1. Introduction
Progesterone is one of the key steroid hormones whose task is to regulate female reproductive functions, largely controlled in many organs such as the uterus, ovaries, mammary glands, and brain [
1]. Progesterone is synthesized mainly in the ovaries, placenta, and adrenal glands [
2]. Apart from securing the normal course of pregnancy, progesterone also controls most reproductive processes in women, including oocyte maturation, ovulation, menstruation, facilitating the implantation of the embryo in the uterus, uterine growth, the inhibition of myometrial contractions, the development of the mammary gland, the regulation of milk production, and sexual behavior [
1,
3].
Although the role of progesterone in humans is well known, the possible activity and implications of progesterone metabolites have yet to be determined. There is increasing evidence that many metabolites are not inactive but have important biological effects [
4,
5,
6]. Progesterone is rapidly and extensively metabolized in the human body, mainly in the liver, but also in the brain, skin, and various other extrahepatic tissues [
7,
8]. Progesterone exhibits a remarkably short elimination half-life of approximately 5 min in the circulatory system. The metabolism of progesterone is intricate, giving rise to a potential formation of up to 35 distinct unconjugated metabolites upon oral ingestion [
9]. The primary metabolic pathway of progesterone involves a reduction by 5α-reductase and 5β-reductase, leading to the formation of the dihydrogenated products, namely 5α-dihydroprogesterone and 5β-dihydroprogesterone, respectively [
10,
11,
12].
In the human organism a relatively small amount of progesterone (
1) is hydroxylated by 17α-hydroxylase (CYP17A1) and 21-hydroxylase (CYP21A2) to become 17α-hydroxyprogesterone and 21-hydroxyprogesterone, respectively [
13,
14]. Even smaller amounts of progesterone can also be hydroxylated by 11β-hydroxylase (CYP11B1), and to a lesser extent by aldosterone synthase (CYP11B2), into 11β-hydroxyprogesterone [
15]. Moreover, progesterone can be hydroxylated in the human liver by other cytochrome P450 enzymes that are not specific to steroids [
16]. 6β-Hydroxylation, which is catalyzed mainly by CYP3A4, is the main transformation and accounts for approximately 70% of the progesterone metabolism mediated by this enzyme [
16]. Other pathways include 6α-, 16α-, and 16β-hydroxylation [
13].
Over the course of more than 70 years of research, it has been observed that the biotransformation of progesterone by filamentous fungi can proceed in two main directions: hydroxylation or Baeyer–Villiger oxidation, and rarely does a combination of these processes occur [
17,
18,
19]. Hydroxylation predominantly occurs at the 6β and/or 11α positions; however, fungal cultures have also demonstrated the capability for hydroxylation at the 14α, 7α, 7β, 11β, 17α, 9α, 8β, 16α, 15α, 15β, 21, and 12β positions [
20,
21,
22]. The Baeyer–Villiger oxidation of progesterone is possible through the biotransformation by some
Aspergillus or
Penicillium species [
23,
24,
25,
26].
The introduction of a double bond between the C1 and C2 positions in the steroid structure has been found to enhance the metabolic stability against enzymes such as 5α-reductase and aromatase [
27,
28]. Notably, even in the early stages of research, it was observed that the medicinal properties of several Δ
1-3-ketosteroids were more potent compared to their corresponding 1(2)-saturated analogues [
29,
30]. This also refers to currently used medications. For instance, the anti-inflammatory efficacy of prednisolone is four times greater than that of hydrocortisone, and prednisone is five times more potent than cortisone [
31]. Δ
1-3-Ketosteroids demonstrate diverse essential biological activities, contributing to their widespread use in medicine and establishing a significant presence in the pharmaceutical industry [
28].
The primary objective of this study was to delineate the spectrum of hydroxylated derivatives arising from the bioconversion of pregn-1,4-diene-3,20-dione (Δ
1-progesterone) (
2). In addition, our aim was to investigate the potential biological activities associated with the selected hydroxy derivatives, utilizing state-of-the-art cheminformatic tools. As a result of the biotechnological functionalization, we obtained new Δ
1-3-ketopregnanes with high potential biological activity. The substrate (
2) was obtained by increasing the scale of the previously described method utilizing cholest-4-en-3-one Δ
1-dehydrogenase (AcmB) from
Sterolibacterium denitrificans, a key enzyme of the central degradation pathway of cholesterol [
32]. Further, biocatalysts of hydroxylation were used including strains of entomopathogenic filamentous fungi. The
Isaria farinosa KCh KW 1.1 strain possesses the ability to effectively dihydroxylate progesterone and its derivatives [
17,
33].
