

Optimization of Monobenzone-Induced Vitiligo Mouse Model by the Addition of Chronic Stress


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
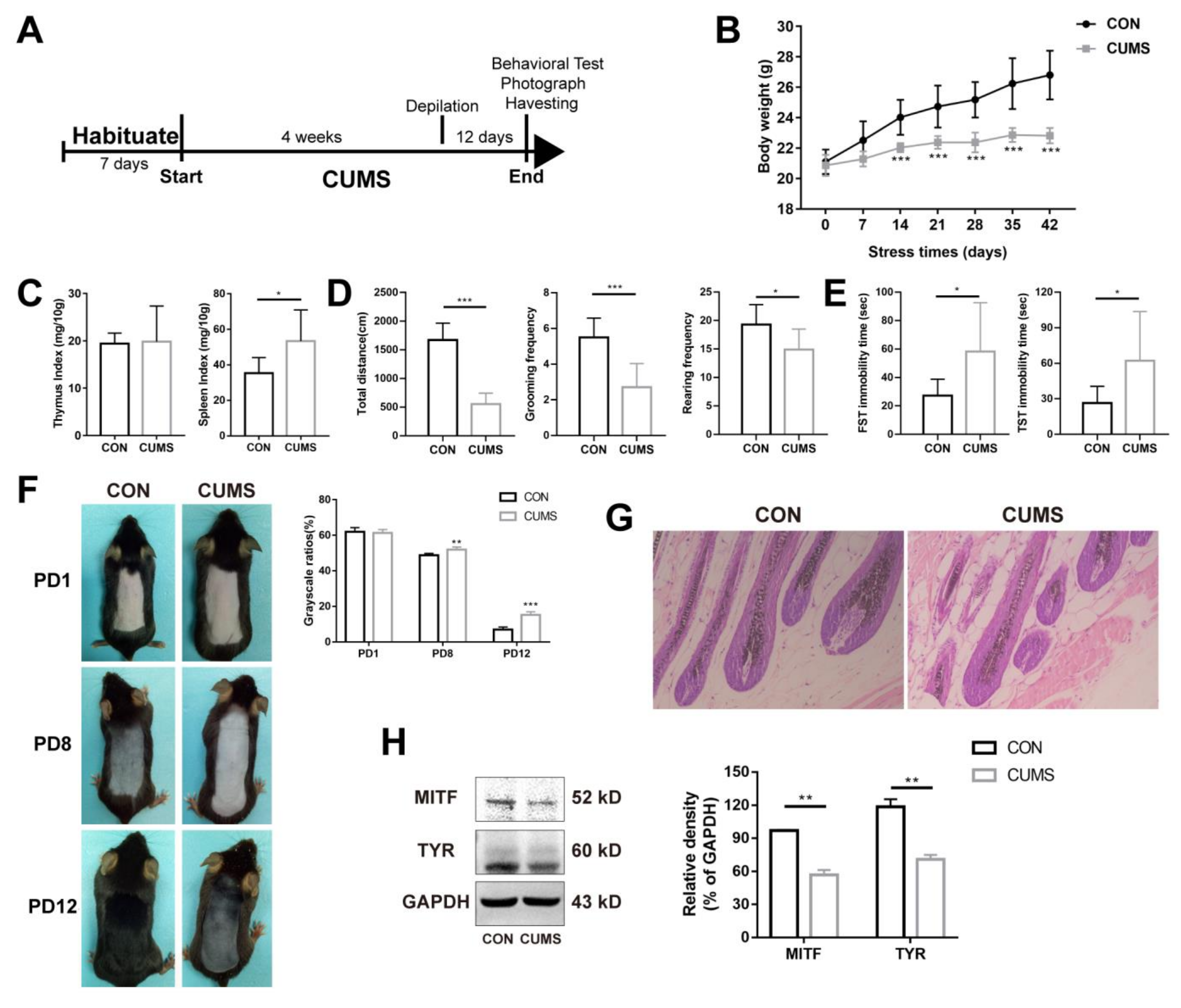
2.1. Chronic Stress Suppresses the Melanogenesis in Skin
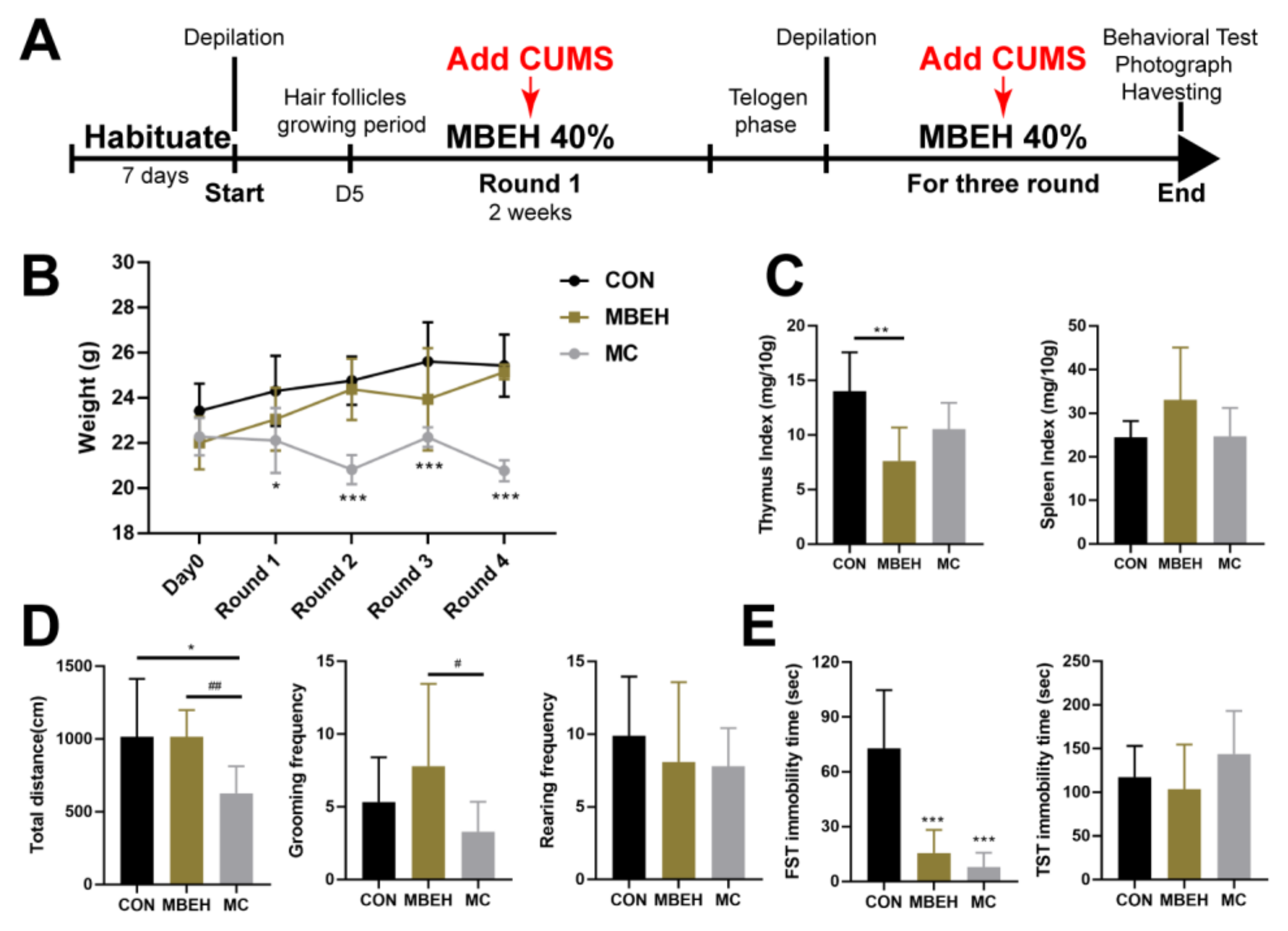
2.2. MBEH Combined with CUMS (MC) Leads to Depression-like Behavior in Mice
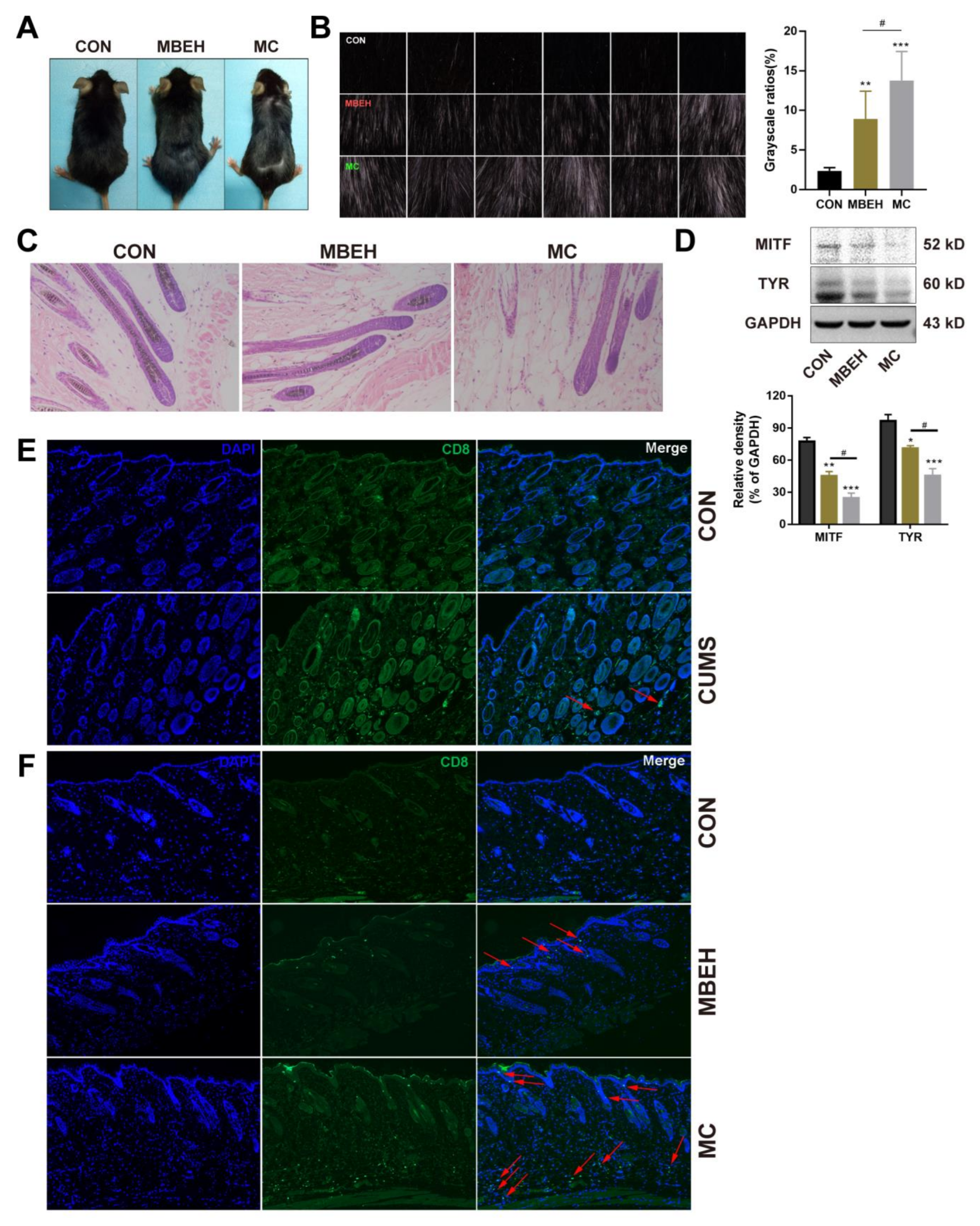
2.3. Mice in the MC Group Showed More Inhibition in Melanogenesis
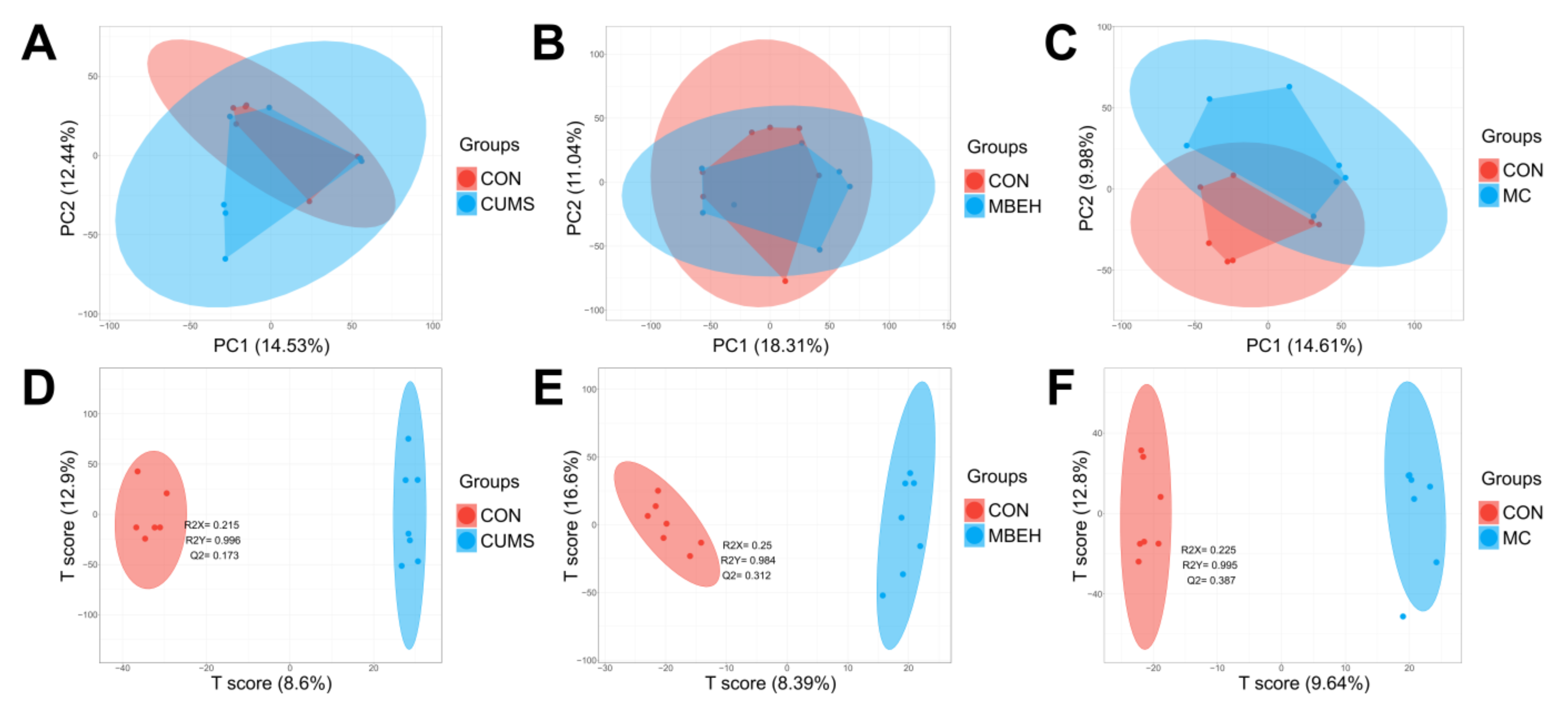
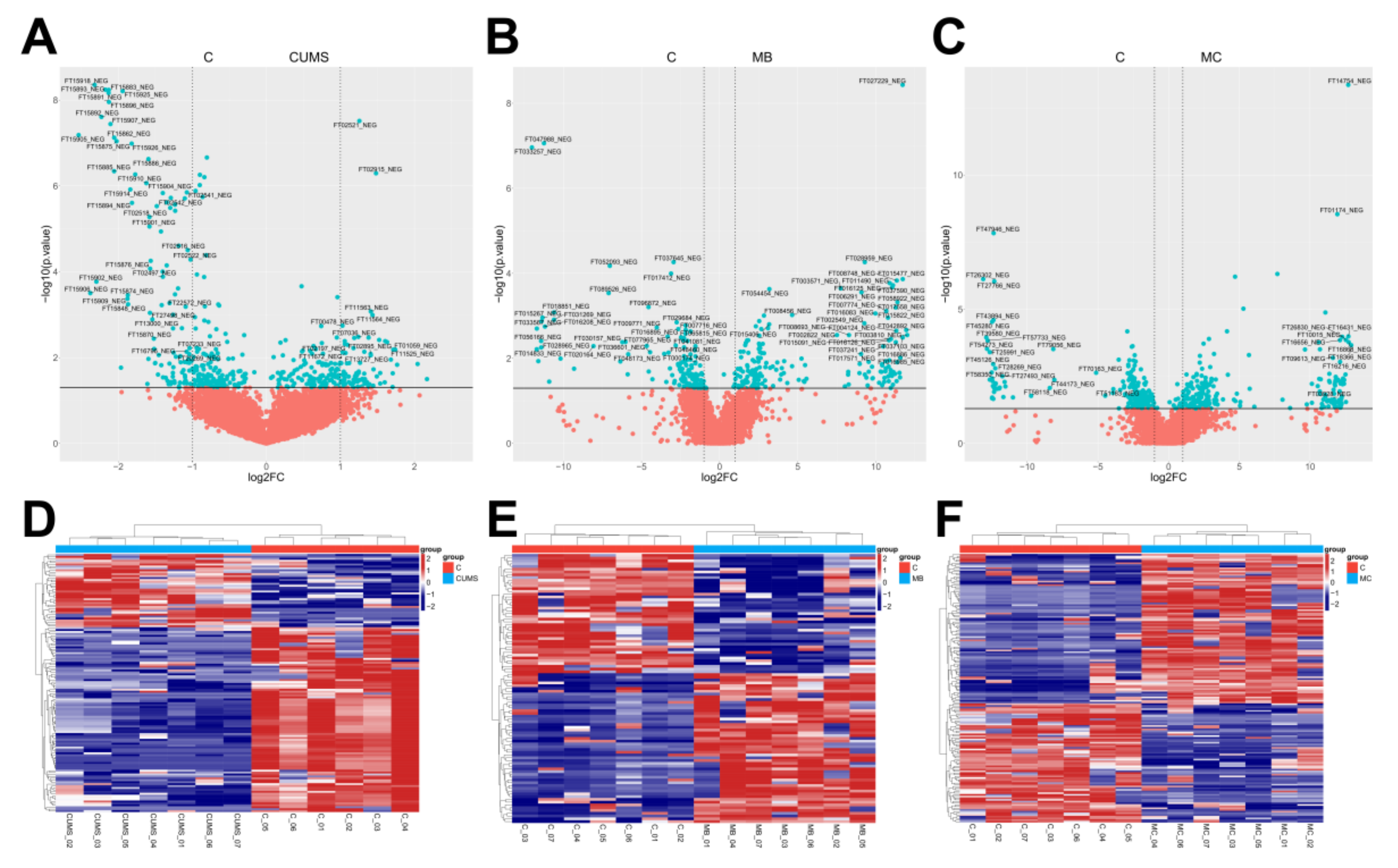
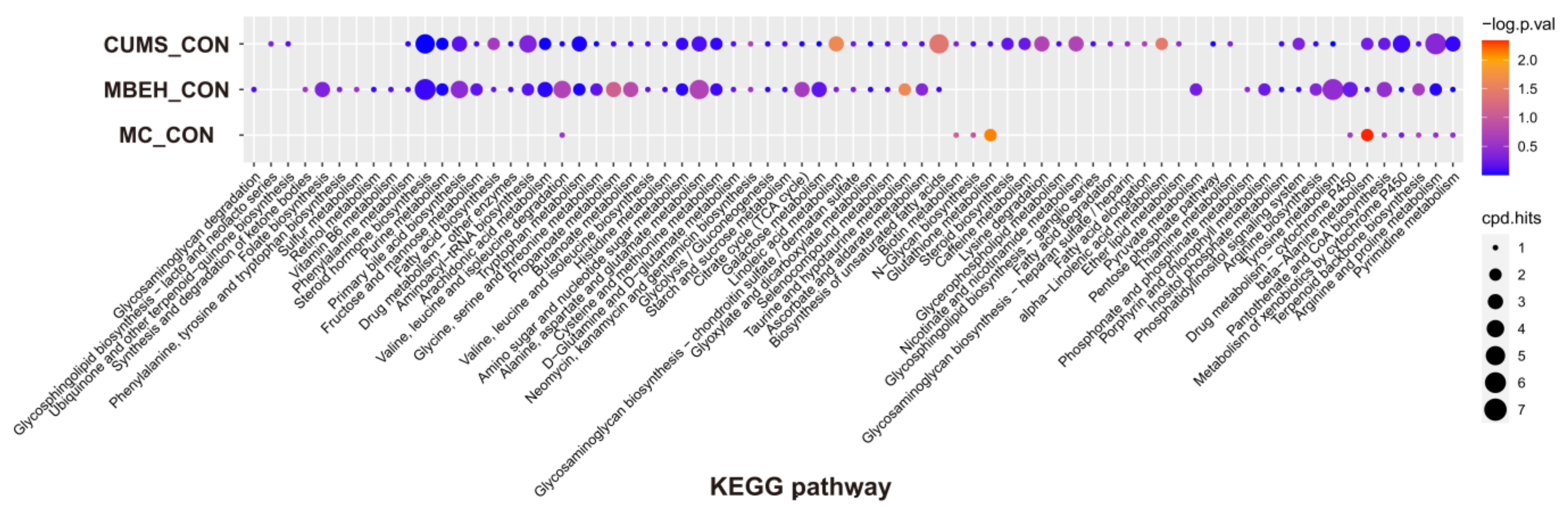
2.4. Metabolic Analysis
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. CUMS Model
4.3. MBEH Model
4.4. MBEH Combined with CUMS (MC) Model
4.5. Behavior Test
4.6. Hematoxylin-Eosin Staining
4.7. Immunofluorescence
4.8. Western Blotting (WB) Analysis
4.9. Liquid Chromatography-Mass Spectrometer (LC-MS) Analysis
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ezzedine, K.; Eleftheriadou, V.; Whitton, M.; van Geel, N. Vitiligo. Lancet 2015, 386, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Picardo, M.; Dell'Anna, M.L.; Ezzedine, K.; Hamzavi, I.; Harris, J.E.; Parsad, D.; Taieb, A. Vitiligo. Nat. Rev. Dis. Prim. 2015, 1, 15011. [Google Scholar] [CrossRef] [PubMed]
- Whitton, M.E.; Pinart, M.; Batchelor, J.; Leonardi-Bee, J.; González, U.; Jiyad, Z.; Eleftheriadou, V.; Ezzedine, K. Interventions for vitiligo. Cochrane Database Syst. Rev. 2015, 24, Cd003263. [Google Scholar] [CrossRef]
