
“Pathomorphogenic” Changes Caused by Citrus Bark Cracking Viroid and Transcription Factor TFIIIA-7ZF Variants Support Viroid Propagation in Tobacco


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Splicing Variants of Viroid-Modulated TFIIIA: TFIIIA-7ZF from N. benthamiana and Humulus lupulus Support CBCVd Propagation in Transgenic Tobacco
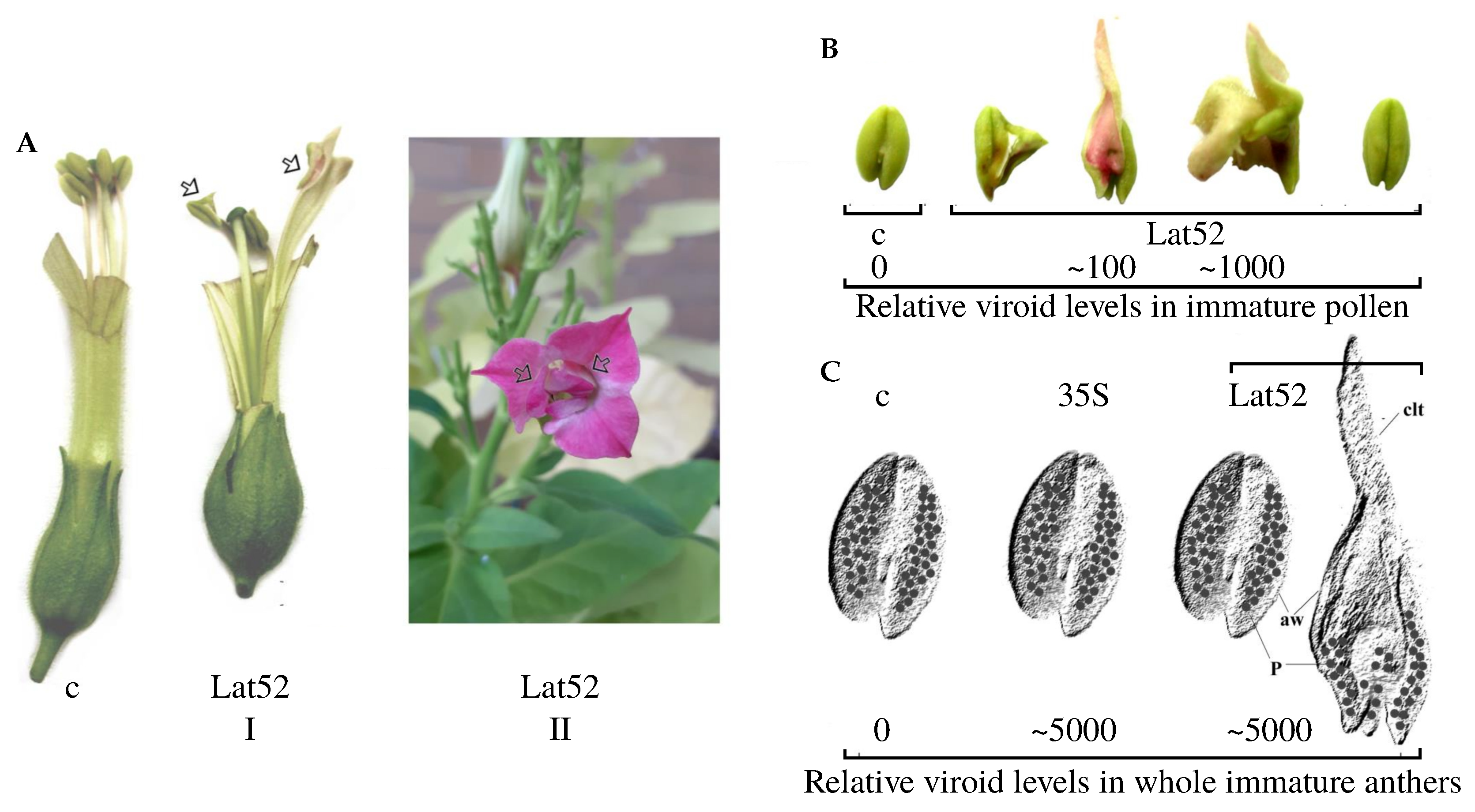
2.2. Specific Morphogenic Changes Caused by Elevated Levels of TFIIIA-7ZF and CBCVd
2.3. Transcriptome Profiling and Identification of Some Regulatory Factors Potentially Involved in CBCVd-Caused “Pathomorphogenesis”
3. Materials and Methods
3.1. Plant Cultivation Conditions, Plant Transformation, Preparation of Tobacco Hybrid Variants, and RNA Sampling
3.2. Viroid Quantification in CBCVd Transformants and Hybrids
3.3. Quantification of mRNA Levels of Selected Tobacco Genes Potentially Involved in Morphogenetic Changes and Cloning
3.4. NGS and Transcriptome Profiling
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sano, T. Progress in 50 years of viroid research—Molecular structure, pathogenicity, and host adaptation. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2021, 97, 371–401. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Vidalakis, G.; Sano, T. Economic significance of fruit tree and grapevine viroids. In Viroids and Satellites; Hadidi, A., Randles, J., Flores, R., Palukaitis, P., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017; pp. 15–21. [Google Scholar]
- Navarro, B.; Flores, R.; Di Serio, F. Advances in viroid-host interactions. Annu. Rev. Virol. 2021, 8, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Diener, T. (Ed.) The Viroids; Plenum Press: New York, NY, USA, 1987. [Google Scholar]
- Diener, T. Viroids and the nature of viroid diseases. Arch. Virol. Suppl. 1999, 15, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Flores, R.; Randles, J.; Semancik, J. (Eds.) Viroids; CSIRO Publishing: Clayton, Australia, 2003. [Google Scholar]
- Jakše, J.; Radišek, S.; Pokorn, T.; Matoušek, J.; Javornik, B. Deep-sequencing revealed Citrus bark cracking viroid (CBCVd) as a highly aggressive pathogen on hop. Plant Pathol. 2015, 64, 831–842. [Google Scholar] [CrossRef]
- Mishra, A.; Duraisamy, G.; Matoušek, J.; Radisek, S.; Javornik, B.; Jakse, J. Identification and characterization of microRNAs in Humulus lupulus using high-throughput sequencing and their response to Citrus bark cracking viroid (CBCVd) infection. BMC Genom. 2016, 17, 919. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Siglová, K.; Jakše, J.; Radišek, S.; Brass, J.; Tsushima, T.; Guček, T.; Duraisamy, G.; Sano, T.; Steger, G. Propagation and some physiological effects of Citrus bark cracking viroid and Apple fruit crinkle viroid in multiple infected hop (Humulus lupulus L.). J. Plant Physiol. 2017, 213, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Štajner, N.; Radišek, S.; Mishra, A.; Nath, V.; Matoušek, J.; Jakše, J. Evaluation of disease severity and global transcriptome response induced by citrus bark cracking viroid, hop latent viroid, and their co-infection in hop (Humulus lupulus L.). Int. J. Mol. Sci. 2019, 20, 3154. [Google Scholar] [CrossRef]
