
Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic



Abstract
:1. Introduction
2. Results
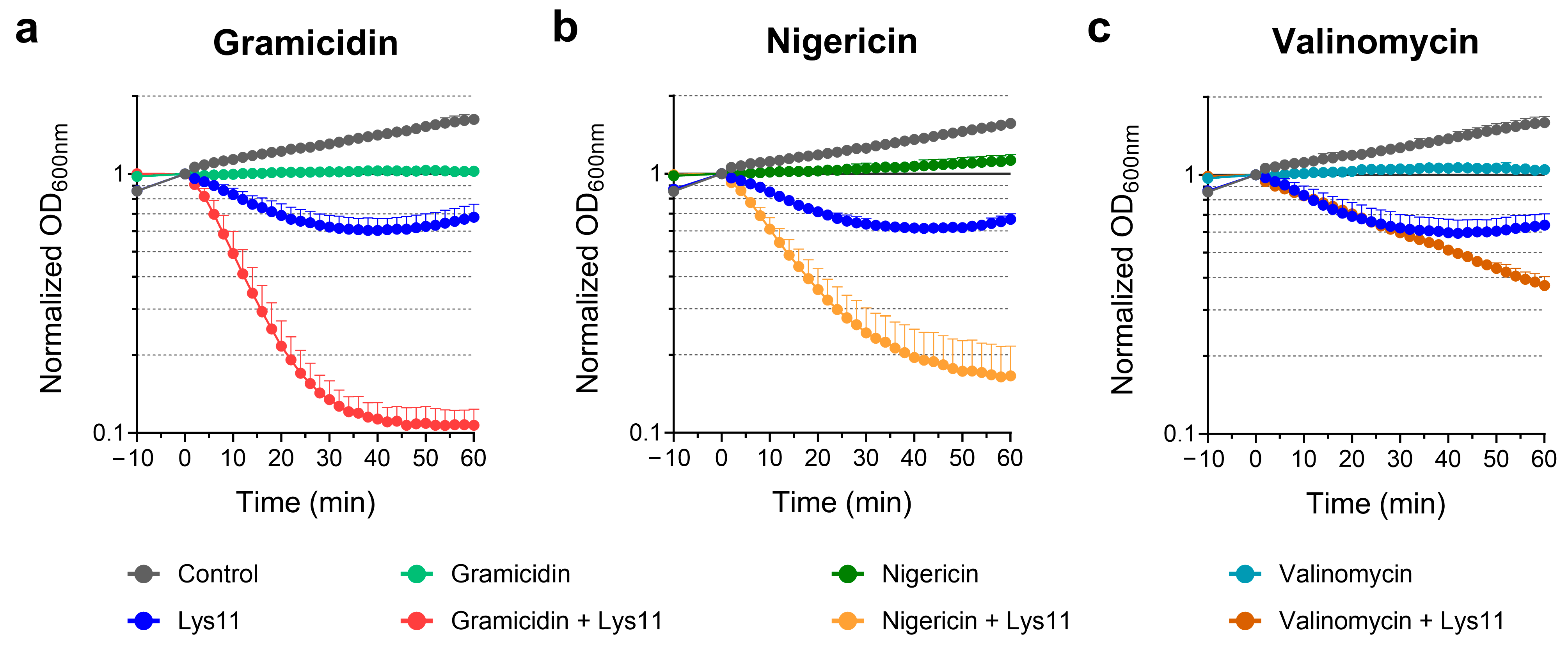
2.1. The ΔpH Component of the PMF Has Major Contribution to Endolysin Tolerance
2.2. Dissipation of Both PMF Components Stimulates Endolysin Binding to Cells
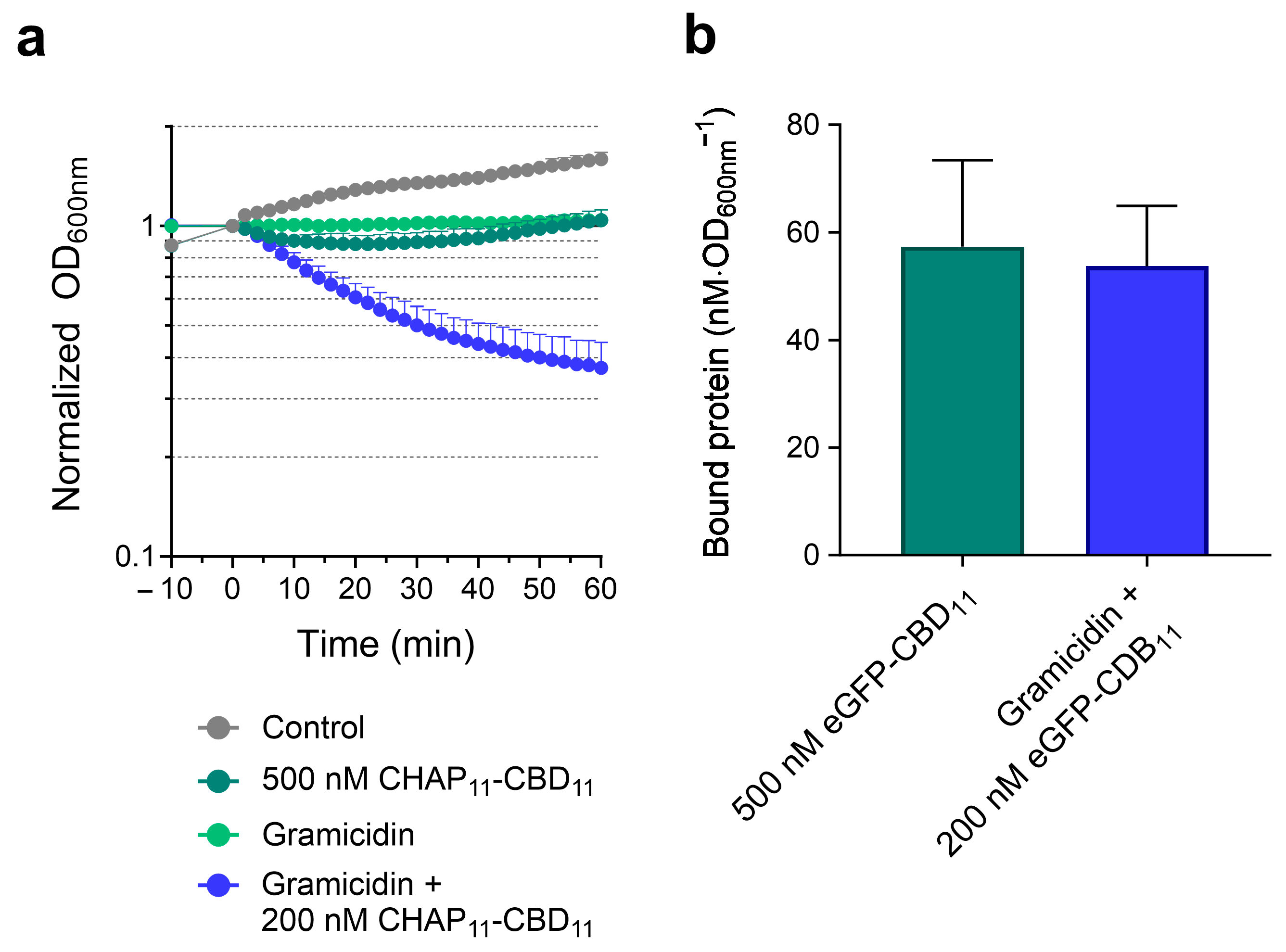
2.3. PMF Dissipation Simultaneously Favors Endolysin Binding and Peptidoglycan Cleavage
2.4. WTA Is the Key CW Glycopolymer Contributing to Endolysin Tolerance
2.5. WTA Drastically Hinders Binding Meditated by CBD11
2.6. Lys11 Catalytic Domains Respond Differently to Cell Tolerance Determinants
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and General Growth Conditions
4.2. Generation of Endolysin Variants
4.3. Protein Production and Purification
4.4. Bacteriolysis Assays
4.5. Binding of eGFP-Endolysin Fusions to Cells
4.6. Bioinformatics Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- CDC—US Center for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 1 November 2023).