- Das, P.K.; van den Wijngaard, R.F.; Wankowicz-Kalinska, A.; Wankowicz-Kalinska, A.; Le Poole, I.C. A symbiotic concept of autoimmunity and tumour immunity: Lessons from vitiligo. Trends Immunol. 2001, 22, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nishikawa, H.; Sugiyama, D.; Ha, D.; Hamaguchi, M.; Saito, T.; Nishioka, M.; Wing, J.B.; Adeegbe, D.; Katayama, I.; et al. Detection of self-reactive cd8(+) t cells with an anergic phenotype in healthy individuals. Science 2014, 346, 1536–1540. [Google Scholar] [CrossRef]
- Taieb, A.; Picardo, M. Clinical practice. Vitiligo. N. Engl. J. Med. 2009, 360, 160–169. [Google Scholar]
- Zhou, T.; Li, D.; Deng, Y. Update on the role of noncoding rnas in vitiligo. Chin. Med. J. 2022, 135, 793–795. [Google Scholar] [CrossRef] [PubMed]
- Ezzedine, K.; Lim, H.W.; Suzuki, T.; Katayama, I.; Hamzavi, I.; Lan, C.C.; Goh, B.K.; Anbar, T.; Silva de Castro, C.; Lee, A.Y.; et al. Revised classification/nomenclature of vitiligo and related issues: The vitiligo global issues consensus conference. Pigment. Cell Melanoma Res. 2012, 25, E1–E13. [Google Scholar] [CrossRef] [Green Version]
- Taieb, A.; Alomar, A.; Böhm, M.; Dell'anna, M.L.; De Pase, A.; Eleftheriadou, V.; Ezzedine, K.; Gauthier, Y.; Gawkrodger, D.J.; Jouary, T.; et al. Guidelines for the management of vitiligo: The european dermatology forum consensus. Br. J. Dermatol. 2013, 168, 5–19. [Google Scholar] [CrossRef]
- Manolache, L.; Benea, V. Stress in patients with alopecia areata and vitiligo. J. Eur. Acad. Dermatol. Venereol. 2007, 21, 921–928. [Google Scholar] [CrossRef]
- Dai, Y.X.; Tai, Y.H.; Chang, Y.T.; Chen, T.J.; Chen, M.H. Association between major depressive disorder and subsequent autoimmune skin diseases: A nationwide population-based cohort study. J. Affect. Disord. 2020, 274, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Ferentinos, P.; Kalogeropoulou, E.; Pappa, G.; Antoniou, A.; Bozi, E.; Kyprianou, M.; Rigopoulos, D.; Katoulis, A.C. Assessing the role of stressful life events in the induction and recurrence of alopecia areata: A case-control study. J. Am. Acad. Dermatol. 