- Nath, V.; Mishra, A.; Kumar, A.; Matoušek, J.; Jakše, J. Revisiting the role of transcription factors in coordinating the defense response against citrus bark cracking viroid infection in commercial hop (Humulus Lupulus L.). Viruses 2019, 11, 419. [Google Scholar] [CrossRef]
- Nath, V.; Mishra, A.; Awasthi, P.; Shrestha, A.; Matoušek, J.; Jakse, J.; Kocábek, T.; Khan, A. Identification and characterization of long non-coding RNA and their response against citrus bark cracking viroid infection in Humulus lupulus. Genomics 2021, 113, 2350–2364. [Google Scholar] [CrossRef]
- Matoušek, J.; Steinbachová, L.; Drábková, L.; Kocábek, T.; Potěšil, D.; Mishra, A.; Honys, D.; Steger, G. Elimination of viroids from tobacco pollen involves a decrease in propagation rate and an increase in the degradation processes. Int. J. Mol. Sci. 2020, 21, 3029. [Google Scholar] [CrossRef]
- Matoušek, J.; Steger, G. The splicing variant TFIIIA-7ZF of viroid-modulated transcription factor IIIA causes physiological irregularities in transgenic tobacco and transient somatic depression of “degradome” characteristic for developing pollen. Cells 2022, 11, 784. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qu, J.; Ji, S.; Wallace, A.; Wu, J.; Li, Y.; Gopalan, V.; Ding, B. A land plant-specific transcription factor directly enhances transcription of a pathogenic noncoding RNA template by DNA-dependent RNA polymerase II. Plant Cell 2016, 28, 1094–1107. [Google Scholar] [CrossRef] [PubMed]
- Dissanayaka Mudiyanselage, S.; Ma, J.; Pechan, T.; Pechanova, O.; Liu, B.; Wang, Y. A remodeled RNA polymerase II complex catalyzing viroid RNA-templated transcription. PLoS Pathog. 2022, 18, e1010850. [Google Scholar] [CrossRef]
- Steinbachová, L.; Matoušek, J.; Steger, G.; Matoušková, H.; Radišek, S.; Honys, D. Transformation of seed non-transmissible hop viroids in Nicotiana benthamiana causes distortions in male gametophyte development. Plants 2021, 10, 2398. [Google Scholar] [CrossRef]
- Patzak, J.; Henychová, A.; Matoušek, J. Developmental regulation of lupulin gland-associated genes in aromatic and bitter hops (Humulus lupulus L.). BMC Plant Biol. 2021, 21, 534. [Google Scholar] [CrossRef] [PubMed]
- Jakše, J.; Wang, Y.; Matoušek, J. Transcriptomic analyses provide insights into plant-viroid interactions. In Fundamentals of Viroid Biology; Adkar-Purushothama, C., Sano, T., Perreault, J.P., Sreenivasa, M., DiSerio, F., Daròs, J.A., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Navarro, B.; Gisel, A.; Rodio, M.; Delgado, S.; Flores, R.; Di Serio, F. Small RNAs containing the pathogenic determinant of a chloroplast-replicating viroid guide the degradation of a host mRNA as predicted by RNA silencing. Plant J. 2012, 70, 991–1003. [Google Scholar] [CrossRef] [PubMed]