Isaria fumosorosea KCh J2 is known for its ability to carry out multienzymatic transformations of steroid compounds [
34,
35]. We also decided to use the
Beauveria bassiana KCh J1.5 strain, which is a representative of the species of entomopathogenic filamentous fungi most often used for the biotransformation of steroid compounds [
36,
37,
38,
39] and the
Beauveria caledonica KCh J3.3 strain belonging to the same genus [
36]. Two strains of species with relatively little known catalytic abilities were also used as biocatalysts:
Metarhizium anisopliae, capable of hydroxylating 13-ethyl-gon-4-ene-3,17-dione [
40], and
Isaria tenuipes, known as the zearalenone biocatalyst [
41].
2. Results and Discussion
The underlying aim of this research was the verification of the ability of six strains of entomopathogenic filamentous fungi (belonging to six different species) to transform progesterone with an additional double bond located between the C-1 and C-2 carbon atoms and to obtain new derivatives of pregn-1,4-diene-3,20-dione (Δ
1-progesterone) (
2). The substrate (
2) (Spectral data presented in
Supplementary Materials Figures S3–S7) was obtained by carrying out an enzymatic 1,2-dehydrogenation on an increased scale (3.5 g/L) using a recombinant cholest-4-en-3-one Δ
1-dehydrogenase (AcmB) from
Sterolibacterium denitrificans [
32] and phenazine methosulfate as an electron acceptor. The 1(2)-dehydrogenation proceeded efficiently under anaerobic conditions and a 100% conversion of
1 to
2 was obtained after 5.5 h of reaction. As biocatalysts for the subsequent transformations, the following strains of entomopathogenic filamentous fungi were used:
Beauveria bassiana KCh J1.5,
Beauveria caledonica KCh J3.3,
Isaria fumosorosea KCh J2,
Isaria farinosa KCh KW1.1, and
Isaria tenuipes MU35,
Metarhizium robertsii MU4 (formerly known as
Metarhizium anisopliae [
42]). Based on TLC and GC analyses, it was observed that the substrate was effectively transformed in the cultures of the tested strains (
Table 1). The resulting products are more polar than the substrate (
2), and the composition of the product mixture in the cultures of most of the tested biocatalysts varies during incubation.
In the culture of the
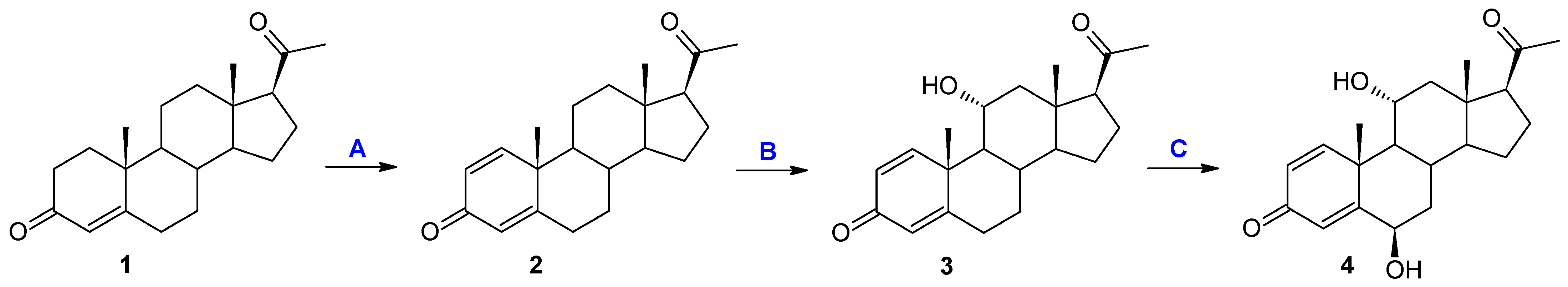
Metarhizium robertsii MU4 strain, one main product was obtained accompanied by a compound formed after 7 days of the transformation process in a small amount (approximately 4%). Based on TLC and GC analyses, after 1 and 3 days of biotransformation, compound
3 was identified as the only product, with a complete reaction of the substrate (
Figure 1). After the biotransformation on an increased scale, it was isolated with high efficiency—90%. After performing NMR analyses, the structure of this compound was determined to be 11α-hydroxypregn-1,4-diene-3,20-dione (
3). In the
1H NMR spectrum performed for this compound, a characteristic signal was observed at 4.07 ppm. The HSQC correlation spectrum shows the coupling of this signal with the carbon signal located at 67.97 ppm. The positions of both signals indicate that the obtained compound is a hydroxylation product. Slight shifts of the remaining signals, both in the
1H NMR and
13C NMR spectra (
Table 2), clearly indicate that one hydroxyl group has been introduced into the structure of the analyzed compound. Based on the analysis of the couplings of the signal at 4.07 ppm with the signals originating from the H-9, H-12α, and H-12β protons visible in the COSY spectrum and the couplings of this signal with the signals originating from the C-9, C-10, and C-12 carbons, the structure of the obtained compound can be unambiguously determined. The chemical shift and multiplicity of the signal originating from the proton H-11β unambiguously indicate the introduction of the hydroxyl group at the 11α position (
Supplementary Materials: Figures S10–S14).
