
Kisspeptin-10 Improves Testicular Redox Status but Does Not Alter the Unfolded Protein Response (UPR) That Is Downregulated by Hypothyroidism in a Rat Model
 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Confirmation of Hypothyroidism
2.2. Immunolocalization of 8-OhdG and Quantification of LOOH, TBARS, ROS and Peroxynitrite
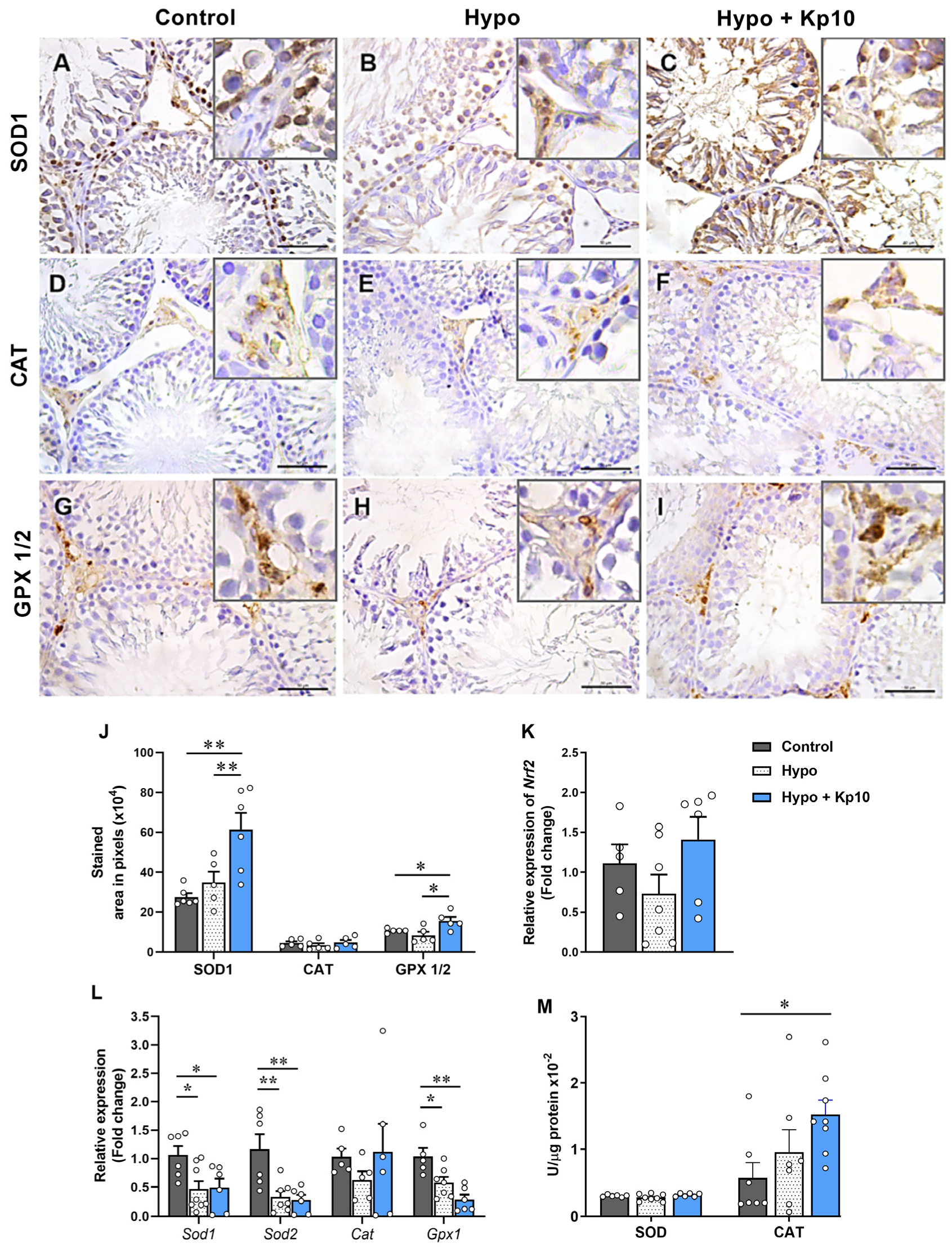
2.3. Treatment with Kp10 Increases Protein Expression of SOD1 and GPX1/2 and Catalase Enzyme Activity in the Testes of Hypothyroid Rats
2.4. Kp10 Treatment Does Not Alter Lower Testicular Gene Expression of UPR Mediators Caused by Hypothyroidism in Rats
2.5. Kp10 Treatment Reduces the Apoptotic Index in Testicular Cells
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Immunohistochemistry (IHC)
4.3. RNA Extraction and Real-Time Quantitative PCR (qPCR)
4.4. Enzymatic Activity of Superoxide Dismutase (SOD) and Catalase (CAT)
4.5. Lipid Peroxidation
4.6. Reactive Oxygen Species (ROS) and Peroxynitrite
4.7. TUNEL Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leslie, S.W.; Soon-Sutton, T.L.; Khan, M.A. Male Infertility. Available online: https://www.statpearls.com/articlelibrary/viewarticle/23503/ (accessed on 13 March 2023).
- Sengupta, P.; Dutta, S.; Karkada, I.R.; Chinni, S.V. Endocrinopathies and Male Infertility. Life 2022, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Alahmar, A.; Dutta, S.; Sengupta, P. Thyroid Hormones in Male Reproduction and Infertility. Asian Pac. J. Reprod. 2019, 8, 203–210. [Google Scholar] [CrossRef]
- Wagner, M.S.; Wajner, S.M.; Maia, A.L. The Role of Thyroid Hormone in Testicular Development and Function. J. Endocrinol. 2008, 199, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.C.; Martinha, J.; Santana, S.; Barbosa, E.M.; Santos, B.R.; Mendonça, L.D.; Clara, M.; Cunha, G.; Machado, W.M.; Santana, L.R.; et al. Kisspeptin Treatment Reverses High Prolactin Levels and Improves Gonadal Function in Hypothyroid Male Rats. Sci. Rep. 2023, 13, 16819. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.A.; Mohammed, N.A.; Eid, K.A.; Abomughaid, M.M.; Abdelazim, A.M.; Aboregela, A.M. Hypothyroidism: Morphological and Metabolic Changes in the Testis of Adult Albino Rat and the Amelioration by Alpha-Lipoic Acid. Folia Morphol. 2021, 80, 352–362. [Google Scholar] [CrossRef] [PubMed]
- François Xavier, K.N.; Patrick Brice, D.D.; Modeste, W.N.; Esther, N.; Albert, K.; Pierre, K.; Pierre, W. Preventive Effects of Aframomum Melegueta Extracts on the Reproductive Complications of Propylthiouracil-Induced Hypothyroidism in Male Rat. Andrologia 2019, 51, e13306. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Roy, A. Compromised Rat Testicular Antioxidant Defence System by Hypothyroidism before Puberty. Int. J. Endocrinol. 2012, 2012, 637825. [Google Scholar] [CrossRef]