- Eiras, M.; Nohales, M.; Kitajima, E.; Flores, R.; Daròs, J. Ribosomal protein L5 and transcription factor IIIA from Arabidopsis thaliana bind in vitro specifically Potato spindle tuber viroid RNA. Arch. Virol. 2010, 156, 529–533. [Google Scholar] [CrossRef]
- Dadami, E.; Boutla, A.; Vrettos, N.; Tzortzakaki, S.; Karakasilioti, I.; Kalantidis, K. DICER-LIKE 4 but not DICER-LIKE 2 may have a positive effect on potato spindle tuber viroid accumulation in Nicotiana benthamiana. Mol. Plant 2012, 6, 232–234. [Google Scholar] [CrossRef]
- Minoia, S.; Carbonell, A.; Di Serio, F.; Gisel, A.; Carrington, J.; Navarro, B.; Flores, R. Specific argonautes selectively bind small RNAs derived from potato spindle tuber viroid and attenuate viroid accumulation in vivo. J. Virol. 2014, 88, 11933–11945. [Google Scholar] [CrossRef]
- Cavallini-Speisser, Q.; Morel, P.; Monniaux, M. Petal Cellular Identities. Front. Plant Sci. 2021, 12, 2430. [Google Scholar] [CrossRef]
- Martin, C.; Bhatt, K.; Baumann, K.; Jin, H.; Zachgo, S.; Roberts, K.; Schwarz-Sommer, Z.; Glover, B.; Perez-Rodrigues, M. The mechanics of cell fate determination in petals. Philos. Trans. R Soc. Lond. B Biol. Sci. 2002, 357, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rodriguez, M.; Jaffe, F.W.; Butelli, E.; Glover, B.J.; Martin, C. Development of three different cell types is associated with the activity of a specific MYB transcription factor in the ventral petal of Antirrhinum majus flowers. Development 2005, 132, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Galego, L.; Almeida, J. Role of DIVARICATA in the control of dorsoventral asymmetry in Antirrhinum flowers. Genes Dev. 2002, 16, 880–891. [Google Scholar] [CrossRef]
- Thomson, B.; Wellmer, F. Molecular regulation of flower development. Curr. Top. Dev. Biol. 2019, 131, 185–210. [Google Scholar] [CrossRef]
- Ali, S.; Khan, N.; Xie, L. Molecular and hormonal regulation of leaf morphogenesis in Arabidopsis. Int. J. Mol. Sci. 2020, 21, 5132. [Google Scholar] [CrossRef] [PubMed]
- Guan, C.; Jiao, Y. Interplay between the shoot apical meristem and lateral organs. Abiotech 2020, 1, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Moulia, B.; Badel, E.; Bastien, R.; Duchemin, L.; Eloy, C. The shaping of plant axes and crowns through tropisms and elasticity: An example of morphogenetic plasticity beyond the shoot apical meristem. New Phytol. 2022, 233, 2354–2379. [Google Scholar] [CrossRef]
- Dodsworth, S. A diverse and intricate signalling network regulates stem cell fate in the shoot apical meristem. Dev. Biol. 2009, 336, 1–9. [Google Scholar] [CrossRef]
- Xu, Q.; Li, R.; Weng, L.; Sun, Y.; Li, M.; Xiao, H. Domain-specific expression of meristematic genes is defined by the LITTLE ZIPPER protein DTM in tomato. Commun. Biol. 2019, 2, 134. [Google Scholar] [CrossRef]
- Wenkel, S.; Emery, J.; Hou, B.; Evans, M.; Barton, M. A feedback regulatory module formed by LITTLE ZIPPER and HD-ZIPIII genes. Plant Cell 2007, 19, 3379–3390. [Google Scholar] [CrossRef]
