; Khan, S.; Singh, J.; Kumar, S.; Sharma, T.R.; et al. LTR retrotransposons and highly informative ISSRs in combination are potential markers for genetic fidelity testing of tissue culture-raised plants in sugarcane. Mol. Breed. 2019, 39, 25. [Google Scholar] [CrossRef]
- Sheik, M.L.; LaBounty, K.L.; Mitchell, E.; Gillespie, E.L. Fourteen polymorphic microsatellite markers for the widespread Labrador tea (Rhododendron groenlandicum). Appl. Plant Sci. 2019, 7, e11306. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.L.; Han, F.P.; Tan, M.; Shan, X.H.; Dong, Y.Z.; Wang, X.Z.; Fedak, G.; Hao, S.; Liu, B. Activation of a rice endogenous retrotransposon Tos17 in tissue culture is accompanied by cytosine demethylation and causes heritable alteration in methylation pattern of flanking genomic regions. Theor. Appl. Genet. 2004, 109, 200–209. [Google Scholar] [CrossRef]
- Araujo, P.G.; Rossi, M.; de Jesus, E.M.; Saccaro, N.L.; Kajihara, D.; Massa, R.; de Felix, J.M.; Drummond, R.D.; Falco, M.C.; Chabregas, S.M.; et al. Transcriptionally active transposable elements in recent hybrid sugarcane. Plant J. 2005, 44, 707–717. [Google Scholar] [CrossRef]
- Novikova, T.I.; Asbaganov, S.V.; Ambros, E.V.; Zaytseva, Y.G. TDZ-induced axillary shoot proliferation of Rhododendron mucronulatum Turcz and assessment of clonal fidelity using DNA-based markers and flow cytometry. In Vitro Cell. Dev. Biol. -Plant 2020, 56, 307–317. [Google Scholar] [CrossRef]
- Mehbub, H.; Akter, A.; Akter, M.A.; Mandal, M.S.H.; Hoque, M.A.; Tuleja, M.; Mehraj, H. Tissue culture in ornamentals: Cultivation factors, propagation techniques, and its application. Plants 2022, 11, 3208. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, B.; Gill, N.; Hamelin, R.C.; Goodwin, S.B. The landscape of transposable elements in the finished genome of the fungal wheat pathogen Mycosphaerella graminicola. BMC Genom. 2014, 15, 1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaytseva, Y.G.; Skaptsov, M.V.; Kutsev, M.G.; Novikova, T.I. In vitro establishment and TDZ-induced regeneration from shoot and leaf explants of Rhododendron sichotense Pojark. Turczaninowia 2020, 23, 106–111. [Google Scholar] [CrossRef]
- Vejsadova, H.; Pretova, A. Somatic embryogenesis in Rhododendron catawbiense ‘Grandiflorum’. In Proceedings of the 1st International Symposium on Acclimatization and Establishment of Micropropagated Plants, Sani-Halkidiki, Greece, 19–22 September 2001; pp. 467–470. [Google Scholar]
- Kumar, A.; Pearce, S.R.; McLean, K.; Harrison, G.; Heslop-Harrison, J.S.; Waugh, R.; Flavell, A.J. The Ty1-Copia group of retrotransposons in plants: Genomic organisation, evolution, and use as molecular markers. Genetica 1997, 100, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Kumekawa, N.; Ohtsubo, E.; Ohtsubo, H. Identification and phylogenetic analysis of Gypsy-type retrotransposons in the plant kingdom. Genes Genet. Syst. 1999, 74, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, P.; Cai, Y.; Ma, L.; Li, S.; Li, S.; Xie, W.; Song, J.; Peng, L.; Yan, H.; et al. The draft genome assembly of Rhododendron delavayi Franch. var. delavayi. Gigascience 2017, 6, gix076. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Yeh, F.; Yang, R.; Boyle, T. POPGENE Version 1.32 Microsoft Windows-Based Freeware for Populations Genetic Analysis; University of Alberta: Edmonton, Canada, 1999. [Google Scholar]
- Adhikari, S.; Saha, S.; Bandyopadhyay, T.K.; Ghosh, P. Efficiency of ISSR marker for characterization of Cymbopogon germplasms and their suitability in molecular barcoding. Plant Syst. Evol. 2014, 301, 439–450. [Google Scholar] [CrossRef]
- Roldan-Ruiz, I.; Dendauw, J.; Van Bockstaele, E.; Depicker, A.; De Loose, M. AFLP markers reveal high polymorphic rates in ryegrasses (Lolium spp.). Mol. Breed. 2000, 6, 125–134. [Google Scholar] [CrossRef]