- Mann, T. Secretory Function of the Prostate, Seminal Vesicle and Other Male Accessory Organs of Reproduction. J. Reprod. Fertil. 1974, 37, 179–188. [Google Scholar] [CrossRef]
- Higgins, S.J.; Burchell, J.M. Effects of Testosterone on Messenger Ribonucleic Acid and Protein Synthesis in Rat Seminal Vesicle. Biochem. J. 1978, 174, 543–551. [Google Scholar] [CrossRef]
- Alipour, F.; Jalali, M.; Nikravesh, M.R.; Fazel, A.; Sankian, M.; Khordad, E. Assessment of Sperm Morphology, Chromatin Integrity, and CatSper Genes Expression in Hypothyroid Mice. Acta Biol. Hung. 2018, 69, 244–258. [Google Scholar] [CrossRef]
- Romano, R.M.; Gomes, S.N.; Cardoso, N.C.S.; Schiessl, L.; Romano, M.A.; Oliveira, C.A. New Insights for Male Infertility Revealed by Alterations in Spermatic Function and Differential Testicular Expression of Thyroid-Related Genes. Endocrine 2017, 55, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Bolisetty, S.; Jaimes, E. Mitochondria and Reactive Oxygen Species: Physiology and Pathophysiology. Int. J. Mol. Sci. 2013, 14, 6306–6344. [Google Scholar] [CrossRef] [PubMed]
- Bayr, H. Reactive Oxygen Species. Crit. Care Med. 2005, 33, S498–S501. [Google Scholar] [CrossRef]
- Burton, G.J.; Yung, H.W. Endoplasmic Reticulum Stress in the Pathogenesis of Early-Onset Pre-Eclampsia. Pregnancy Hypertens 2011, 1, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Kaufman, R.J. From Endoplasmic-Reticulum Stress to the Inflammatory Response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Almanza, A.; Carlesso, A.; Chintha, C.; Creedican, S.; Doultsinos, D.; Leuzzi, B.; Luís, A.; McCarthy, N.; Montibeller, L.; More, S.; et al. Endoplasmic Reticulum Stress Signalling—From Basic Mechanisms to Clinical Applications. FEBS J. 2019, 286, 241–278. [Google Scholar] [CrossRef]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of Endoplasmic Reticulum Stress-Induced Apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A Cell’s Response to Stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [CrossRef]
- Veerbeek, J.H.W.; Tissot Van Patot, M.C.; Burton, G.J.; Yung, H.W. Endoplasmic Reticulum Stress Is Induced in the Human Placenta during Labour. Placenta 2015, 36, 88–92. [Google Scholar] [CrossRef]
- Torres-Manzo, A.P.; Franco-Colín, M.; Blas-Valdivia, V.; Pineda-Reynoso, M.; Cano-Europa, E. Hypothyroidism Causes Endoplasmic Reticulum Stress in Adult Rat Hippocampus: A Mechanism Associated with Hippocampal Damage. Oxid. Med. Cell. Longev. 2018, 2018, 2089404. [Google Scholar] [CrossRef]