- Yin, X.; Meicenheimer, R. Anisotomous dichotomy results from an unequal bifurcation of the original shoot apical meristem in Diphasiastrum digitatum (Lycopodiaceae). Am. J. Bot. 2017, 104, 782–786. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, S.; Shchennikova, A.; Franken, J.; Immink, R.; Angenent, G. Control of floral meristem determinacy in petunia by MADS-box transcription factors. Plant Physiol. 2006, 140, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Danilevskaya, O.; Meng, X.; Selinger, D.; Deschamps, S.; Hermon, P.; Vansant, G.; Gupta, R.; Ananiev, E.; Muszynski, M. Involvement of the MADS-box gene ZMM4 in floral induction and inflorescence development in maize. Plant Physiol. 2008, 147, 2054–2069. [Google Scholar] [CrossRef]
- Pérez-Ruiz, R.; García-Ponce, B.; Marsch-Martínez, N.; Ugartechea-Chirino, Y.; Villajuana-Bonequi, M.; de Folter, S.; Azpeitia, E.; Dávila-Velderrain, J.; Cruz-Sánchez, D.; Garay-Arroyo, A.; et al. XAANTAL2 (AGL14) is an important component of the complex gene regulatory network that underlies Arabidopsis shoot apical meristem transitions. Mol. Plant 2015, 8, 796–813. [Google Scholar] [CrossRef]
- Li, Y.; Feng, J.; Cheng, L.; Dai, C.; Gao, Q.; Liu, Z.; Kang, C. Gene expression profiling of the shoot meristematic tissues in woodland strawberry Fragaria vesca. Front. Plant Sci. 2019, 10, 1624. [Google Scholar] [CrossRef]
- Byrne, M.; Groover, A.; Fontana, J.; Martienssen, R. Phyllotactic pattern and stem cell fate are determined by the Arabidopsis homeobox gene BELLRINGER. Development 2003, 130, 3941–3950. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.; Hake, S. The interaction of two homeobox genes, Brevipedicellus and Pennywise, regulates internode patterning in the Arabidopsis inflorescence. Plant Cell 2003, 15, 1717–1727. [Google Scholar] [CrossRef]
- Bhatt, A.; Etchells, J.; Canales, C.; Lagodienko, A.; Dickinson, H. VAAMANA—A BEL1-like homeodomain protein, interacts with KNOX proteins BP and STM and regulates inflorescence stem growth in Arabidopsis. Gene 2004, 328, 103–111. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, F.; Wang, R.; Zhou, X.; Sze, S.; Liou, L.; Barefoot, A.; Dickman, M.; Zhang, X. Arabidopsis Argonaute10 specifically sequesters miR166/165 to regulate shoot apical meristem development. Cell 2011, 145, 242–256. [Google Scholar] [CrossRef]
- Kano-Murakami, Y.; Yanai, T.; Tagiri, A.; Matsuoka, M. A rice homeotic gene, OSH1, causes unusual phenotypes in transgenic tobacco. FEBS Lett. 1993, 334, 365–368. [Google Scholar] [CrossRef]
- Gou, X.; He, K.; Yang, H.; Yuan, T.; Lin, H.; Clouse, S.; Li, J. Genome-wide cloning and sequence analysis of leucine-rich repeat receptor-like protein kinase genes in Arabidopsis thaliana. BMC Genom. 2010, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, X.; Li, M.; He, P.; Zhang, Y. Loss-of-function of Arabidopsis receptor-like kinase BIR1 activates cell death and defense responses mediated by BAK1 and SOBIR1. New Phytol. 2016, 212, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Wang, X.; Wang, D.; Xu, F.; Ding, X.; Zhang, Z.; Bi, D.; Cheng, Y.; Chen, S.; Li, X.; et al. Regulation of cell death and innate immunity by two receptor-like kinases in Arabidopsis. Cell. Host Microbe 2009, 6, 34–44. [Google Scholar] [CrossRef]