- Varshney, R.K.; Chabane, K.; Hendre, P.S.; Aggarwal, R.K.; Graner, A. Comparative assessment of EST-SSR, EST-SNP and AFLP markers for evaluation of genetic diversity and conservation of genetic resources using wild, cultivated and elite barleys. Plant Sci. 2007, 173, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ne | H | I | Number of Polymorphic Alleles | Range of Alleles (kb) | PIC | EMR | Rp | MI | Annealing Temperature (°C) | |
|---|---|---|---|---|---|---|---|---|---|---|
| IRAP | ||||||||||
| LTR1–7 | 1.62 | 0.37 | 0.55 | 13 | 0.2–2.3 | 0.37 | 12.07 | 6.98 | 4.41 | 56 |
| LTR1–10 | 1.54 | 0.33 | 0.51 | 15 | 0.3–2.7 | 0.33 | 15.00 | 7.20 | 5.01 | 59 |
| LTR1–11 | 1.64 | 0.37 | 0.56 | 13 | 0.4–2.4 | 0.37 | 14.00 | 7.07 | 5.25 | 59 |
| LTR1–12 | 1.47 | 0.30 | 0.47 | 16 | 0.3–2.9 | 0.30 | 16.00 | 6.38 | 4.83 | 59 |
| LTR1–15 | 1.51 | 0.32 | 0.49 | 15 | 0.2–2.4 | 0.32 | 13.24 | 6.70 | 4.20 | 55 |
| LTR1–16 | 1.51 | 0.31 | 0.48 | 14 | 0.15–2.6 | 0.31 | 13.07 | 6.18 | 4.11 | 55 |
| LTR1–17 | 1.77 | 0.42 | 0.61 | 16 | 0.2–2.5 | 0.42 | 16.00 | 11.07 | 6.78 | 56 |
| LTR3–21 | 1.76 | 0.43 | 0.61 | 17 | 0.2–2.5 | 0.43 | 17.00 | 11.00 | 7.23 | 57 |
| Mean | 1.60 | 0.36 | 0.53 | 14.88 | - | 0.36 | 14.55 | 7.82 | 5.23 | 57 |
| ISSR | ||||||||||
| UBC826 | 1.79 | 0.43 | 0.62 | 10 | 0.3–2.0 | 0.43 | 8.33 | 7.16 | 3.61 | 59 |
| UBC835 | 1.72 | 0.41 | 0.60 | 15 | 0.3–2.4 | 0.41 | 15.00 | 9.14 | 6.20 | 59 |
| UBC836 | 1.69 | 0.39 | 0.58 | 16 | 0.2–2.6 | 0.38 | 16.00 | 9.54 | 6.05 | 59 |
| UBC840 | 1.83 | 0.45 | 0.64 | 16 | 0.2–2.3 | 0.45 | 16.00 | 12.00 | 7.15 | 59 |
| UBC890 | 1.59 | 0.34 | 0.51 | 11 | 0.3–2.3 | 0.34 | 9.31 | 6.18 | 3.18 | 59 |
| UBCM06 | 1.48 | 0.30 | 0.46 | 11 | 0.2–2.2 | 0.30 | 8.64 | 4.75 | 2.55 | 59 |
| Mean | 1.69 | 0.39 | 0.57 | 13.17 | - | 0.38 | 12.21 | 8.13 | 4.79 | 59 |
| Populations | Sample Size | Ne | H | I |
|---|---|---|---|---|
| R. delavayi | 11 | 1.32 | 0.21 | 0.33 |
| R. decorum | 13 | 1.38 | 0.25 | 0.39 |
| R. agastum | 5 | 1.35 | 0.22 | 0.33 |
| Primer ID | Primer Sequence (5′–3′) | Number of Scorable Bands | Number of Polymorphic Band |
|---|---|---|---|
| IRAP | |||
| LTR1-10 | ATGGTCTTAAGCAGTCACCTC | 8 | 2 |
| LTR1-11 | GGGCTGAAACAGTCTCCAAGA | 7 | 6 |
| LTR1-12 | CTCCCAGACAGTGGTACAGAA | 8 | 6 |
| LTR1-15 | CCCAGACAGTGGTACAGAAAG | 7 | 7 |
| LTR1-16 | ACAGGCTCCCAGACAGTGGTA | 6 | 2 |
| LTR1-17 | GAGTCTGTATGGTTTGAAACA | 9 | 9 |
| ISSR | |||
| UBC826 | ACACACACACACACACC | 9 | 7 |
| UBC835 | AGAGAGAGAGAGAGAGTC | 7 | 4 |
| UBC836 | AGAGAGAGAGAGAGAGYA | 5 | 3 |
| UBC840 | GAGAGAGAGAGAGAGAYT | 5 | 2 |
| UBC890 | VHVGTGTGTGTGTGTGT | 7 | 0 |
| UBCM06 | AGCAGCAGCAGCY | 9 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, S.; Zhao, H.; Zhang, M.; Qiao, G.; Shen, X. IRAPs in Combination with Highly Informative ISSRs Confer Effective Potentials for Genetic Diversity and Fidelity Assessment in Rhododendron. Int. J. Mol. Sci. 2023, 24, 6902. https://doi.org/10.3390/ijms24086902
Wen S, Zhao H, Zhang M, Qiao G, Shen X. IRAPs in Combination with Highly Informative ISSRs Confer Effective Potentials for Genetic Diversity and Fidelity Assessment in Rhododendron. International Journal of Molecular Sciences. 2023; 24(8):6902. https://doi.org/10.3390/ijms24086902
Chicago/Turabian StyleWen, Sulin, Hong Zhao, Manying Zhang, Guang Qiao, and Xiaohui Shen. 2023. "IRAPs in Combination with Highly Informative ISSRs Confer Effective Potentials for Genetic Diversity and Fidelity Assessment in Rhododendron" International Journal of Molecular Sciences 24, no. 8: 6902. https://doi.org/10.3390/ijms24086902
APA StyleWen, S., Zhao, H., Zhang, M., Qiao, G., & Shen, X. (2023). IRAPs in Combination with Highly Informative ISSRs Confer Effective Potentials for Genetic Diversity and Fidelity Assessment in Rhododendron. International Journal of Molecular Sciences, 24(8), 6902. https://doi.org/10.3390/ijms24086902