- IACG—Interagency Coordination Group on Antimicrobial Research. No Time to Wait: Securing the Future from Drug-Resistant Infections. Report to the Secretary-General of the United Nations. 2019. Available online: https://www.who.int/docs/default-source/documents/no-time-to-wait-securing-the-future-from-drug-resistant-infections-en.pdf (accessed on 1 November 2023).
- World Bank. Drug-Resistant Infections: A Threat to Our Economic Future; World Bank: Washington, DC, USA, 2017; Available online: https://documents1.worldbank.org/curated/en/323311493396993758/pdf/final-report.pdf (accessed on 1 November 2023).
- Theuretzbacher, U.; Gottwalt, S.; Beyer, P.; Butler, M.; Czaplewski, L.; Lienhardt, C.; Moja, L.; Paul, M.; Paulin, S.; Rex, J.H.; et al. Analysis of the clinical antibacterial and antituberculosis pipeline. Lancet Infect. Dis. 2019, 19, e40–e50. [Google Scholar] [CrossRef] [PubMed]
- De Maesschalck, V.; Gutiérrez, D.; Paeshuyse, J.; Lavigne, R.; Briers, Y. Advanced engineering of third-generation lysins and formulation strategies for clinical applications. Crit. Rev. Microbiol. 2020, 46, 548–564. [Google Scholar] [CrossRef]
- Dams, D.; Briers, Y. Enzybiotics: Enzyme-Based Antibacterials as Therapeutics. Adv. Exp. Med. Biol. 2019, 1148, 233–253. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Loessner, M.J. Bacteriophage endolysins—Extending their application to tissues and the bloodstream. Curr. Opin. Biotechnol. 2021, 68, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, K.; Gerstmans, H.; Saafan, A.; Dishisha, T.; Briers, Y. The Preclinical and Clinical Progress of Bacteriophages and Their Lytic Enzymes: The Parts are Easier than the Whole. Viruses 2019, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.; São-José, C. Enzymes and Mechanisms Employed by Tailed Bacteriophages to Breach the Bacterial Cell Barriers. Viruses 2018, 10, 396. [Google Scholar] [CrossRef]
- Cahill, J.; Young, R. Phage Lysis: Multiple Genes for Multiple Barriers. Adv. Virus Res. 2019, 103, 33–70. [Google Scholar] [CrossRef]
- Harold, F.M. Conservation and transformation of energy by bacterial membranes. Bacteriol. Rev. 1972, 36, 172–230. [Google Scholar] [CrossRef]
- Bakker, E.P.; Mangerich, W.E. Interconversion of components of the bacterial proton motive force by electrogenic potassium transport. J. Bacteriol. 1981, 147, 820–826. [Google Scholar] [CrossRef]
- Krulwich, T.A.; Sachs, G.; Padan, E. Molecular aspects of bacterial pH sensing and homeostasis. Nat. Rev. Microbiol. 2011, 9, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Weidenmaier, C.; Peschel, A. Teichoic acids and related cell-wall glycopolymers in Gram-positive physiology and host interactions. Nat. Rev. Microbiol. 2008, 6, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.; Santa Maria, J.P.; Walker, S. Wall teichoic acids of Gram-positive bacteria. Annu. Rev. Microbiol. 2013, 67, 313–336. [Google Scholar] [CrossRef] [PubMed]
- Percy, M.G.; Gründling, A. Lipoteichoic acid synthesis and function in Gram-positive bacteria. Annu. Rev. Microbiol. 2014, 68, 81–100. [Google Scholar] [CrossRef]
- Catalão, M.J.; Pimentel, M. Mycobacteriophage Lysis Enzymes: Targeting the Mycobacterial Cell Envelope. Viruses 2018, 10, 428. [Google Scholar] [CrossRef]
- Nascimento, J.G.; Guerreiro-Pereira, M.C.; Costa, S.F.; São-José, C.; Santos, M.A. Nisin-triggered activity of Lys44, the secreted endolysin from Oenococcus oeni phage fOg44. J. Bacteriol. 2008, 190, 457–461. [Google Scholar] [CrossRef]