- dos Anjos Cordeiro, J.M.; Santos, L.C.; de Oliveira, L.S.; Santos, B.R.; Santos, E.O.; Barbosa, E.M.; de Macêdo, I.O.; de Freitas, G.J.C.; de Assis Santos, D.; de Lavor, M.S.L.; et al. Maternal Hypothyroidism Causes Oxidative Stress and Endoplasmic Reticulum Stress in the Maternal-Fetal Interface of Rats. Free Radic. Biol. Med. 2022, 191, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Shaygannia, E.; Rahmani, M.; Eskandari, A.; Golsefid, A.A.; Tavalaee, M.; Gharagozloo, P.; Drevet, J.R.; Nasr-Esfahani, M.H. Endoplasmic Reticulum Stress (ER Stress) and Unfolded Protein Response (UPR) Occur in a Rat Varicocele Testis Model. Oxid. Med. Cell. Longev. 2020, 2020, 5909306. [Google Scholar] [CrossRef]
- Yu, C.; Jiang, F.; Zhang, M.; Luo, D.; Shao, S.; Zhao, J.; Gao, L.; Zuo, C.; Guan, Q. HC Diet Inhibited Testosterone Synthesis by Activating Endoplasmic Reticulum Stress in Testicular Leydig Cells. J. Cell. Mol. Med. 2019, 23, 3140–3150. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.L.; Wang, H.; Zhang, C.; Zhang, Y.; Zhao, M.; Chen, Y.H.; Xu, D.X. N-Acetylcysteine Protects against Cadmium-Induced Germ Cell Apoptosis by Inhibiting Endoplasmic Reticulum Stress in Testes. Asian J. Androl. 2013, 15, 290. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, C.; Liu, H.; Guan, Z.; Zeng, Q.; Zhang, C.; Lei, R.; Xia, T.; Gao, H.; Yang, L.; et al. Fluoride-Elicited Developmental Testicular Toxicity in Rats: Roles of Endoplasmic Reticulum Stress and Inflammatory Response. Toxicol. Appl. Pharmacol. 2013, 271, 206–215. [Google Scholar] [CrossRef]
- Santos, B.R.; dos Anjos Cordeiro, J.M.; Santos, L.C.; Barbosa, E.M.; Mendonça, L.D.; Santos, E.O.; de Macedo, I.O.; de Lavor, M.S.L.; Szawka, R.E.; Serakides, R.; et al. Kisspeptin Treatment Improves Fetal-Placental Development and Blocks Placental Oxidative Damage Caused by Maternal Hypothyroidism in an Experimental Rat Model. Front. Endocrinol. 2022, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Asadi, N.; Bahmani, M.; Kheradmand, A.; Rafieian-Kopaei, M. The Impact of Oxidative Stress on Testicular Function and the Role of Antioxidants in Improving It: A Review. J. Clin. Diagn. Res. 2017, 11, IE01–IE05. [Google Scholar] [CrossRef]
- Gottsch, M.L.; Cunningham, M.J.; Smith, J.T.; Popa, S.M.; Acohido, B.V.; Crowley, W.F.; Seminara, S.; Clifton, D.K.; Steiner, R.A. A Role for Kisspeptins in the Regulation of Gonadotropin Secretion in the Mouse. Endocrinology 2004, 145, 4073–4077. [Google Scholar] [CrossRef]
