

Detailed Characterization of the Lung–Gut Microbiome Axis Reveals the Link between PD-L1 and the Microbiome in Non-Small-Cell Lung Cancer Patients
 , ,
, , 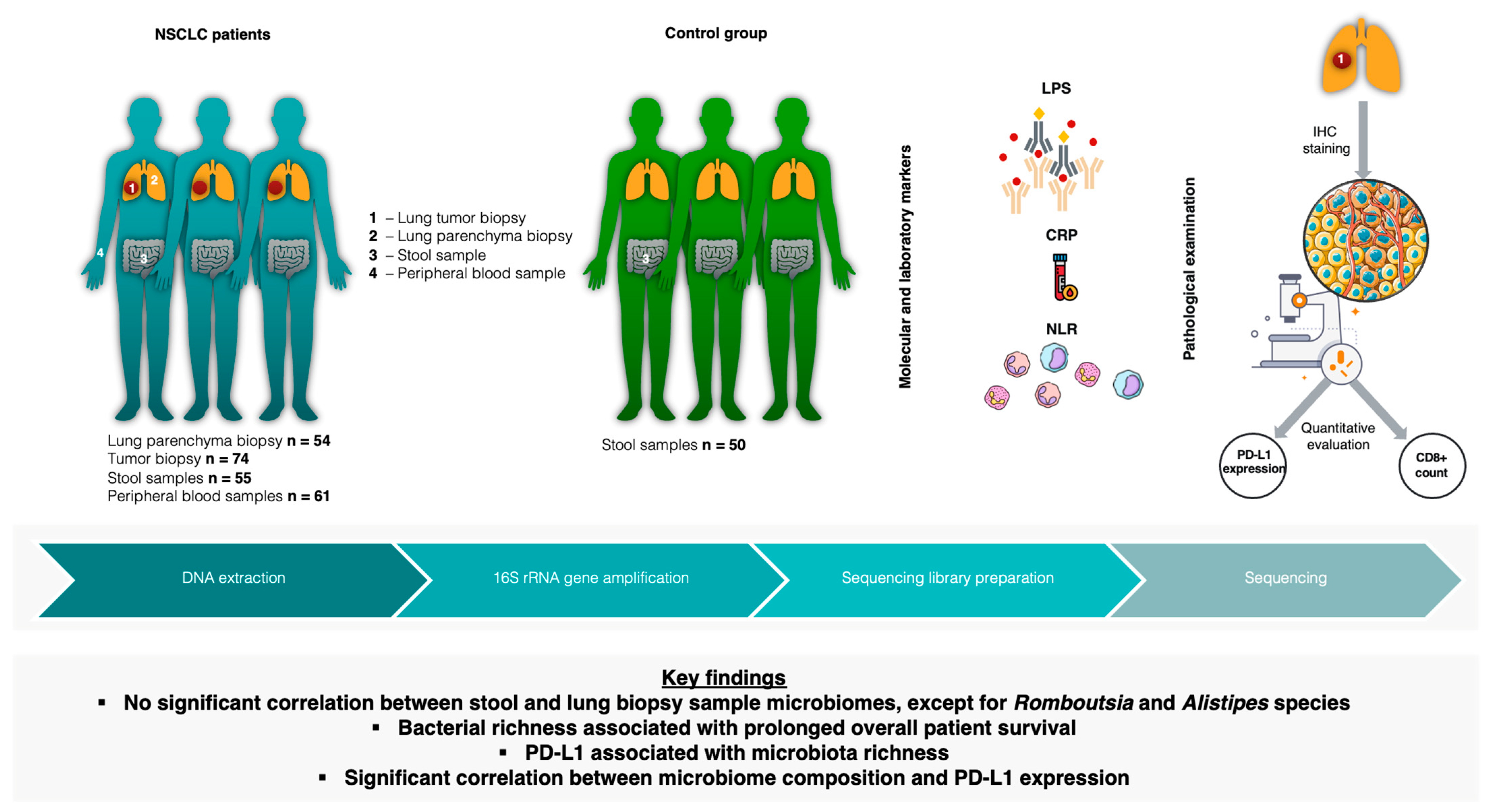{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
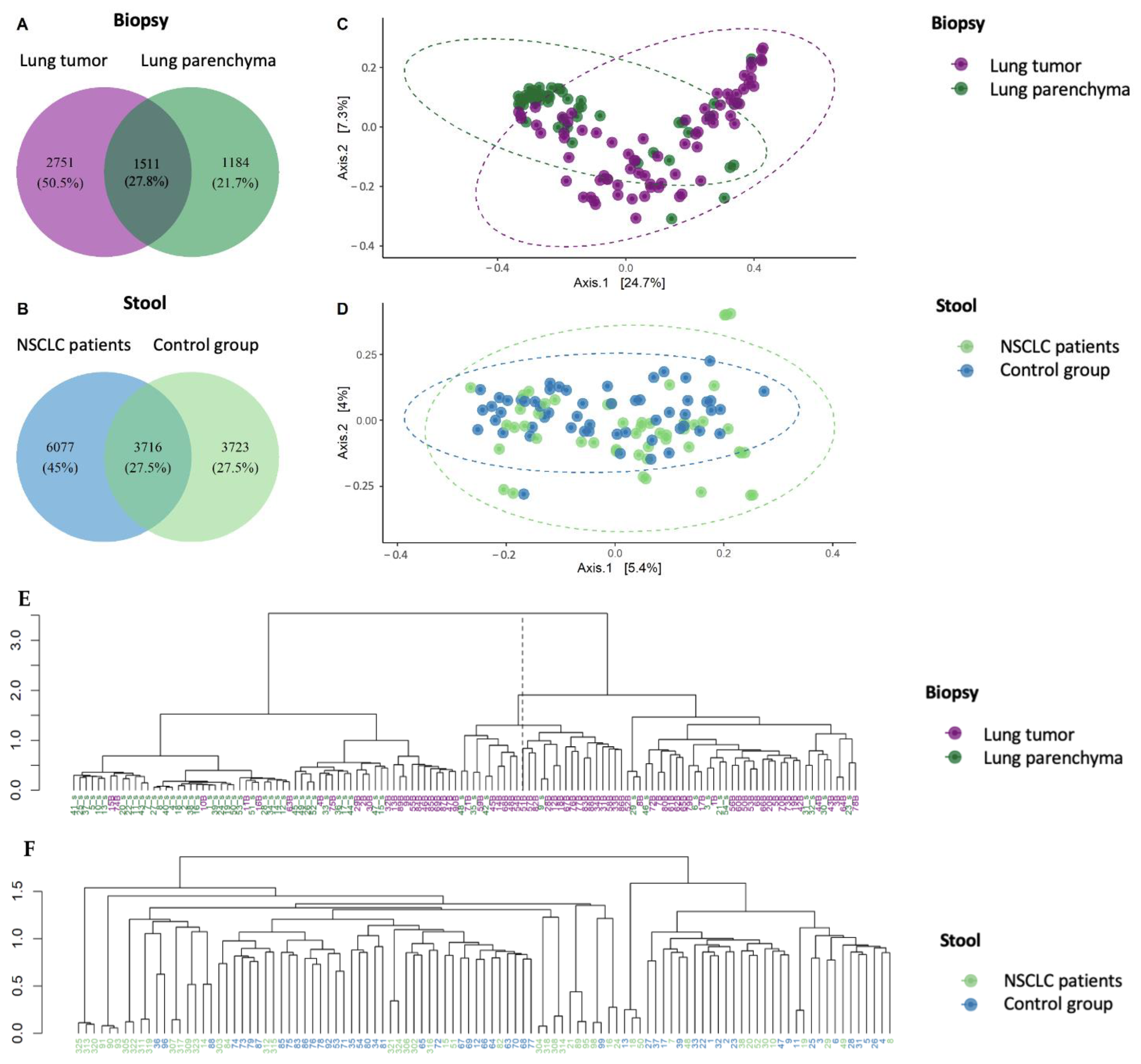
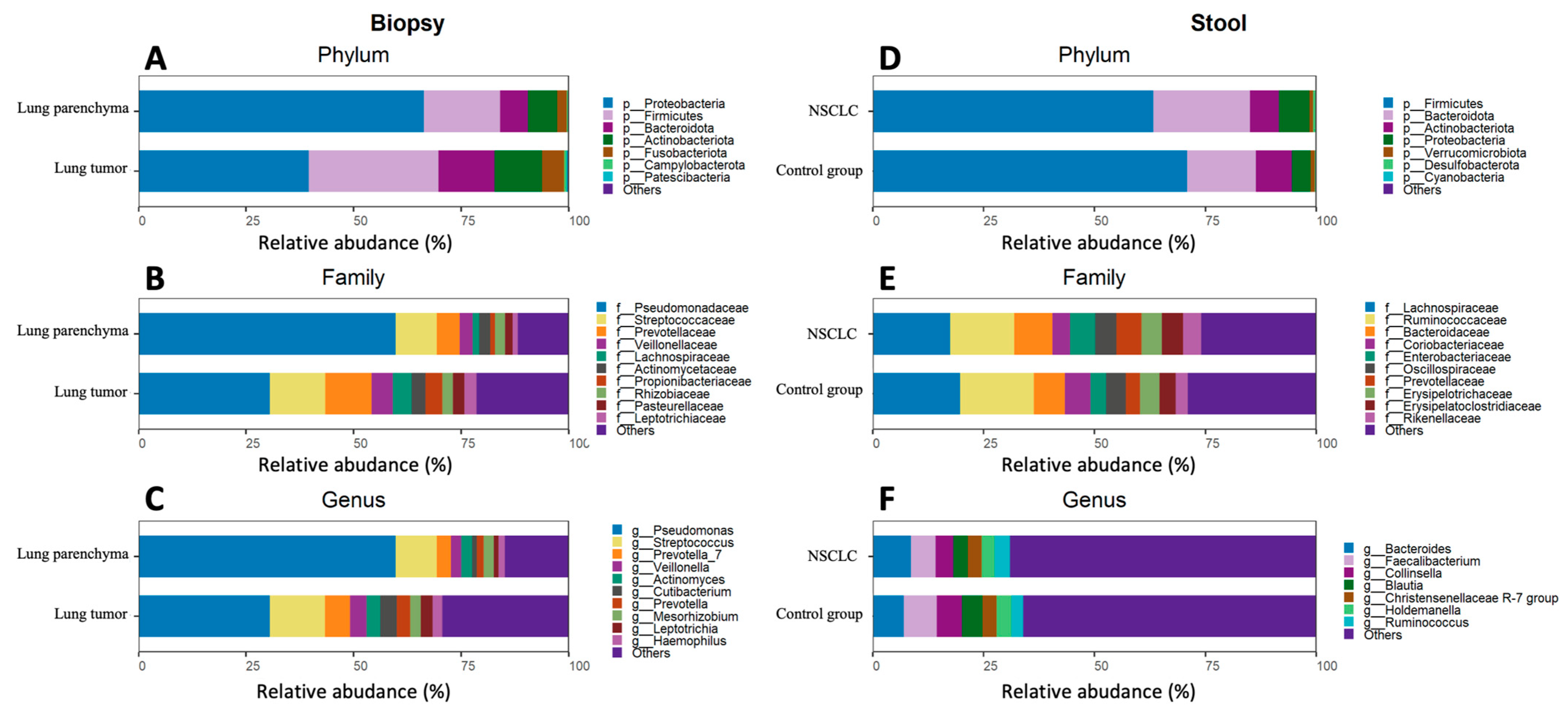
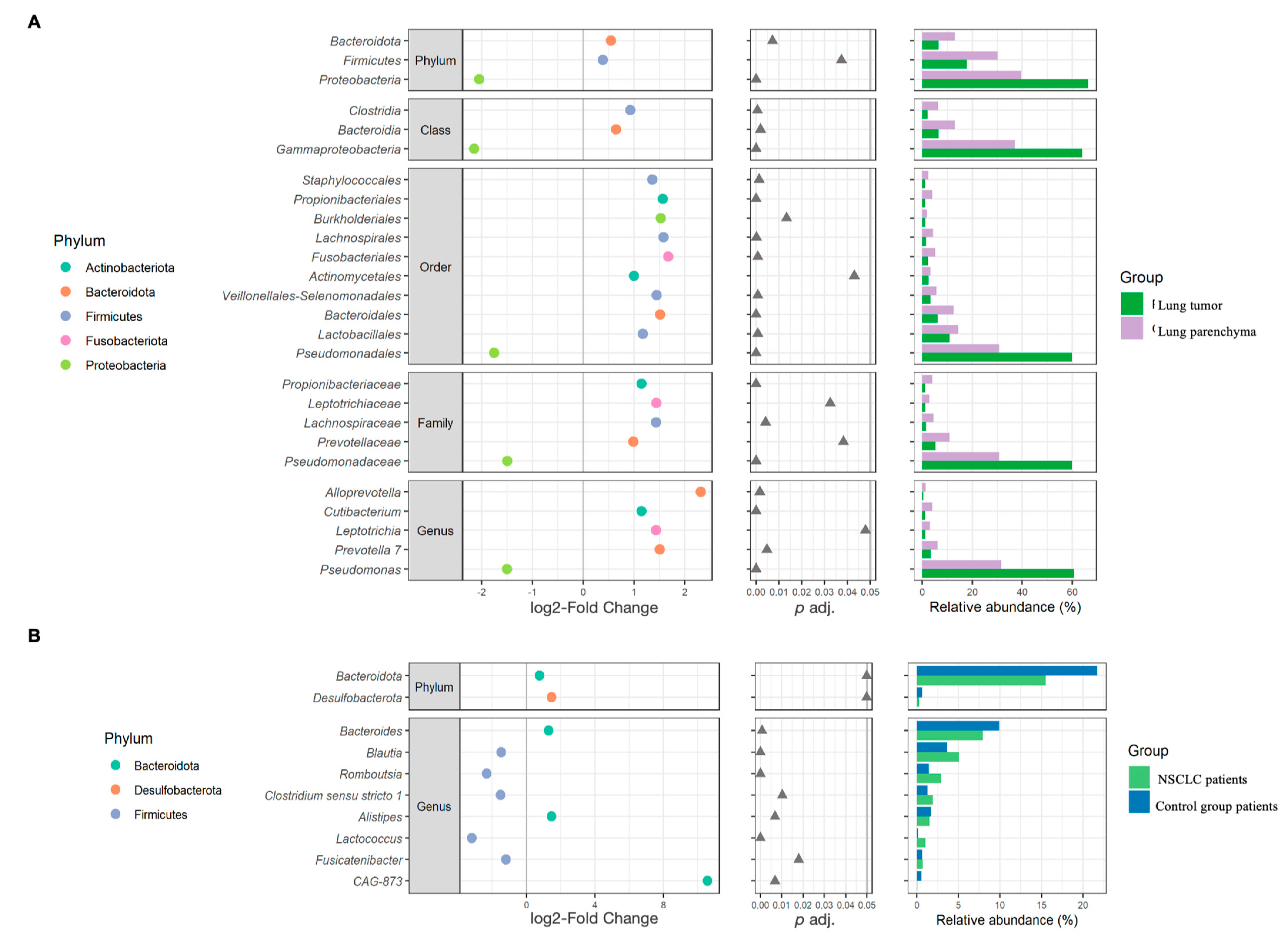