- Proença, D.; Leandro, C.; Garcia, M.; Pimentel, M.; São-José, C. EC300: A phage-based, bacteriolysin-like protein with enhanced antibacterial activity against Enterococcus faecalis. Appl. Microbiol. Biotechnol. 2015, 99, 5137–5149. [Google Scholar] [CrossRef]
- Fernandes, S.; São-José, C. More than a hole: The holin lethal function may be required to fully sensitize bacteria to the lytic action of canonical endolysins. Mol. Microbiol. 2016, 102, 92–106. [Google Scholar] [CrossRef]
- García, P.; Martínez, B.; Rodríguez, L.; Rodríguez, A. Synergy between the phage endolysin LysH5 and nisin to kill Staphylococcus aureus in pasteurized milk. Int. J. Food Microbiol. 2010, 141, 151–155. [Google Scholar] [CrossRef]
- Ibarra-Sánchez, L.A.; Van Tassell, M.L.; Miller, M.J. Antimicrobial behavior of phage endolysin PlyP100 and its synergy with nisin to control Listeria monocytogenes in Queso Fresco. Food Microbiol. 2018, 72, 128–134. [Google Scholar] [CrossRef]
- Gouveia, A.; Pinto, D.; Veiga, H.; Antunes, W.; Pinho, M.G.; São-José, C. Synthetic antimicrobial peptides as enhancers of the bacteriolytic action of staphylococcal phage endolysins. Sci. Rep. 2022, 12, 1245. [Google Scholar] [CrossRef] [PubMed]
- Eugster, M.R.; Loessner, M.J. Wall teichoic acids restrict access of bacteriophage endolysin Ply118, Ply511, and PlyP40 cell wall binding domains to the Listeria monocytogenes peptidoglycan. J. Bacteriol. 2012, 194, 6498–6506. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Paskaleva, E.E.; Mehta, K.K.; Dordick, J.S.; Kane, R.S. Wall Teichoic Acids Are Involved in the Medium-Induced Loss of Function of the Autolysin CD11 against Clostridium difficile. Sci. Rep. 2016, 6, 35616. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zha, J.; Koffas, M.A.G.; Dordick, J.S. Reducing Staphylococcus aureus resistance to lysostaphin using CRISPR-dCas9. Biotechnol. Bioeng. 2019, 116, 3149–3159. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Ton-That, H.; Faull, K.F.; Schneewind, O. Multiple enzymatic activities of the murein hydrolase from staphylococcal phage phi11. Identification of a D-alanyl-glycine endopeptidase activity. J. Biol. Chem. 1999, 274, 15847–15856. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, L.K.; Doyle, R.J.; Streips, U.N. The energized membrane and cellular autolysis in Bacillus subtilis. Cell 1981, 25, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Patton, T.G.; Yang, S.J.; Bayles, K.W. The role of proton motive force in expression of the Staphylococcus aureus cid and lrg operons. Mol. Microbiol. 2006, 59, 1395–1404. [Google Scholar] [CrossRef]
- Son, B.; Kong, M.; Ryu, S. The Auxiliary Role of the Amidase Domain in Cell Wall Binding and Exolytic Activity of Staphylococcal Phage Endolysins. Viruses 2018, 10, 284. [Google Scholar] [CrossRef]
- Corrigan, R.M.; Abbott, J.C.; Burhenne, H.; Kaever, V.; Gründling, A. c-di-AMP is a new second messenger in Staphylococcus aureus with a role in controlling cell size and envelope stress. PLoS Pathog. 2011, 7, e1002217. [Google Scholar] [CrossRef]
- Brown, S.; Xia, G.; Luhachack, L.G.; Campbell, J.; Meredith, T.C.; Chen, C.; Winstel, V.; Gekeler, C.; Irazoqui, J.E.; Peschel, A.; et al. Methicillin resistance in Staphylococcus aureus requires glycosylated wall teichoic acids. Proc. Natl. Acad. Sci. USA 2012, 109, 18909–18914. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M.; Jack, R.W.; Kalbacher, H.; Jung, G.; Götz, F. Inactivation of the dlt operon in Staphylococcus aureus confers sensitivity to defensins, protegrins, and other antimicrobial peptides. J. Biol. Chem. 1999, 274, 8405–8410. [Google Scholar] [CrossRef] [PubMed]