- Pinilla, L.; Aguilar, E.; Dieguez, C.; Millar, R.P.; Tena-Sempere, M. Kisspeptins and Reproduction: Physiological Roles and Regulatory Mechanisms. Physiol. Rev. 2012, 92, 1235–1316. [Google Scholar] [CrossRef]
- Cao, Y.; Li, Z.; Jiang, W.; Ling, Y.; Kuang, H. Reproductive Functions of Kisspeptin/KISS1R Systems in the Periphery. Reprod. Biol. Endocrinol. 2019, 17, 65. [Google Scholar] [CrossRef]
- Feng, T.; Bai, J.H.; Xu, X.L.; Liu, Y. Kisspeptin and Its Effect on Mammalian Spermatogensis. Curr. Drug Metab. 2019, 20, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Aslan, M.; Erkanli Senturk, G.; Akkaya, H.; Sahin, S.; Yılmaz, B. The Effect of Oxytocin and Kisspeptin-10 in Ovary and Uterus of Ischemia-Reperfusion Injured Rats. Taiwan J. Obstet. Gynecol. 2017, 56, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Akkaya, H.; Eyuboglu, S.; Erkanlı Senturk, G.; Yilmaz, B. Investigation of the Effects of Kisspeptin-10 in Methionine-Induced Lipid Peroxidation in Testicle Tissue of Young Rats. J. Biochem. Mol. Toxicol. 2017, 31, e21881. [Google Scholar] [CrossRef]
- Akkaya, H. Kisspeptin-10 Administration Regulates MTOR and AKT Activities and Oxidative Stress in Mouse Cardiac Tissue. J. Evol. Biochem. Physiol. 2021, 57, 270–280. [Google Scholar] [CrossRef]
- Akkaya, H.; Sümer, E.; Kutlu, S.; Solak, H.; Yilmaz, B. What Is the Protective Effect of Preischemic Kisspeptin-10 Administration against Ischemia/Reperfusion Injury of Striatum on Mice? Turk. J. Med. Sci. 2022, 52, 1532–1542. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, Y.; Sun, L.; Sun, N.; Wang, J.; Ma, H. Kisspeptin Regulates the Proliferation and Apoptosis of Ovary Granulosa Cells in Polycystic Ovary Syndrome by Modulating the PI3K/AKT/ERK Signalling Pathway. BMC Womens Health 2023, 23, 15. [Google Scholar] [CrossRef]
- Francisqueti-Ferron, F.V.; Ferron, A.J.T.; Garcia, J.L.; de Almeida Silva, C.C.V.; Costa, M.R.; Gregolin, C.S.; Moreto, F.; Ferreira, A.L.A.; Minatel, I.O.; Correa, C.R. Basic Concepts on the Role of Nuclear Factor Erythroid-Derived 2-Like 2 (Nrf2) in Age-Related Diseases. Int. J. Mol. Sci. 2019, 20, 3208. [Google Scholar] [CrossRef]
- Carrington, E.M.; Tarlinton, D.M.; Gray, D.H.; Huntington, N.D.; Zhan, Y.; Lew, A.M. The Life and Death of Immune Cell Types: The Role of BCL-2 Anti-Apoptotic Molecules. Immunol. Cell Biol. 2017, 95, 870–877. [Google Scholar] [CrossRef]
- Salehi, F.; Behboudi, H.; Kavoosi, G.; Ardestani, S.K. Oxidative DNA Damage Induced by ROS-Modulating Agents with the Ability to Target DNA: A Comparison of the Biological Characteristics of Citrus Pectin and Apple Pectin. Sci. Rep. 2018, 8, 13902. [Google Scholar] [CrossRef]