2022, 87, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- O'Sullivan, J.D.B.; Peters, E.M.J.; Amer, Y.; Atuluru, P.; Chéret, J.; Rosenberg, A.M.; Picard, M.; Paus, R. The impact of perceived stress on the hair follicle: Towards solving a psychoneuroendocrine and neuroimmunological puzzle. Front. Neuroendocrinol. 2022, 66, 101008. [Google Scholar] [CrossRef] [PubMed]
- Silpa-Archa, N.; Pruksaeakanan, C.; Angkoolpakdeekul, N.; Chaiyabutr, C.; Kulthanan, K.; Ratta-Apha, W.; Wongpraparut, C. Relationship between depression and quality of life among vitiligo patients: A self-assessment questionnaire-based study. Clin. Cosmet. Investig. Dermatol. 2020, 13, 511–520. [Google Scholar] [CrossRef]
- Condamina, M.; Shourick, J.; Seneschal, J.; Sbidian, E.; Andreu, N.; Pane, I.; Ravaud, P.; Tran, V.T.; Ezzedine, K. Factors associated with perceived stress in patients with vitiligo in the compare e-cohort. J. Am. Acad. Dermatol. 2022, 86, 696–698. [Google Scholar] [CrossRef]
- Kussainova, A.; Kassym, L.; Akhmetova, A.; Glushkova, N.; Sabirov, U.; Adilgozhina, S.; Tuleutayeva, R.; Semenova, Y. Vitiligo and anxiety: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0241445. [Google Scholar] [CrossRef]
- Vallerand, I.A.; Lewinson, R.T.; Parsons, L.M.; Hardin, J.; Haber, R.M.; Lowerison, M.W.; Barnabe, C.; Patten, S.B. Vitiligo and major depressive disorder: A bidirectional population-based cohort study. J. Am. Acad. Dermatol. 2019, 80, 1371–1379. [Google Scholar] [CrossRef]