2.1. Global Bacterial Profile Changes at Both Lung and Gut Levels in NSCLC Patients
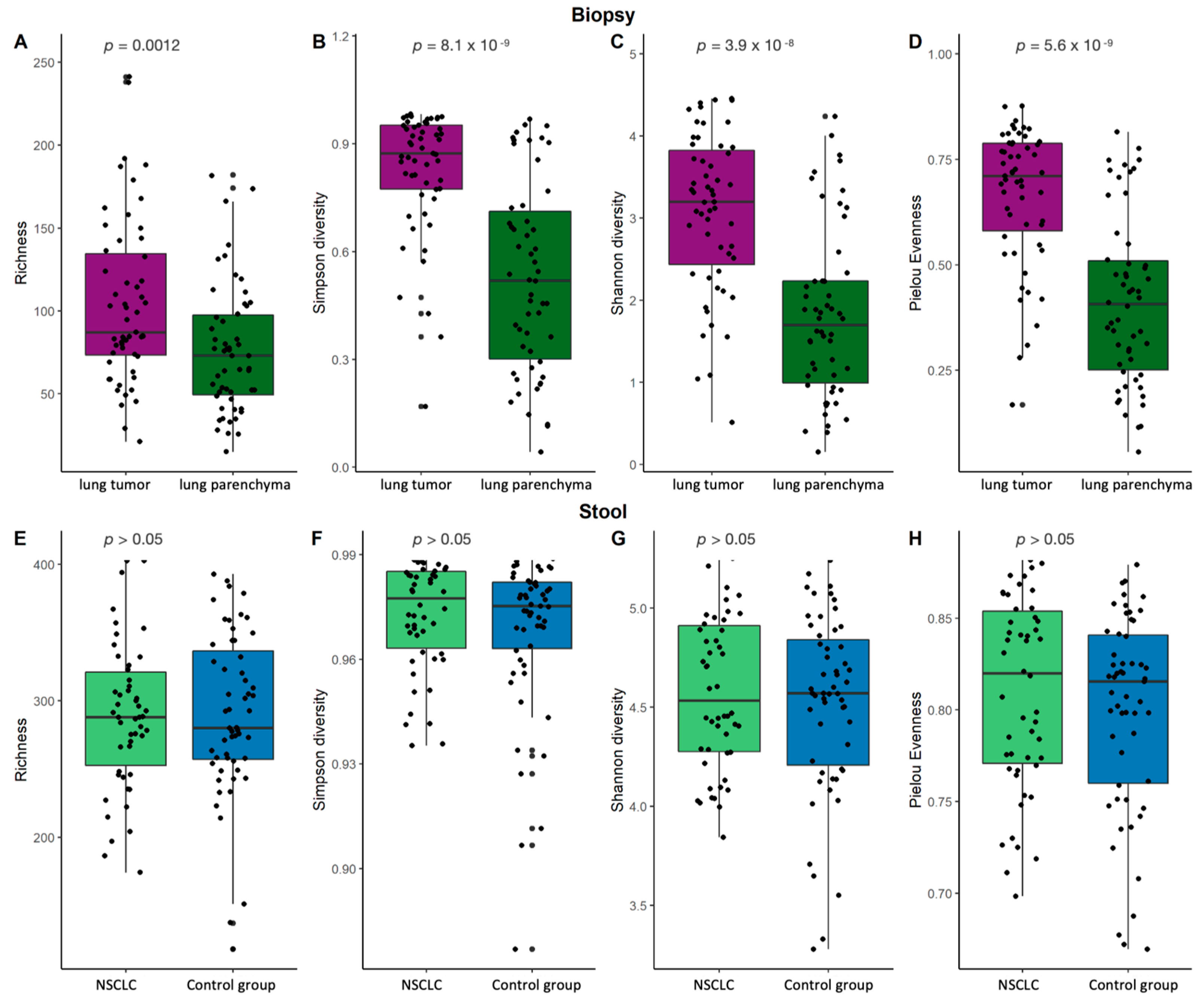
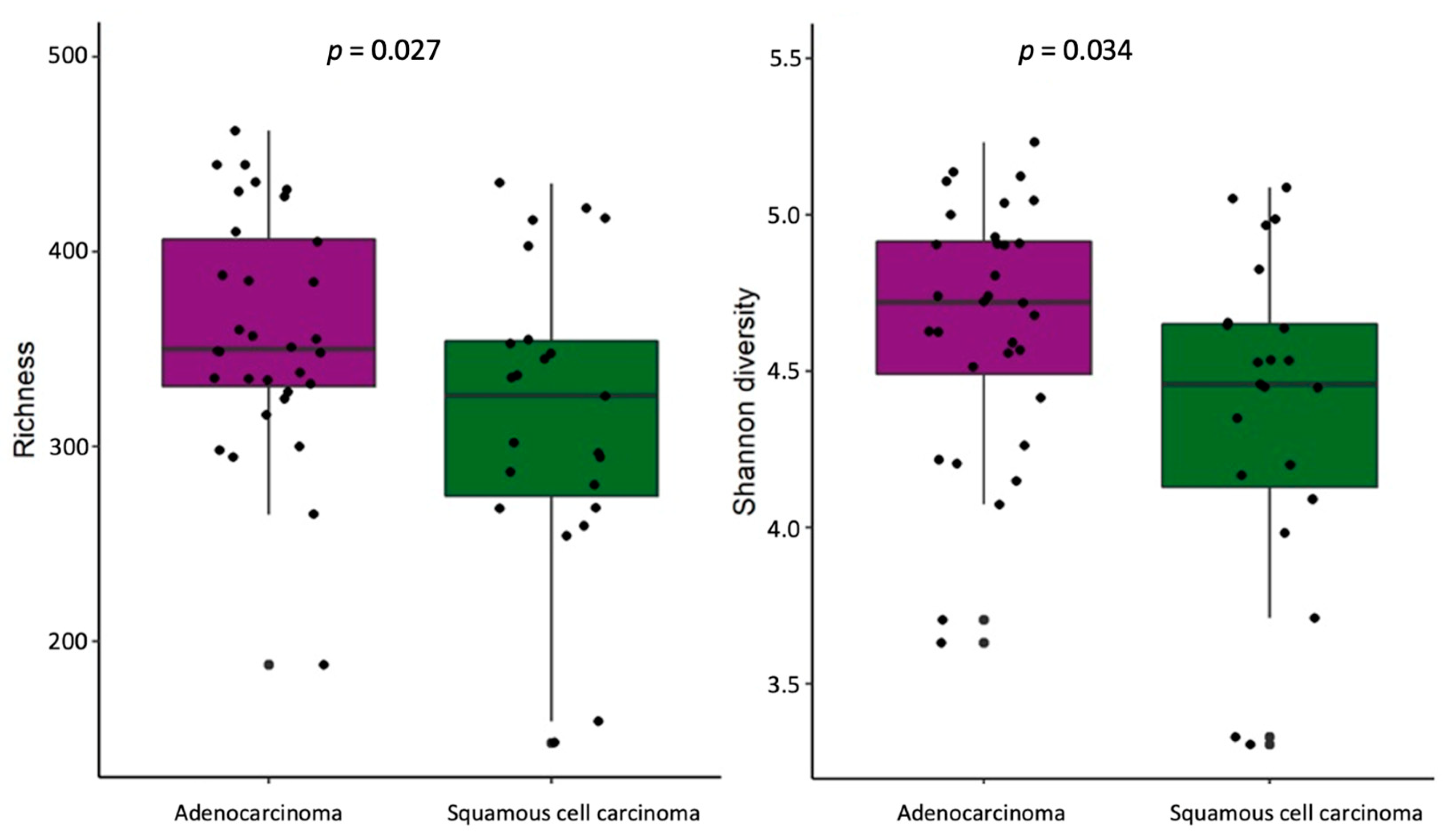
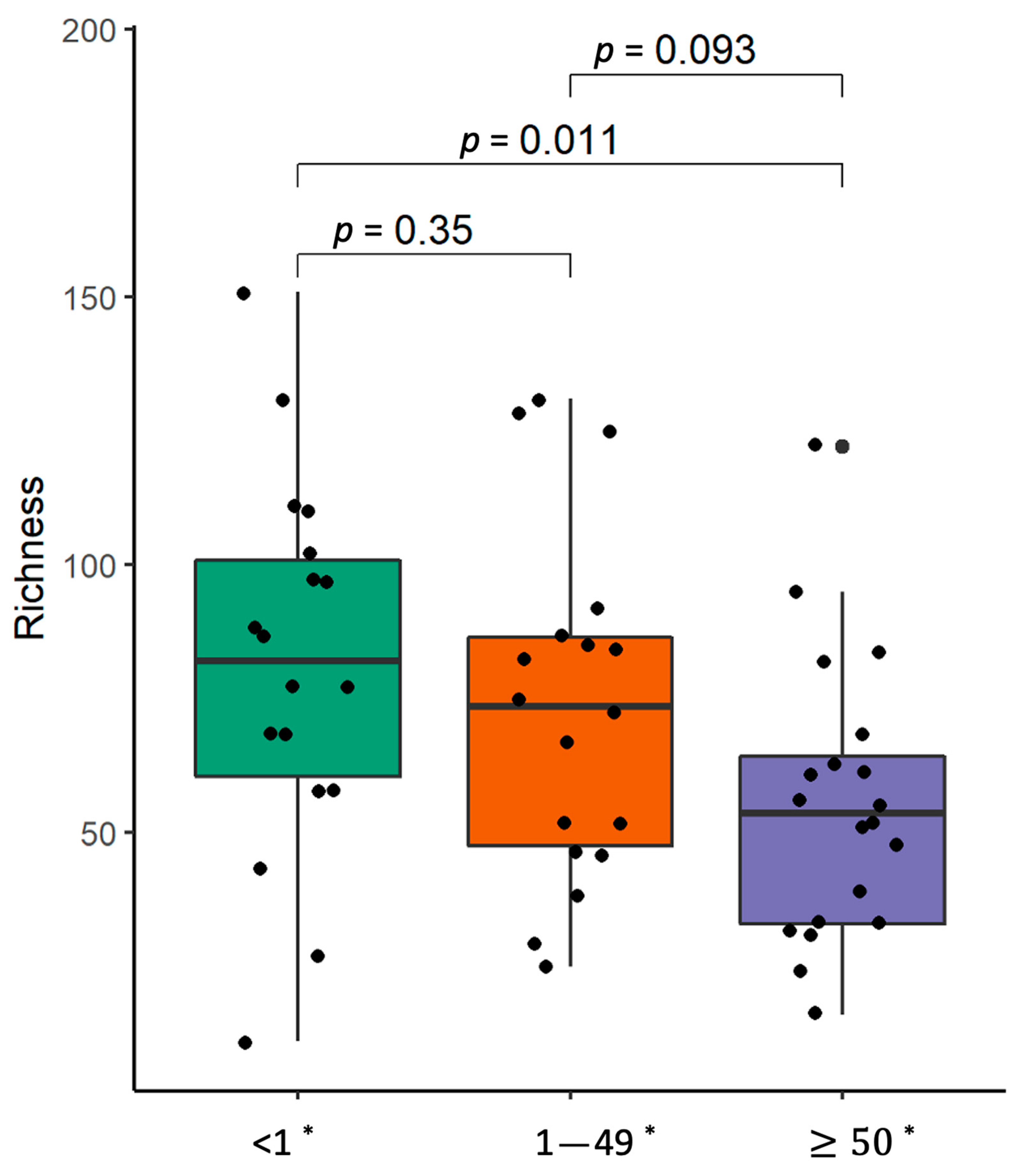
2.2. Bacterial α-Diversity and Compositional Changes in NSCLC Patients at Lung and Gut Levels
2.3. Evaluation of Lung–Gut Axis at the Microbiome Compositional Level
2.4. Differences in Bacteria Abundance in Relation to NSCLC Clinical Data