- Feng, D.; Huang, H.; Yang, Y.; Yan, T.; Jin, Y.; Cheng, X.; Cui, L. Ameliorative Effects of N-Acetylcysteine on Fluoride-Induced Oxidative Stress and DNA Damage in Male Rats’ Testis. Mutat. Res. Toxicol. Environ. Mutagen. 2015, 792, 35–45. [Google Scholar] [CrossRef]
- El-Kashlan, A.M.; Nooh, M.M.; Hassan, W.A.; Rizk, S.M. Therapeutic Potential of Date Palm Pollen for Testicular Dysfunction Induced by Thyroid Disorders in Male Rats. PLoS ONE 2015, 10, e0139493. [Google Scholar] [CrossRef]
- Wang, J.L.; Zhang, H.J.; Wang, H.L.; Wang, J.W.; Gou, P.H.; Ye, Z.H.; Wang, Y.L. Influence of Hypothyroidism on Oxidative Stress, c-Fos Expression, Cell Cycle, and Apoptosis in Rats Testes. Toxicol. Environ. Chem. 2015, 97, 1394–1407. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, A.; Ahsan, H. Peroxynitrite: Cellular Pathology and Implications in Autoimmunity. J. Immunoass. Immunochem. 2019, 40, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G. Increasing the Endogenous NO Level Causes Catalase Inactivation and Reactivation of Intercellular Apoptosis Signaling Specifically in Tumor Cells. Redox Biol. 2015, 6, 353–371. [Google Scholar] [CrossRef]
- Sivrikaya, A.; Kolayli, S.; Kucuk, M.; Aliyazicioglu, R. In Vitro Effects of Peroxynitrite Treatment on Fish Liver Catalase Activity. J. Enzym. Inhib. Med. Chem. 2009, 24, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Heinzelmanna, S.; Bauer, G. Multiple Protective Functions of Catalase against Intercellular Apoptosis-Inducing ROS Signaling of Human Tumor Cells. Biol. Chem. 2010, 391, 675–693. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.R.; Yao, Y.L.; Wang, D.; Ma, H.T.; Gou, P.H.; Li, C.Y.; Wang, J.L. Influence of Hypothyroidism on Testicular Mitochondrial Oxidative Stress by Activating the P38 Mitogen-Activated Protein Kinase and c-Jun NH 2 -Terminal Kinase Signaling Pathways in Rats. Hum. Exp. Toxicol. 2019, 38, 95–105. [Google Scholar] [CrossRef]
- Bhattarai, K.R.; Chaudhary, M.; Kim, H.-R.; Chae, H.-J. Endoplasmic Reticulum (ER) Stress Response Failure in Diseases. Trends Cell Biol. 2020, 30, 672–675. [Google Scholar] [CrossRef]
- Wu, J.; Kaufman, R.J. From Acute ER Stress to Physiological Roles of the Unfolded Protein Response. Cell Death Differ. 2006, 13, 374–384. [Google Scholar] [CrossRef]
- Zhang, L.H.; Zhang, X. Roles of GRP78 in Physiology and Cancer. J. Cell. Biochem. 2010, 110, 1299–1305. [Google Scholar] [CrossRef]