- Patel, K.R.; Singam, V.; Rastogi, S.; Lee, H.H.; Silverberg, N.B.; Silverberg, J.I. Association of vitiligo with hospitalization for mental health disorders in us adults. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Erdoğan, A.; Mutlu, H.S.; Solakoğlu, S. Autologously transplanted dermis-derived cells alleviated monobenzone-induced vitiligo in mouse. Exp. Dermatol. 2022, 31, 1355–1363. [Google Scholar] [CrossRef]
- Huo, S.X.; Wang, Q.; Liu, X.M.; Ge, C.H.; Gao, L.; Peng, X.M.; Yan, M. The effect of butin on the vitiligo mouse model induced by hydroquinone. Phytother. Res. 2017, 31, 740–746. [Google Scholar] [CrossRef]
- Harris, J.E. Chemical-induced vitiligo. Dermatol. Clin. 2017, 35, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Boorn, J.G.; Picavet, D.I.; van Swieten, P.F.; van Veen, H.A.; Konijnenberg, D.; van Veelen, P.A.; van Capel, T.; Jong, E.C.; Reits, E.A.; Drijfhout, J.W.; et al. Skin-depigmenting agent monobenzone induces potent t-cell autoimmunity toward pigmented cells by tyrosinase haptenation and melanosome autophagy. J. Investig. Dermatol. 2011, 131, 1240–1251. [Google Scholar] [CrossRef] [Green Version]
- Teulings, H.E.; Tjin, E.P.M.; Willemsen, K.J.; van der Kleij, S.; Ter Meulen, S.; Kemp, E.H.; Krebbers, G.; van Noesel, C.J.M.; Franken, C.; Drijfhout, J.W.; et al. Anti-melanoma immunity and local regression of cutaneous metastases in melanoma patients treated with monobenzone and imiquimod; a phase 2 a trial. Oncoimmunology 2018, 7, e1419113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wang, S.; Xu, A. A mouse model of vitiligo induced by monobenzone. Exp. Dermatol. 2013, 22, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Speeckaert, R.; Voet, S.; Hoste, E.; van Geel, N. S100b is a potential disease activity marker in nonsegmental vitiligo. J. Investig. Dermatol. 