- Wanner, S.; Schade, J.; Keinhörster, D.; Weller, N.; George, S.E.; Kull, L.; Bauer, J.; Grau, T.; Winstel, V.; Stoy, H.; et al. Wall teichoic acids mediate increased virulence in Staphylococcus aureus. Nat. Microbiol. 2017, 2, 16257. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Feng, Y.; Feng, X.; Sun, C.; Lei, L.; Ding, W.; Niu, F.; Jiao, L.; Yang, M.; Li, Y.; et al. Structural and biochemical characterization reveals LysGH15 as an unprecedented “EF-hand-like” calcium-binding phage lysin. PLoS Pathog. 2014, 10, e1004109. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; São-José, C.; Azeredo, J. Phage-Derived Peptidoglycan Degrading Enzymes: Challenges and Future Prospects for In Vivo Therapy. Viruses 2018, 10, 292. [Google Scholar] [CrossRef]
- Koch, A.L. The pH in the neighborhood of membranes generating a protonmotive force. J. Theor. Biol. 1986, 120, 73–84. [Google Scholar] [CrossRef]
- Kemper, M.A.; Urrutia, M.M.; Beveridge, T.J.; Koch, A.L.; Doyle, R.J. Proton motive force may regulate cell wall-associated enzymes of Bacillus subtilis. J. Bacteriol. 1993, 175, 5690–5696. [Google Scholar] [CrossRef]
- Calamita, H.G.; Ehringer, W.D.; Koch, A.L.; Doyle, R.J. Evidence that the cell wall of Bacillus subtilis is protonated during respiration. Proc. Natl. Acad. Sci. USA 2001, 98, 15260–15263. [Google Scholar] [CrossRef]
- Calamita, H.G.; Doyle, R.J. Regulation of autolysins in teichuronic acid-containing Bacillus subtilis cells. Mol. Microbiol. 2002, 44, 601–606. [Google Scholar] [CrossRef]
- Biswas, R.; Martinez, R.E.; Göhring, N.; Schlag, M.; Josten, M.; Xia, G.; Hegler, F.; Gekeler, C.; Gleske, A.K.; Götz, F.; et al. Proton-binding capacity of Staphylococcus aureus wall teichoic acid and its role in controlling autolysin activity. PLoS ONE 2012, 7, e41415. [Google Scholar] [CrossRef]
- López, R.; García, E. Recent trends on the molecular biology of pneumococcal capsules, lytic enzymes, and bacteriophage. FEMS Microbiol. Rev. 2004, 28, 553–580. [Google Scholar] [CrossRef]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [PubMed]
- São-José, C. Engineering of Phage-Derived Lytic Enzymes: Improving Their Potential as Antimicrobials. Antibiotics 2018, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Sass, P.; Bierbaum, G. Lytic activity of recombinant bacteriophage phi11 and phi12 endolysins on whole cells and biofilms of Staphylococcus aureus. Appl. Environ. Microbiol. 2007, 73, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Ruas-Madiedo, P.; Fernández, L.; Campelo, A.B.; Briers, Y.; Nielsen, M.W.; Pedersen, K.; Lavigne, R.; García, P.; et al. Design and Selection of Engineered Lytic Proteins with Staphylococcus aureus decolonizing activity. Front. Microbiol. 2021, 12, 723834. [Google Scholar] [CrossRef]
- Wysocka, A.; Łężniak, Ł.; Jagielska, E.; Sabała, I. Electrostatic Interaction with the Bacterial Cell Envelope Tunes the Lytic Activity of Two Novel Peptidoglycan Hydrolases. Microbiol. Spectr. 2022, 10, e0045522. [Google Scholar] [CrossRef]
- Shang, X.; Nelson, D.C. Contributions of Net Charge on the PlyC Endolysin CHAP Domain. Antibiotics 2019, 8, 70. [Google Scholar] [CrossRef]