- Ham, J.; Lim, W.; You, S.; Song, G. Butylated Hydroxyanisole Induces Testicular Dysfunction in Mouse Testis Cells by Dysregulating Calcium Homeostasis and Stimulating Endoplasmic Reticulum Stress. Sci. Total Environ. 2020, 702, 134775. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.; You, S.; Lim, W.; Song, G. Etoxazole Induces Testicular Malfunction in Mice by Dysregulating Mitochondrial Function and Calcium Homeostasis. Environ. Pollut. 2020, 263, 114573. [Google Scholar] [CrossRef] [PubMed]
- Hillary, R.F.; Fitzgerald, U. A Lifetime of Stress: ATF6 in Development and Homeostasis. J. Biomed. Sci. 2018, 25, 48. [Google Scholar] [CrossRef]
- Yu, R.; Chen, X.; Zhu, X.; He, B.; Lu, C.; Liu, Y.; Xu, X.; Wu, X. ATF6 Deficiency Damages the Development of Spermatogenesis in Male Atf6 Knockout Mice. Andrologia 2022, 54, e14350. [Google Scholar] [CrossRef] [PubMed]
- Yachie, A. Heme Oxygenase-1 Deficiency and Oxidative Stress: A Review of 9 Independent Human Cases and Animal Models. Int. J. Mol. Sci. 2021, 22, 1514. [Google Scholar] [CrossRef]
- Campbell, N.K.; Fitzgerald, H.K.; Dunne, A. Regulation of Inflammation by the Antioxidant Haem Oxygenase 1. Nat. Rev. Immunol. 2021, 21, 411–425. [Google Scholar] [CrossRef]
- Poss, K.D.; Tonegawa, S. Heme Oxygenase 1 Is Required for Mammalian Iron Reutilization. Proc. Natl. Acad. Sci. USA 1997, 94, 10919–10924. [Google Scholar] [CrossRef]
- Yao, H.; Peterson, A.L.; Li, J.; Xu, H.; Dennery, P.A. Heme Oxygenase 1 and 2 Differentially Regulate Glucose Metabolism and Adipose Tissue Mitochondrial Respiration: Implications for Metabolic Dysregulation. Int. J. Mol. Sci. 2020, 21, 7123. [Google Scholar] [CrossRef]
- Hegab, I.I.; El-Latif, R.N.A.; El-Horony, H.E. Targeting Heme Oxygenase-1 in Hypothyroidism Induced Reproductive Dysfunction in Adult Male Rats. Med. J. Cairo Univ. 2019, 87, 607–617. [Google Scholar] [CrossRef]
- Yao, Y.; Chang, X.; Wang, D.; Ma, H.; Wang, H.; Zhang, H.; Li, C.; Wang, J. Roles of ERK1/2 and PI3K/AKT Signaling Pathways in Mitochondria-Mediated Apoptosis in Testes of Hypothyroid Rats. Toxicol. Res. 2018, 7, 1214–1224. [Google Scholar] [CrossRef]
- Huang, Y.; Guo, Y.; Huang, L.; Fang, Y.; Li, D.; Liu, R.; Lu, Q.; Ren, R.; Tang, L.; Lian, L.; et al. Kisspeptin-54 Attenuates Oxidative Stress and Neuronal Apoptosis in Early Brain Injury after Subarachnoid Hemorrhage in Rats via GPR54/ARRB2/AKT/GSK3β Signaling Pathway. Free Radic. Biol. Med. 2021, 171, 99–111. [Google Scholar] [CrossRef]
- Ilie, M.; Khambata-Ford, S.; Copie-Bergman, C.; Huang, L.; Juco, J.; Hofman, V.; Hofman, P. Correction: Use of the 22C3 Anti-PD-L1 Antibody to Determine PD-L1 Expression in Multiple Automated Immunohistochemistry Platforms. PLoS ONE 2017, 12, e0186537. [Google Scholar] [CrossRef]