2017, 137, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Manini, P.; Napolitano, A.; Westerhof, W.; Riley, P.A.; d'Ischia, M. A reactive ortho-quinone generated by tyrosinase-catalyzed oxidation of the skin depigmenting agent monobenzone: Self-coupling and thiol-conjugation reactions and possible implications for melanocyte toxicity. Chem. Res. Toxicol. 2009, 22, 1398–1405. [Google Scholar] [CrossRef]
- Mosenson, J.A.; Flood, K.; Klarquist, J.; Eby, J.M.; Koshoffer, A.; Boissy, R.E.; Overbeck, A.; Tung, R.C.; Le Poole, I.C. Preferential secretion of inducible hsp70 by vitiligo melanocytes under stress. Pigment. Cell Melanoma Res. 2014, 27, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Liao, S.; Lv, J.; Zhou, J.; Kalavagunta, P.K.; Shang, J. Effects of two chronic stresses on mental state and hair follicle melanogenesis in mice. Exp. Dermatol. 2017, 26, 1083–1090. [Google Scholar] [CrossRef]
- Chen, M.; Cai, J.; Zhang, X.; Liao, Z.; Zhong, M.; Shang, J.; Yue, Y. Keratinocytes take part in the regulation of substance p in melanogenesis through the hpa axis. J. Dermatol. Sci. 2022, 106, 141–149. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, Y.; Pang, S.; Zhou, J.; Cai, J.; Shang, J. Alcohol extract from vernonia anthelmintica willd (L.) seed counteracts stress-induced murine hair follicle growth inhibition. BMC Complement. Altern. Med. 2019, 19, 372. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wu, H.; Zhou, J.; Pei, S.; Li, J.; Cai, Y.; Shang, J. Involvement of the central hypothalamic-pituitary-adrenal axis in hair growth and melanogenesis among different mouse strains. PLoS ONE 2018, 13, e0202955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, M.; Tang, Q.; Li, X.; Zhao, R.; Li, J.; Xu, H.; Gao, Y.; Mao, Y. Shen-qi-jie-yu-fang has antidepressant effects in a rodent model of postpartum depression by regulating the immune organs and subsets of t lymphocytes. Neuropsychiatr. Dis. Treat. 