- Ito, J.; Fukuda, H. ZEN1 is a key enzyme in the degradation of nuclear DNA during programmed cell death of tracheary elements. Plant Cell 2002, 14, 3201–3211. [Google Scholar] [CrossRef]
- Pérez-Amador, M.; Abler, M.; De Rocher, E.; Thompson, D.; van Hoof, A.; LeBrasseur, N.; Lers, A.; Green, P. Identification of BFN1, a bifunctional nuclease induced during leaf and stem senescence in Arabidopsis. Plant Physiol. 2000, 122, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Farage-Barhom, S.; Burd, S.; Sonego, L.; Mett, A.; Belausov, E.; Gidoni, D.; Lers, A. Localization of the Arabidopsis senescence- and cell death-associated BFN1 nuclease: From the ER to fragmented nuclei. Mol. Plant 2011, 4, 1062–1073. [Google Scholar] [CrossRef]
- Zong, Y.; Hao, Z.; Tu, Z.; Shen, Y.; Zhang, C.; Wen, S.; Yang, L.; Ma, J.; Li, H. Genome-wide survey and identification of AP2/ERF genes involved in shoot and leaf development in Liriodendron chinense. BMC Genom. 2021, 22, 807. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Yuan, L. ERF Gene Clusters: Working Together to Regulate Metabolism. Trends Plant Sci. 2021, 26, 23–32. [Google Scholar] [CrossRef]
- Phukan, U.; Jeena, G.; Tripathi, V.; Shukla, R. Regulation of Apetala2/Ethylene Response Factors in Plants. Front. Plant Sci. 2017, 8, 150. [Google Scholar] [CrossRef]
- Mirzaei, K.; Bahramnejad, B.; Fatemi, S. Genome-wide identification and characterization of the bZIP gene family in potato (Solanum tuberosum). Plant Gene 2020, 24, 100257. [Google Scholar] [CrossRef]
- Herath, V.; Verchot, J. Insight into the bZIP gene family in Solanum tuberosum: Genome and transcriptome analysis to understand the roles of gene diversification in spatiotemporal gene expression and function. Int. J. Mol. Sci. 2020, 22, 253. [Google Scholar] [CrossRef] [PubMed]
- Irish, V. The flowering of Arabidopsis flower development. Plant J. 2010, 61, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Valdés, A.; Overnäs, E.; Johansson, H.; Rada-Iglesias, A.; Engström, P. The homeodomain-leucine zipper (HD-Zip) class I transcription factors ATHB7 and ATHB12 modulate abscisic acid signalling by regulating protein phosphatase 2C and abscisic acid receptor gene activities. Plant Mol. Biol. 2012, 80, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Donnelly, L.; Sun, D.; Rao, J.; Reid, M.; Jiang, C. A Petunia homeodomain-leucine zipper protein, PhHD-Zip, plays an important role in flower senescence. PLoS ONE 2014, 9, e88320. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, L.; Zhang, X.; Zhang, S.; Xie, D.; Liang, C.; Huang, W.; Fan, L.; Fang, Y.; Chang, Y. OFP1 Interaction with ATH1 Regulates Stem Growth, Flowering Time and Flower Basal Boundary Formation in Arabidopsis. Genes 2018, 9, 399. [Google Scholar] [CrossRef]