The 11α-hydroxylation is the most frequently reported transformation of progesterone and its derivatives during biotransformation in cultures of filamentous fungi [
20,
43,
44,
45,
46,
47,
48,
49,
50]. However, in the vast majority of cases, apart from the 11α-hydroxy derivative, other products were also identified [
20,
45,
46,
47,
48,
49,
50]. In the culture of the
M. robertsii MU4 strain, after the first and third day of transformation, it is possible to obtain only one compound with very high efficiency. Due to the small amounts of formation of the second product, the structure of compound
4 was determined as a result of the preparative biotransformation in the culture of the
Beauveria bassiana KCh J1.5 strain. In the culture of this strain, after one day of biotransformation (analogously to the culture of the
M. robertsii MU4 strain), one main product—11α-hydroxypregn-1,4-diene-3,20-dione (
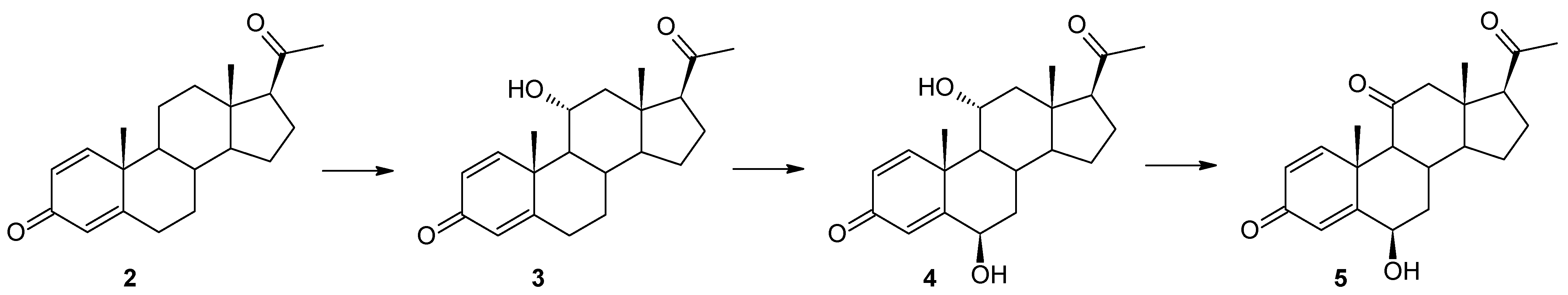
3)—was observed. However, after just three days of the transformation process, a dynamic change in the composition of the products was observed. Three products (
3,
4, and
5) were visible in the reaction mixture, and after the seventh day only compounds
4 and
5 were identified (
Table 1 and
Figure 2). Based on the course of the biotransformation in the culture of the
B. bassiana KCh J1.5 strain over time, it can be assumed that the transformations of the tested compound by
M. robertsii MU4 are of a cascade nature (
Figure 1). These assumptions were confirmed by the analysis of the structure of the resulting products. The
1H NMR spectrum obtained for compound
4 shows signals indicating the preservation of the basic skeleton of pregn-1,4-diene-3,20-dione. A characteristic signal at 3.93 ppm is also visible (as for product
3), which in the COSY spectrum couples with signals coming from the H-9, H-12α, and H-12β protons. The
13C NMR spectrum shows two signals in a position characteristic of signals coming from carbon atoms connected with the hydroxyl group: 66.37 ppm coming from carbon C-11 and the second in the position of 72.10 ppm. The latter signal couples in the HMBC spectrum with signals coming from the H-4 proton and, weakly, from the H-1 proton, and in the COSY spectrum with signals coming from H-7α and H-7β protons. Additionally, the characteristic multiplicity and shifts of the methyl groups at C-18, C-19, and C-21 clearly indicate that in the structure of the isolated compound of the hydroxyl group is also located in the 6β position. On this basis, compound
4 was characterized as 6β,11α-dihydroxypregn-1,4-diene-3,20-dione (
4) (
Supplementary Materials: Figures S17–S21). Based on the
1H NMR spectrum obtained for compound
5, it was found that it possesses the structure of pregn-1,4-diene-3,20-dione and (like compound
4) a hydroxyl group at carbon C-6. However, based on the
13C NMR spectrum (
Table 2), it was determined that it also contains three carbonyl groups in its structure. A clear shift towards a lower field of the signal coming from the protons of the H-19 methyl group and the analysis of the biotransformation of compound
2 in the culture of the
B. bassiana KCh J1.5 strain (compound
5 is formed from compound