- Silva, J.F.; Ocarino, N.M.; Serakides, R. Maternal Thyroid Dysfunction Affects Placental Profile of Inflammatory Mediators and the Intrauterine Trophoblast Migration Kinetics. Reproduction 2014, 147, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S.; Marklund, G. Involvement of the Superoxide Anion Radical in the Autoxidation of Pyrogallol and a Convenient Assay for Superoxide Dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Oliveira, K.M.; Binda, N.S.; Lavor, M.S.L.; Silva, C.M.O.; Rosado, I.R.; Gabellini, E.L.A.; Da Silva, J.F.; Oliveira, C.M.; Melo, M.M.; Gomez, M.V.; et al. Conotoxin MVIIA Improves Cell Viability and Antioxidant System after Spinal Cord Injury in Rats. PLoS ONE 2018, 13, e0204948. [Google Scholar] [CrossRef]
- Borges, S.C.; Tironi, L.M.T.; da Silva, L.M.; Buttow, N.C. Curcumin Protects Remote Organs against Injury That Is Caused by Intestinal Ischemia and Reperfusion. Acta Sci. Biol. Sci. 2020, 42, e50588. [Google Scholar] [CrossRef]
- Ferreira, G.F.; de Matos Baltazar, L.; Santos, J.R.A.; Monteiro, A.S.; de Oliveira Fraga, L.A.; Resende-Stoianoff, M.A.; Santos, D.A. The Role of Oxidative and Nitrosative Bursts Caused by Azoles and Amphotericin B against the Fungal Pathogen Cryptococcus Gattii. J. Antimicrob. Chemother. 2013, 68, 1801–1811. [Google Scholar] [CrossRef]
- Silva, J.F.; Ocarino, N.M.; Vieira, A.L.S.; Nascimento, E.F.; Serakides, R. Effects of Hypo-and Hyperthyroidism on Proliferation, Angiogenesis, Apoptosis and Expression of COX-2 in the Corpus Luteum of Female Rats. Reprod. Domest. Anim. 2013, 48, 691–698. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | Hypothyroid | Hypothyroid + Kp10 |
|---|---|---|---|
| Initial body mass (g) | 370.7 ± 6.20 | 371.7 ± 6.27 | 371.7 ± 7.68 |
| Final body mass (g) | 423.1 ± 5.87 | 345.0 ± 6.55 **** | 333.5 ± 8.07 **** |
| Free T4 (ng/dL) | 1.699 ± 0.097 | 0.029 ± 0.014 **** | 0.041 ± 0.016 **** |
| Gene | Sequences (5′ → 3′) | Accession No. | |
|---|---|---|---|
| Superoxide dismutase 1 | Sod1 | GAAAGGACGGTGTGGCCAAT | NM_017050.1 |
| CTCGTGGACCACCATAGTACG | |||
| Superoxide dismutase 2 | Sod2 | CGGGGGCCATATCAATCACA | NM_017051.2 |
| GCCTCCAGCAACTCTCCTTT | |||
| Catalase | Cat | CTGACTGACGCGATTGCCTA | NM_012520.2 |
| GTGGTCAGGACATCGGGTTT | |||
| Glutathione peroxidase 1 | Gpx1 | GCGCTACAGCGGATTTTTGA | NM_030826.4 |
| GAAGGCATACACGGTGGACT | |||
| Nuclear factor, erythroid 2-like 2 | Nrf2 | CCCATTGAGGGCTGTGATCT | NM_031789.2 |
| GCCTTCAGTGTGCTTCTGGTT | |||
| Heat shock protein family A (Hsp70) member 5 | Grp78 | TGAAGGGGAGCGTCTGATTG | NM_013083.2 |
| TCATTCCAAGTGCGTCCGAT | |||