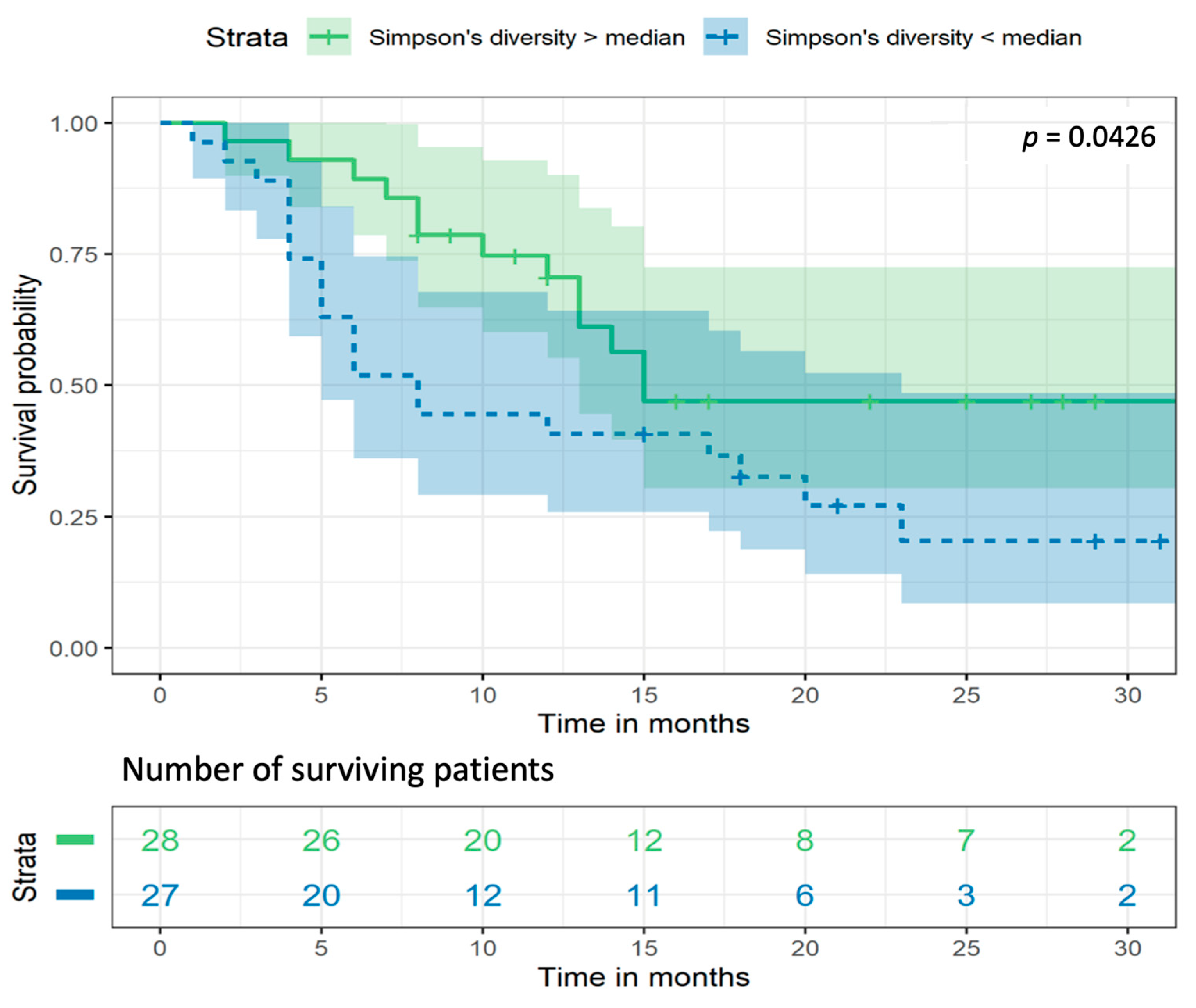
2.5. Gut Bacterial Diversity Is Associated with Survival Rate of NSCLC Patients
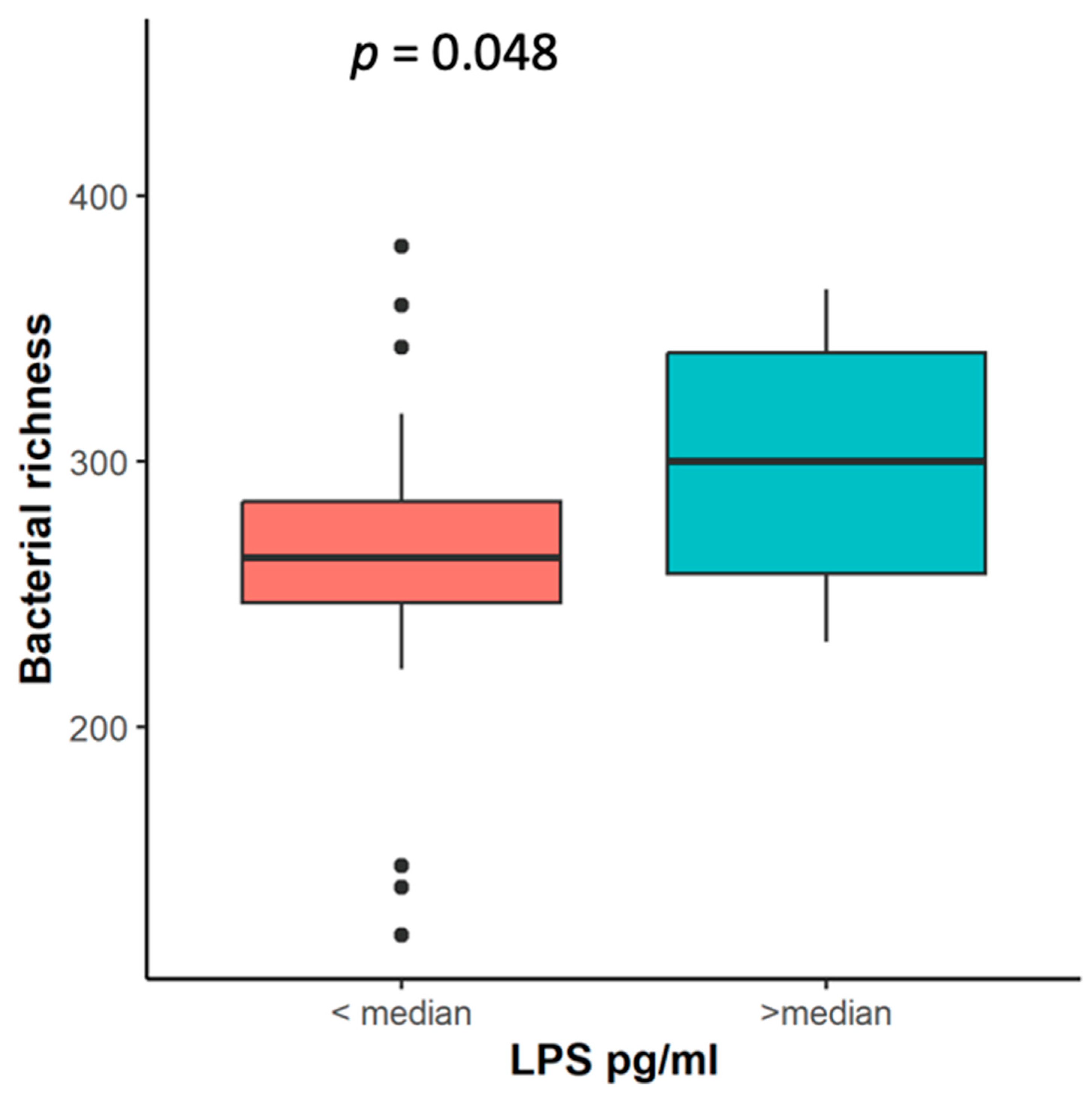
2.6. Association of Systemic Inflammation Markers LPS, NLR, and CRP with Microbiota Changes
2.7. PD-L1 Expression in Malignant Lung Tissue
3. Discussion
4. Materials and Methods
4.1. Study Cohort
4.2. Biopsy and Stool Sample Collection
4.3. Pathological Examination