4) allow us to clearly determine the structure of this product to be 6β-hydroxypregn-1,4-diene-3,11,20-trione (
5) (
Supplementary Materials: Figures S24–S28). On this basis, the course of the transformations of the tested substrate in the culture of the
B. bassiana KCh J1.5 strain can be determined as follows: the first stage is the hydroxylation leading to the 11α-hydroxyl derivative (
3), which undergoes further hydroxylation leading to the 6β,11α-dihydroxy derivative, and the last step is the oxidation of the hydroxyl group at carbon C-11, leading to the formation of compound
5 (
Figure 2). An analogous cascade course was previously described during the biotransformation of progesterone leading to 6β-hydroxy-11-oxoprogesterone in cultures of
Mucor M881 strains [
51] and
Isaria farinosa KCh KW1.1 [
17,
33]. On the other hand, the ability to dihydroxylate progesterone leading to the 6β,11α-dihydroxy derivative was previously observed in cultures of the following strains:
Aspergillus nidulans VKPM F-1069 [
45],
A. niger N402 [
43],
A. ochraceus [
49],
Rhizomucor pusillus and
Absidia griseolla var.
igachii [
48],
Rhizomucor tauricus IMI23312 [
50],
Cephalosporium aphidicola [
46], and
Trichoderma koningii,
T. hamatum,
T. aureoviride, and
T. pseudokoningii [
20].
A similar course was observed in the culture of the
Beauveria caledonica KCh J3.3 strain, although, only in the culture of this strain, an unreacted substrate was observed after one day (almost 30% of the reaction mixture). However, after the third day of the transformation process, in addition to the hydroxylation products 11α and 6β,11α, other products were also observed. Unfortunately, their large number and individual percentages did not allow for their isolation and a determination of their structure. Additionally, a progressive decrease over time in the identified compounds can be observed, which may indicate degradation processes taking place in the cells of the
Beauveria caledonica KCh J3.3 strain. An even greater number of products formed were observed during the incubation of pregn-1,4-diene-3,20-dione (Δ
1-progesterone) (
2) in the culture of the
Isaria tenuipes MU35 strain. After just one day, the reaction of the substrate was observed to be complete and a multitude of products were observed both on TLC and GC. Due to the large number of compounds formed, attempts to isolate them were not pursued. Nevertheless, the obtained result indicates that the tested biocatalyst produces a multitude of enzymes capable of transforming steroid compounds and indicates the possibility of using this strain in the biocatalysis of various bioactive compounds; e.g., in our previous work, we described the effective glycosylation of quercetin by
Isaria tenuipes MU35 [
52]. It was also proven that a strain of this species is capable of an effective metabolism of zearalenone and, similar to the two representatives of this genus more popular in biocatalysis (
I. fumosorosea,
I. farinosa), multienzymatic transformations of the used substrate were observed [
41]. Moreover,
Isaria tenuipes are important entomopathogenic fungi used in health foods and traditional herbal medicines in East Asia, thus a source of valuable metabolites [
53,
54].