2015, 11, 1523–1540. [Google Scholar] [PubMed] [Green Version]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A.; Brański, P.; Burnat, G. The influence of the duration of chronic unpredictable mild stress on the behavioural responses of c57bl/6j mice. Behav. Pharmacol. 2020, 31, 574–582. [Google Scholar] [CrossRef]
- Mort, R.L.; Jackson, I.J.; Patton, E.E. The melanocyte lineage in development and disease. Development 2015, 142, 1387. [Google Scholar] [CrossRef] [Green Version]
- Steingrímsson, E.; Copeland, N.G.; Jenkins, N.A. Melanocytes and the microphthalmia transcription factor network. Annu. Rev. Genet. 2004, 38, 365–411. [Google Scholar] [CrossRef]
- Lister, J.A.; Capper, A.; Zeng, Z.; Mathers, M.E.; Richardson, J.; Paranthaman, K.; Jackson, I.J.; Patton, E.E. A conditional zebrafish mitf mutation reveals mitf levels are critical for melanoma promotion vs. Regression in vivo. J. Investig. Dermatol. 2014, 134, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Fu, Y.; Cao, Y.; Jiang, S.; Yang, Y.; Song, G.; Yun, C.; Gao, R. Isoliquiritigenin inhibits melanogenesis, melanocyte dendricity and melanosome transport by regulating erk-mediated mitf degradation. Exp. Dermatol. 2020, 29, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahin, Z.; Leheta, T.M.; Abdel Hay, R.M.; Aal, H.M.A.; Rashed, L.A. Detection of plasma and urinary monoamines and their metabolites in nonsegmental vitiligo. Acta Dermatovenerol. Croat. 2012, 20, 14–20. [Google Scholar]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin's neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 5–115. [Google Scholar]