- Zhao, H.; Eszterhas, S.; Furlon, J.; Cheng, H.; Griswold, K.E. Electrostatic-Mediated Affinity Tuning of Lysostaphin Accelerates Bacterial Lysis Kinetics and Enhances In Vivo Efficacy. Antimicrob. Agents Chemother. 2021, 65, e02199-20. [Google Scholar] [CrossRef]
- Fischer, W.; Rösel, P.; Koch, H.U. Effect of alanine ester substitution and other structural features of lipoteichoic acids on their inhibitory activity against autolysins of Staphylococcus aureus. J. Bacteriol. 1981, 146, 467–475. [Google Scholar] [CrossRef]
- Bierbaum, G.; Sahl, H.G. Autolytic system of Staphylococcus simulans 22: Influence of cationic peptides on activity of N-acetylmuramoyl-L-alanine amidase. J. Bacteriol. 1987, 169, 5452–5458. [Google Scholar] [CrossRef]
- Fedtke, I.; Mader, D.; Kohler, T.; Moll, H.; Nicholson, G.; Biswas, R.; Henseler, K.; Götz, F.; Zähringer, U.; Peschel, A. A Staphylococcus aureus ypfP mutant with strongly reduced lipoteichoic acid (LTA) content: LTA governs bacterial surface properties and autolysin activity. Mol. Microbiol. 2007, 65, 1078–1091. [Google Scholar] [CrossRef]
- Reichmann, N.T.; Piçarra Cassona, C.; Monteiro, J.M.; Bottomley, A.L.; Corrigan, R.M.; Foster, S.J.; Pinho, M.G.; Gründling, A. Differential localization of LTA synthesis proteins and their interaction with the cell division machinery in Staphylococcus aureus. Mol. Microbiol. 2014, 92, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Schlag, M.; Biswas, R.; Krismer, B.; Kohler, T.; Zoll, S.; Yu, W.; Schwarz, H.; Peschel, A.; Götz, F. Role of staphylococcal wall teichoic acid in targeting the major autolysin Atl. Mol. Microbiol. 2010, 75, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Flores-Kim, J.; Dobihal, G.S.; Fenton, A.; Rudner, D.Z.; Bernhardt, T.G. A switch in surface polymer biogenesis triggers growth-phase-dependent and antibiotic-induced bacteriolysis. Elife 2019, 8, e44912. [Google Scholar] [CrossRef] [PubMed]
- Rice, K.C.; Bayles, K.W. Molecular control of bacterial death and lysis. Microbiol. Mol. Biol. Rev. 2008, 72, 85–109. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, F.C.; Baddiley, J. A continuum of anionic charge: Structures and functions of D-alanyl-teichoic acids in Gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [PubMed]
- Bernal, P.; Zloh, M.; Taylor, P.W. Disruption of D-alanyl esterification of Staphylococcus aureus cell wall teichoic acid by the β-lactam resistance modifier (-)-epicatechin gallate. J. Antimicrob. Chemother. 2009, 63, 1156–1162. [Google Scholar] [CrossRef] [PubMed]
- Saar-Dover, R.; Bitler, A.; Nezer, R.; Shmuel-Galia, L.; Firon, A.; Shimoni, E.; Trieu-Cuot, P.; Shai, Y. D-alanylation of lipoteichoic acids confers resistance to cationic peptides in group B Streptococcus by increasing the cell wall density. PLoS Pathog. 2012, 8, e1002891. [Google Scholar] [CrossRef]
- São-José, C.; Parreira, R.; Vieira, G.; Santos, M.A. The N-terminal region of the Oenococcus oeni bacteriophage fOg44 lysin behaves as a bona fide signal peptide in Escherichia coli and as a cis-inhibitory element, preventing lytic activity on oenococcal cells. J. Bacteriol. 2000, 182, 5823–5831. [Google Scholar] [CrossRef]
- Kreiswirth, B.N.; Löfdahl, S.; Betley, M.J.; O’Reilly, M.; Schlievert, P.M.; Bergdoll, M.S.; Novick, R.P. The toxic shock syndrome exotoxin structural gene is not detectably transmitted by a prophage. Nature 1983, 305, 709–712. [Google Scholar] [CrossRef]
- Gründling, A.; Schneewind, O. Genes required for glycolipid synthesis and lipoteichoic acid anchoring in Staphylococcus aureus. J. Bacteriol. 2007, 189, 2521–2530. [Google Scholar] [CrossRef]