The
Isaria fumosorosea KCh J2 strain has already been described in previous manuscripts as a biocatalyst capable of producing many effective enzymes involved in the transformation of the substrates used. It is known for its effective cascade reactions of flavonoid compounds (demethylation, hydroxylation, oxidation of hydroxyl groups, methylglycosidation) [
55], steroid compounds—dehydroepiandrosterone (DHEA) (hydroxylation, Baeyer–Villiger oxidation) [
34]—and estrones (hydroxylation, oxidation of hydroxyl groups, methylglycosidation, Baeyer–Villiger oxidation) [
35]. However, the high activity of these many enzymes led to the formation of many products during the biotransformation of progesterone, which consequently prevented their separation [
34]. During the transformation of pregn-1,4-diene-3,20-dione (
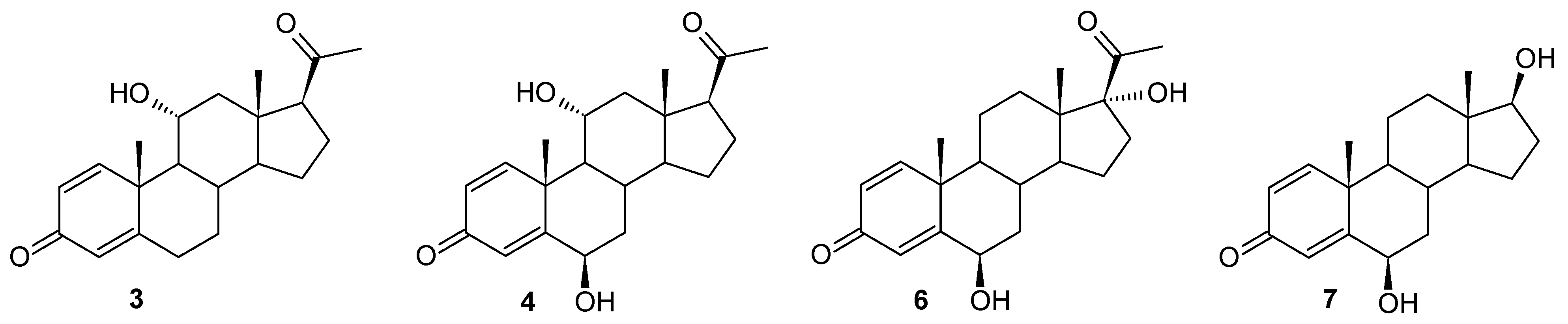
2) in the culture of this strain, dynamic transformations of the resulting compounds were also observed, but in this case it was possible to determine the structure of most of the resulting metabolites. In the culture of the
I. fumosorosea KCh J2 strain, a hydroxylation effect was observed, leading to 11α-hydroxypregn-1,4-diene-3,20-dione (
3) and 6β,11α-dihydroxypregn-1,4-diene-3,20-dione (
4)—a transformation pathway observed in the cultures of previously described strains of entomopathogenic fungi (
B. bassiana KCh J1.5,
B. caledonica KCh J3.3, and
M. robertsii MU4). However, in parallel, the following were identified as the dominant products: 6β,17α-dihydroxypregn-1,4-diene-3,20-dione (
6) and 6β,17β-dihydroxyandrost-1,4-diene-3-one (
7) (
Figure 3). It was observed in the
1H NMR spectrum that the skeleton of pregn-1,4-diene-3,20-dione in compound
6 was retained. The presence of a signal at 4.53 ppm indicates the location of a hydroxyl group present at the secondary carbon atom. The presence of this functionalization is confirmed by the coupling of this signal with the signal located at 74.00 ppm, visible in the HMQC spectrum. Additionally, this signal visible in the
13C NMR spectrum is coupled in the HMBC spectrum with the signal coming from the H-4 proton, which indicates that the C-6 carbon atom has been hydroxylated. The position and multiplicity of the signal visible in the
1H NMR spectrum and the position of the C-18 and C-19 methyl groups indicate that the hydroxyl group is located in the β position. Additionally, the shift of the C-21 methyl group towards a higher field indicates a structural modification of compound
6 within the D ring. The location of the signal coming from the C-17 carbon at 89.79 ppm (
13C NMR spectrum) and the coupling of this signal with the signal coming from the C-18 methyl protons visible on the HMBC spectrum indicate that carbon atom C-17 has undergone hydroxylation as well. Compound
6 is therefore a dihydroxy derivative of the substrate used (
Supplementary Materials: Figures S31–S35).
Analyzing the course of the biotransformation of pregn-1,4-diene-3,20-dione (
2) in the culture of the
I. fumosorosea KCh J2 strain, one can notice that the percentage of 6β,17α-dihydroxypregn-1,4-diene-3,20-dione (
6) decreases during biotransformation, while the amount of compound
7 increases (exceeding the value of 50% after seven days of reaction). This indicates a high probability that compound
6 is an intermediate during the formation of compound
7 (
Table 1). Based on the insight provided by the NMR spectra, it was determined that, compared to compound
6, in the structure of compound
7, the structural modification occurred only within the D ring. The absence of a signal in the
1H NMR and
13C NMR spectra coming from the C-21 methyl group and the presence of a triplet at 3.64 ppm (coming from proton H-17α visible on the
1H NMR), which couples with the signal coming from carbon C-17 in the HMQC spectrum, clearly indicate that the obtained product is 6β,17β-dihydroxyandrost-1,4-diene-3-one (
7), i.e., 6β-hydroxy-Δ
1-testosterone (
Supplementary Materials: Figures S31–S35). Similar mechanisms are known in the available literature, involving subsequent oxidations leading to the degradation of the acyl substituent located at the C-17 carbon of progesterone. Over the past decades, the microbial production of 17α-hydroxyprogesterone (17-HP) using specific strains (e.g.,
Curvularia lunata ATCC 12017,
Cunninghamella blakesleeana ATCC 8688a, and
Cladosporium sp. F 5394) [
56] has attracted increasing attention due to its environmental benefits, flexible production conditions and high catalytic efficiency [
57]. During hydroxylation, the progesterone-17α-hydroxylase belonging to cytochrome P450 monooxygenases (CYP) converts progesterone to 17-HP [
57,
58]. In mammals, including humans, there is only one CYP17A (CYP17A1), which plays a key role in the oxidation of progesterone to androstenedione [
59,
60]. CYP17A1 catalyzes two different oxidase reactions of a mixed function: the first step is the hydroxylation reaction at the C17 position of progesterone to produce 17-HP, followed by the cleavage of the C17,20 bond to produce testosterone and androstenedione [
60,
61]. On the other hand, the 17α-hydroxylation reaction of progesterone and its derivatives is one of the most desirable transformations of steroid compounds leading to valuable products [
57]. The 17α-hydroxyprogesterone caproate can be used to prevent premature birth, while 17-HP is used to treat amenorrhea, endometrial cancer, abnormal uterine bleeding, and premature birth [
57,
62,
63].