- Korabel, H.; Dudek, D.; Jaworek, A.; Wojas-Pelc, A. psychodermatology: Psychological and psychiatrical aspects of dermatology. Przegl. Lek. 2008, 65, 244–248. [Google Scholar]
- Dainichi, T.; Kabashima, K. Alopecia areata: What's new in epidemiology, pathogenesis, diagnosis, and therapeutic options? J. Dermatol. Sci. 2017, 86, 3–12. [Google Scholar] [CrossRef]
- Evers, A.W.; Verhoeven, E.W.; Kraaimaat, F.W.; de Jong, E.M.; de Brouwer, S.J.; Schalkwijk, J.; Sweep, F.C.; van de Kerkhof, P.C. How stress gets under the skin: Cortisol and stress reactivity in psoriasis. Br. J. Dermatol. 2010, 163, 986–991. [Google Scholar] [CrossRef]
- Hunter, H.J.; Griffiths, C.E.; Kleyn, C.E. Does psychosocial stress play a role in the exacerbation of psoriasis? Br. J. Dermatol. 2013, 169, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Lonne-Rahm, S.B.; Rickberg, H.; El-Nour, H.; Mårin, P.; Azmitia, E.C.; Nordlind, K. Neuroimmune mechanisms in patients with atopic dermatitis during chronic stress. J. Eur. Acad. Dermatol. Venereol. 2008, 22, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.M.; Cruser, D.; Podawiltz, A.; Mummert, D.I.; Jones, H.; Mummert, M.E. Psychological stress and the cutaneous immune response: Roles of the hpa axis and the sympathetic nervous system in atopic dermatitis and psoriasis. Dermatol. Res. Pract. 2012, 2012, 403908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kossakowska, M.M.; Cieścińska, C.; Jaszewska, J.; Placek, W.J. Control of negative emotions and its implication for illness perception among psoriasis and vitiligo patients. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 429–433. [Google Scholar] [CrossRef] [PubMed]