- Tupý, J.; Süss, J.; Hrabětová, E.; Říhova, L. Developmental changes in gene expression during pollen differentiation and maturation in Nicotiana tabacum L. Biol. Plant. 1983, 25, 231. [Google Scholar] [CrossRef]
- Horsch, R.; Fry, J.; Hoffman, N.; Eichholtz, D.; Rogers, S.; Fraley, R. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar] [CrossRef]
- Pfaffl, M. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Matoušek, J.; Kozlová, P.; Orctová, L.; Schmitz, A.; Pešina, K.; Bannach, O.; Diermann, D.; Steger, G.; Riesner, D. Accumulation of viroid-specific small RNAs and increase in nucleolytic activities linked to viroid-caused pathogenesis. Biol. Chem. 2007, 388, 1–13. [Google Scholar] [CrossRef]
- Puchta, H.; Ramm, K.; Sänger, H. The molecular structure of hop latent viroid (HLV), a new viroid occurring worldwide in hops. Nucleic Acids Res. 1988, 16, 4197–4216. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.; Park, C.; Bennett, C.; Salzberg, S. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.; Leek, J.; Salzberg, S. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Flexible analysis of transcriptome assemblies with Ballgown. Biorxiv 2014, 1, 003665. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef]
- Edwards, K.D.; Fernandez-Pozo, N.; Drake-Stowe, K.; Humphry, M.; Evans, A.D.; Bombarely, A.; Allen, F.; Hurst, R.; White, B.; Kernodle, S.P.; et al. A reference genome for Nicotiana tabacum enables map-based cloning of homeologous loci implicated in nitrogen utilization efficiency. BMC Genom. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Matoušek, J.; Junker, V.; Vrba, L.; Schubert, J.; Patzak, J.; Steger, G. Molecular characterization and genome organization of 7 SL RNA genes from hop (Humulus lupulus L.). Gene 1999, 239, 173–183. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matoušek, J.; Wüsthoff, K.P.; Steger, G. “Pathomorphogenic” Changes Caused by Citrus Bark Cracking Viroid and Transcription Factor TFIIIA-7ZF Variants Support Viroid Propagation in Tobacco. Int. J. Mol. Sci. 2023, 24, 7790. https://doi.org/10.3390/ijms24097790
Matoušek J, Wüsthoff KP, Steger G. “Pathomorphogenic” Changes Caused by Citrus Bark Cracking Viroid and Transcription Factor TFIIIA-7ZF Variants Support Viroid Propagation in Tobacco. International Journal of Molecular Sciences. 2023; 24(9):7790. https://doi.org/10.3390/ijms24097790
Chicago/Turabian StyleMatoušek, Jaroslav, Kevin P. Wüsthoff, and Gerhard Steger. 2023. "“Pathomorphogenic” Changes Caused by Citrus Bark Cracking Viroid and Transcription Factor TFIIIA-7ZF Variants Support Viroid Propagation in Tobacco" International Journal of Molecular Sciences 24, no. 9: 7790. https://doi.org/10.3390/ijms24097790
APA StyleMatoušek, J., Wüsthoff, K. P., & Steger, G. (2023). “Pathomorphogenic” Changes Caused by Citrus Bark Cracking Viroid and Transcription Factor TFIIIA-7ZF Variants Support Viroid Propagation in Tobacco. International Journal of Molecular Sciences, 24(9), 7790. https://doi.org/10.3390/ijms24097790