In our previous manuscripts, we described the ability of the
Isaria farinosa KCh KW 1.1 strain to hydroxylate progesterone and its derivatives effectively, leading to 6β,11α-dihydroxy derivatives [
17,
33]. These metabolites were obtained during the transformation of progesterone, 11α-hydroxyprogesterone, and 16α,17α-epoxyprogesterone [
17,
33]. However, during the transformation of 17α-hydroxyprogesterone, we isolated three hydroxy derivatives: 6β,17α-dihydroxyprogesterone, 12β,17α-dihydroxyprogesta-1,4-diene-3-one, and 6β,12β,17α-trihydroxyprogesterone [
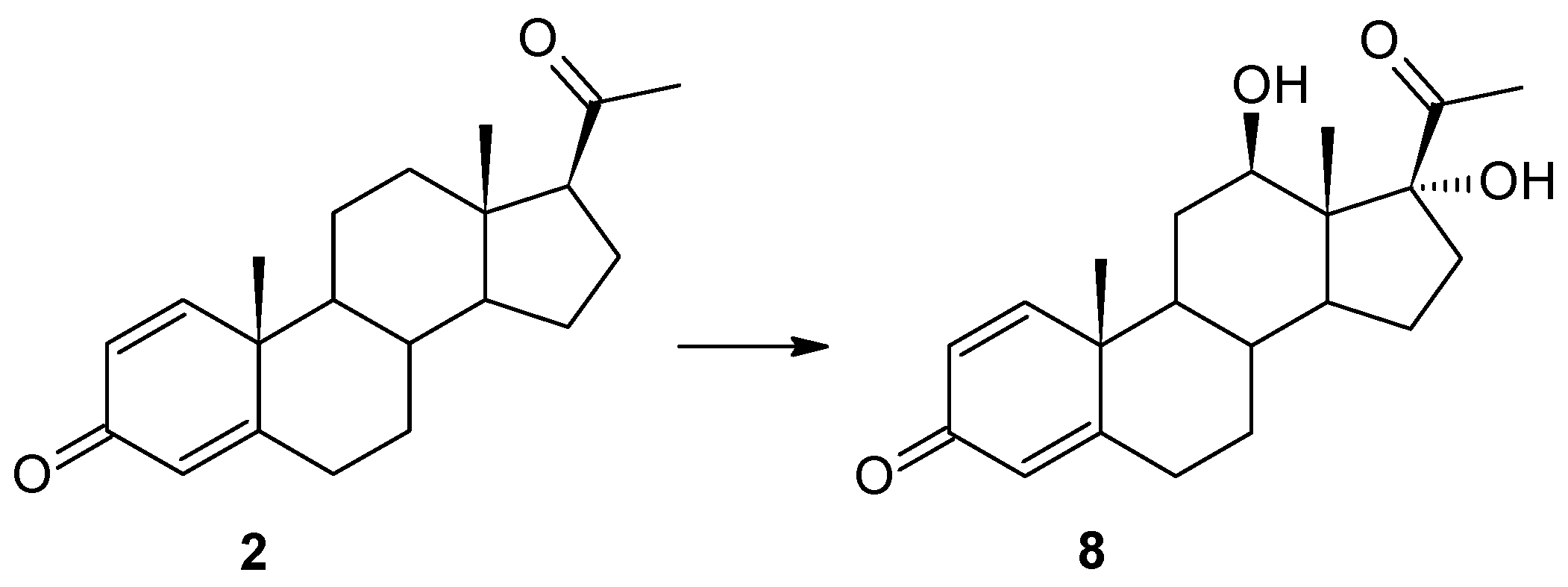
17]. Surprisingly, as a result of using the progesterone with an additional double bond between the C1 and C-2 carbon atoms (Δ
1-progesterone (
2)) as a substrate, 12β,17α-dihydroxypregn-1,4-diene-3,20-dione (
8) was obtained in the culture of this biocatalyst. In the
1H NMR spectrum, a characteristic position of the signal originating from the protons of the H-21 methyl group was observed: 2.26 ppm, indicating the hydroxylation of carbon C-17. The shift of the signal from this carbon towards the lower field to a value of 88.55 ppm confirms this assumption. The presence of a signal at 68.04 ppm in the
13C NMR spectrum, and the coupling of this signal (HMQC spectrum) with the signal visible at 3.86 ppm in the
1H NMR spectra, is evidence of the hydroxylation of the secondary carbon atom as well. The coupling visible in the HMBC spectrum of signals from both protons of the hydroxyl groups with the signal from carbon C-13 indicates that hydroxylation occurred at carbons C-12 and C-17. The shapes and positions of all the signals indicate that the isolated compound is 12β,17α-dihydroxypregn-1,4-diene-3,20-dione (
8) (
Supplementary Materials: Figures S40–S44). After a shorter biotransformation time, other steroid products were also observed in the culture of the
Isaria farinosa KCh KW 1.1 strain. However, it was not possible to isolate them. Based on the structure of compound
8, it can be assumed that two of them may be intermediate products leading to 12β,17α-dihydroxypregn-1,4-diene-3,20-dione (
8). Their structure is shown in
Figure 4. Based on the results obtained, both the
Isaria farinosa KCh KW 1.1 and
Isaria fumosorosea KCh J2 strains can be qualified for further research on the activity of monooxygenases, with a particular emphasis on CYP17A.
To evaluate the physicochemical properties, pharmacokinetics, and potential biological activity of Δ
1-progesterone derivatives, cheminformatic tools such as SwissADME and passOnline were employed. Physicochemical descriptor calculations were conducted for both the substrate and the obtained products (
2–
8), as summarized in
Table 3. Predictions encompassed their ADME parameters (absorption, distribution, metabolism, and excretion), pharmacokinetic properties, and suitability for medicinal chemistry. The analysis utilized the online tool SwissADME, accessible at