- van den Boorn, J.G.; Jakobs, C.; Hagen, C.; Renn, M.; Luiten, R.M.; Melief, C.J.; Tüting, T.; Garbi, N.; Hartmann, G.; Hornung, V. Inflammasome-dependent induction of adaptive nk cell memory. Immunity 2016, 44, 1406–1421. [Google Scholar] [CrossRef] [Green Version]
- Bian, Y.; Yu, H.; Jin, M.; Gao, X. Repigmentation by combined narrow-band ultraviolet b/adipose-derived stem cell transplantation in the mouse model: Role of nrf2/ho-1-mediated Ca2+ homeostasis. Mol. Med. Rep. 2022, 25, 6. [Google Scholar] [CrossRef]
- Zhang, R.; Qiao, C.; Liu, Q.; He, J.; Lai, Y.; Shang, J.; Zhong, H. A reliable high-throughput screening model for antidepressant. Int. J. Mol. Sci. 2021, 22, 9505. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, J.; Lai, Y.; Zhang, X.; Yue, Y.; Zhong, H.; Shang, J. Optimization of Monobenzone-Induced Vitiligo Mouse Model by the Addition of Chronic Stress. Int. J. Mol. Sci. 2023, 24, 6990. https://doi.org/10.3390/ijms24086990
Dong J, Lai Y, Zhang X, Yue Y, Zhong H, Shang J. Optimization of Monobenzone-Induced Vitiligo Mouse Model by the Addition of Chronic Stress. International Journal of Molecular Sciences. 2023; 24(8):6990. https://doi.org/10.3390/ijms24086990
Chicago/Turabian StyleDong, Jing, Yifan Lai, Xiaofeng Zhang, Yunyun Yue, Hui Zhong, and Jing Shang. 2023. "Optimization of Monobenzone-Induced Vitiligo Mouse Model by the Addition of Chronic Stress" International Journal of Molecular Sciences 24, no. 8: 6990. https://doi.org/10.3390/ijms24086990
APA StyleDong, J., Lai, Y., Zhang, X., Yue, Y., Zhong, H., & Shang, J. (2023). Optimization of Monobenzone-Induced Vitiligo Mouse Model by the Addition of Chronic Stress. International Journal of Molecular Sciences, 24(8), 6990. https://doi.org/10.3390/ijms24086990