| Activating transcription factor 6, | Atf6 | CCAGCAGAAAACCCGCATTC | XM_017598829.1 |
| CAGAATTCCTGATGCTAGTGGTT | |||
| Eukaryotic translation initiation factor 2 alpha kinase 3 | Perk | GGCTGGTGAGGGATGGTAAA | NM_031599.2 |
| TTGGCTGTGTAACTTGTGTCATCA | |||
| X-box binding protein 1 | sXbp1 | CTGAGTCCGCAGCAGGTG | NM_001271731.1 |
| AAGAGGCAACAGCGTCAGAA | |||
| Heme oxygenase 1 | Ho1 | ACAGCACTACGTAAAGCGTCTCCA | NM_012580.2 |
| CATGGCCTTCTGCGCAATCTTCTT | |||
| Apoptosis regulator BCL-2 | Bcl-2 | ACTTCTCTCGTCGCTACCGTC | NM_016993.2 |
| AAGAGTTCCTCCACCACCGT | |||
| Glyceraldehyde-3-Phosphate dehydrogenase | Gapdh | ACAGCCGCATCTTCTTGTGC | NM_017008.4 |
| GCCTCACCCCATTTGATGTT | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, L.C.; dos Anjos Cordeiro, J.M.; Cunha, M.C.d.S.G.; Santos, B.R.; Oliveira, L.S.d.; da Silva, A.L.; Barbosa, E.M.; Niella, R.V.; de Freitas, G.J.C.; Santos, D.d.A.; et al. Kisspeptin-10 Improves Testicular Redox Status but Does Not Alter the Unfolded Protein Response (UPR) That Is Downregulated by Hypothyroidism in a Rat Model. Int. J. Mol. Sci. 2024, 25, 1514. https://doi.org/10.3390/ijms25031514
Santos LC, dos Anjos Cordeiro JM, Cunha MCdSG, Santos BR, Oliveira LSd, da Silva AL, Barbosa EM, Niella RV, de Freitas GJC, Santos DdA, et al. Kisspeptin-10 Improves Testicular Redox Status but Does Not Alter the Unfolded Protein Response (UPR) That Is Downregulated by Hypothyroidism in a Rat Model. International Journal of Molecular Sciences. 2024; 25(3):1514. https://doi.org/10.3390/ijms25031514
Chicago/Turabian StyleSantos, Luciano Cardoso, Jeane Martinha dos Anjos Cordeiro, Maria Clara da Silva Galrão Cunha, Bianca Reis Santos, Luciana Santos de Oliveira, Adriana Lopes da Silva, Erikles Macêdo Barbosa, Raquel Vieira Niella, Gustavo José Cota de Freitas, Daniel de Assis Santos, and et al. 2024. "Kisspeptin-10 Improves Testicular Redox Status but Does Not Alter the Unfolded Protein Response (UPR) That Is Downregulated by Hypothyroidism in a Rat Model" International Journal of Molecular Sciences 25, no. 3: 1514. https://doi.org/10.3390/ijms25031514
APA StyleSantos, L. C., dos Anjos Cordeiro, J. M., Cunha, M. C. d. S. G., Santos, B. R., Oliveira, L. S. d., da Silva, A. L., Barbosa, E. M., Niella, R. V., de Freitas, G. J. C., Santos, D. d. A., Serakides, R., Ocarino, N. d. M., Borges, S. C., de Lavor, M. S. L., & Silva, J. F. (2024). Kisspeptin-10 Improves Testicular Redox Status but Does Not Alter the Unfolded Protein Response (UPR) That Is Downregulated by Hypothyroidism in a Rat Model. International Journal of Molecular Sciences, 25(3), 1514. https://doi.org/10.3390/ijms25031514