http://www.swissadme.ch (accessed on 15 June 2023), developed and managed by the Molecular Modeling Group of the Swiss Institute of Bioinformatics (SIB) [
64]. According to the results obtained from this tool, it was observed that all the products exhibited significantly lower lipophilicity and markedly higher water solubility compared to Δ
1-progesterone (
2) (
Table 3). This shift is attributed to the introduction of polar -OH groups during the transformation.
According to the results we obtained from in silico studies, both substrates (controls)—progesterone (
1) and Δ
1-progesterone (
2)—as well as the obtained products (
3–
8) can effectively penetrate the intestine/blood barrier in passive mode and should passively penetrate the blood/brain barrier [
65]. Additionally, the model predicts that the tested compounds (
3–
8) may be efficiently removed from these compartments by the P-glycoprotein. We can observe that most of the obtained compounds are predicted to be removed by P-glycoprotein from the bloodstream and the central nervous system (
Supplementary Materials: Figures S1, S8, S15, S22, S29, S36, S38 and S45). P-glycoprotein (P-gp), also known as multidrug resistance protein 1 (MDR1), is an efflux transporter that influences the absorption, distribution, and elimination of a variety of compounds [
66]. The attachment of a hydroxy group impacts the stability and solubility of steroids and increases their hydrophilicity, thus affecting their bioavailability and bioactivity [
67,
68]. However, the dosage form of the steroid can also impact its bioavailability [
69].
Naturally occurring sex steroids, such as estradiol, estrogen sulfates, and progesterone, or their derivatives pose challenges in determining bioavailability. These compounds demonstrate effective absorption but undergo rapid metabolism and elimination. Primarily used in hormone replacement therapy, they necessitate high doses—for instance, 2000 µg of estradiol or at least 625 µg of estrone sulfate, compared to 10 µg of the synthetic estrogen ethinylestradiol. Similarly, 200,000 µg of progesterone is required compared to 150–1000 µg of most synthetic gestogens. The need for such elevated doses serves as an indicative factor of the low bioavailability within this category of sex steroids [
70]. In addition, the described products (
3–
8) should not be inhibitors of monooxygenases necessary for the proper functioning of the human body. Based on the predicted pharmacokinetic and pharmacodynamic data (
Table 3), 11α-hydroxypregn-1,4-diene-3,20-dione (
3), 6β,11α-dihydroxypregn-1,4-diene-3,20-dione (
4), 6β-hydroxypregn-1,4-diene-3,11,20-trione (
5), 6β,17α-dihydroxypregn-1,4-diene-3,20-dione (
6), 6β,17β-dihydroxyandrost-1,4-diene-3-one (
7), and 12β,17α-dihydroxypregn-1,4-diene-3,20-dione (
8) can be considered as potential drugs [
64].