- Iordanescu, S.; Surdeanu, M. Two restriction and modification systems in Staphylococcus aureus NCTC8325. J. Gen. Microbiol. 1976, 96, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Lamsa, A.; Liu, W.T.; Dorrestein, P.C.; Pogliano, K. The Bacillus subtilis cannibalism toxin SDP collapses the proton motive force and induces autolysis. Mol. Microbiol. 2012, 84, 486–500. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, A.; Pinto, D.; Vítor, J.; São-José, C. Research Data of Gouveia et al.—Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic; FigShare: Iasi, Romania, 2023; Available online: https://figshare.com/articles/dataset/Research_data_of_Gouveia_et_al_-_Cellular_and_enzymatic_determinants_impacting_the_exolytic_action_of_an_anti-staphylococcal_enzybiotic/24747519 (accessed on 8 December 2023).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Features | Reference/Source |
|---|---|---|
| E. coli | ||
| XL1-Blue MRF’ | Cloning strain for recovering of plasmid constructs | Stratagene |
| CG61 | Protein expression strain; produces phage T7 RNA polymerase upon thermal induction. Used to produce all Lys11 variants. Selection with 40 μg/mL kanamycin | [61] |
| S. aureus | ||
| RN4220 | Prophage-cured, restriction-deficient mutant of strain 8325-4 | [62] |
| RN4220ΔtarMΔtarS | Derivative of RN4220 lacking α- and β-O-GlcNAcylation due to tarM and tarS deletion | [33] |
| RN4220Δspa | In-frame deletion in spa coding for protein A | [63] |
| RN4220ΔspaΔltaS (supressor strain 4S5) | Derivative of RN4220Δspa lacking LTA as result of ltaS deletion. Carries a mutation suppressing the ΔltaS lethal phenotype | [32] |
| SA113 | Mutant strain of 8325, with an agr- background and 11-bp deletion in rsbU | [64] |
| SA113ΔdltA | Derivative of SA113 lacking D-alanylation of TA due to dltA deletion. Selection with 120 µg/mL spectinomycin | [34] |
| Lys11 Variant | Features | Reference/Source |
|---|---|---|
| Lys11 | 3-domain endolysin: CHAP11, Ami11 and CBD11 | [21] |
| CHAP11-CBD11 | Lys11 lacking Ami11. Deletion encompassing residues 151 to 360 of Lys11 | This work |
| Ami11-CBD11 | Lys11 lacking CHAP11. Deletion encompassing residues 2 to 178 of Lys11 | This work |
| eGFP-Ami11-CBD11 | Ami11-CBD11 (P149 to S481 of Lys11) fused to the C-terminus of eGFP | [24] |
| eGFP-Ami11 | Ami11 (P149 to M360 of Lys11) fused to the C-terminus of eGFP | This work |
| eGFP-CBD11 | CBD11 (D361 to S481 of Lys11) fused to the C-terminus of eGFP | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouveia, A.; Pinto, D.; Vítor, J.M.B.; São-José, C. Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic. Int. J. Mol. Sci. 2024, 25, 523. https://doi.org/10.3390/ijms25010523
Gouveia A, Pinto D, Vítor JMB, São-José C. Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic. International Journal of Molecular Sciences. 2024; 25(1):523. https://doi.org/10.3390/ijms25010523
Chicago/Turabian StyleGouveia, Ana, Daniela Pinto, Jorge M. B. Vítor, and Carlos São-José. 2024. "Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic" International Journal of Molecular Sciences 25, no. 1: 523. https://doi.org/10.3390/ijms25010523
APA StyleGouveia, A., Pinto, D., Vítor, J. M. B., & São-José, C. (2024). Cellular and Enzymatic Determinants Impacting the Exolytic Action of an Anti-Staphylococcal Enzybiotic. International Journal of Molecular Sciences, 25(1), 523. https://doi.org/10.3390/ijms25010523