Based on our in silico studies using the platform Way2Drug PASS Online (
http://www.way2drug.com/PASSOnline/predict.php, accessed on 30 May 2023), information was obtained that the described substrates (
1,
2) and products (
3–
8) should be, with very high probability, the substrates for many cytochrome P-450 monooxygenases (
Table 4), which should result in their rapid metabolism within the human body. Additionally, all described compounds should be, with high probability, the inducers of CYP3A monooxygenases. The CYP3A subfamily of cytochrome P450 (CYP) in humans predominantly consists of CYP3A4, 3A5, and 3A7 (
Supplementary Materials: Figures S2, S9, S16, S23, S30, S37, S39 and S46). CYP3A forms play a crucial role in the metabolism of a surprisingly large number of structurally diverse xenobiotics and endobiotics [
71,
72]. The pregnane X receptor (PXR), efficiently activated by natural C21 steroids (pregnanes), has been implicated in regulating
CYP3A genes in response to xenobiotics [
73]. PXR can bind to and activate transcription from specific response elements present in the
CYP3A gene promoter across multiple species [
74,
75].
Progesterone (
1), based on the predictions made (
Table 5), should be an effective inhibitor of CYP17. However, the resulting compounds (
2–
8) should be slightly weaker inhibitors of this enzyme than progesterone (
1). The inhibition of the key enzyme catalyzing the biosynthesis of androgens from the pregnane precursors, specifically the 17-hydroxy/17,20-lyase (referred to as CYP17), could effectively prevent androgen production from all sources. Consequently, the complete suppression of androgen production through the potent CYP17 inhibitors may offer an effective treatment strategy for prostate cancer patients [
76,
77]. However, it is worth noting that an increased risk of advanced breast cancer is associated with a common allele of the cytochrome P450cl7α gene (CYP17), denoted as A2. The CYP17 genotype demonstrates an association with serum hormone levels among 83 young, nulliparous women. Women heterozygous and homozygous for the CYP17 A2 allele exhibit 7% and 28% higher serum estradiol levels, respectively, and 24% and 30% higher progesterone levels around day 22 of the menstrual cycle compared to women with the A1/A1 genotype [
78]. It is important to acknowledge that CYP17A1 serves as an essential human steroidogenic enzyme, facilitating two sequential reactions leading to the formation of androstenedione from progesterone and dehydroepiandrosterone from pregnenolone. The second reaction involves C17–C20 bond scission, which is significantly influenced by the presence of cytochrome b5. Moreover, this reaction displays a more pronounced acceleration, particularly when 17α-hydroxyprogesterone is a substrate [
79].
The described compounds (
2–
8) should very likely be the substrates for UDP-glucuronosyltransferases (UGTs) (
Table 5). The UGT1A enzymes are involved in the metabolism and detoxification of numerous therapeutic drugs and other xenobiotics [
80,
81]. The UGT1A and UGT2B subfamilies of enzymes are responsible for glucuronidation, which is a major Phase II metabolic pathway that conjugates numerous drugs [
82]. They should also, with a high probability, be the substrates for sulfotransferase (
Table 6). Sulfotransferases (SULTs) are enzymes that catalyze the transfer of a sulfonate group from the universal sulfate donor to an acceptor group of numerous substrates, which facilitates sulfated compound transport into cells. This process is important for the metabolism and regulation of steroids in various physiological processes [
83,
84].
Based on our prediction studies using the Way2Drug PASS Online platform (
http://www.way2drug.com/PASSOnline/predict.php, accessed on 30 May 2023), information was obtained that the described substrates (
1,
2) and products (
3–
8) with very high probability should be inhibitors of testosterone 17β-dehydrogenase (NADP
+), known also as 17β-HSD3: an enzyme playing an important role in the regulation of steroid hormones, such as estrogens and androgens, by catalyzing the reduction of 17-ketosteroids or the oxidation of 17β-hydroxysteroids using NAD(P)H or NAD(P)
+ as a cofactor [
85]. Most of the obtained products should also, with a high probability, find potential uses as androgen antagonists, and prostate as well as menopausal disorders treatments. They should also demonstrate immunosuppressive, erythropoiesis-stimulating, and anti-inflammatory properties (
Table 6).








{kind=link}
{kind=link}
{kind=link}
{kind=link}