4.4. Blood Sample Collection and Biomarker Testing
4.5. DNA Extraction and Library Preparation
4.6. Bioinformatic and Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovaleva, O.V.; Romashin, D.; Zborovskaya, I.B.; Davydov, M.M.; Shogenov, M.S.; Gratchev, A. Human Lung Microbiome on the Way to Cancer. J. Immunol. Res. 2019, 2019, 1394191. [Google Scholar] [CrossRef]
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the Human Microbiome. Nutr. Rev. 2012, 70 (Suppl. S1), S38–S44. [Google Scholar] [CrossRef] [PubMed]
- Vilchez-Vargas, R.; Skieceviciene, J.; Lehr, K.; Varkalaite, G.; Thon, C.; Urba, M.; Morkūnas, E.; Kucinskas, L.; Bauraite, K.; Schanze, D.; et al. Gut Microbial Similarity in twins is Driven by shared Environment and Aging. eBioMedicine 2022, 79, 104011. [Google Scholar] [CrossRef]
- Lehr, K.; Nikitina, D.; Vilchez-Vargas, R.; Steponaitiene, R.; Thon, C.; Skieceviciene, J.; Schanze, D.; Zenker, M.; Malfertheiner, P.; Kupcinskas, J.; et al. Microbial Composition of Tumorous and Adjacent Gastric Tissue Is Associated with Prognosis of Gastric Cancer. Sci. Rep. 2023, 13, 4640. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the Human Gut Microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Morkūnas, E.; Skiecevičienė, J.; Kupčinskas, J. The Impact of Modulating the Gastrointestinal Microbiota in Cancer Patients. Best Pract. Res. Clin. Gastroenterol. 2020, 48–49, 101700. [Google Scholar] [CrossRef] [PubMed]
- Nikitina, D.; Lehr, K.; Vilchez-Vargas, R.; Jonaitis, L.V.; Urba, M.; Kupcinskas, J.; Skieceviciene, J.; Link, A. Comparison of Genomic and Transcriptional Microbiome Analysis in Gastric Cancer Patients and Healthy Individuals. World J. Gastroenterol. 2023, 29, 1202–1218. [Google Scholar] [CrossRef] [PubMed]
- Smet, A.; Kupcinskas, J.; Link, A.; Hold, G.L.; Bornschein, J. The Role of Microbiota in Gastrointestinal Cancer and Cancer Treatment: Chance or Curse? Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 857–874. [Google Scholar] [CrossRef]
- Helmink, B.A.; Khan, M.A.W.; Hermann, A.; Gopalakrishnan, V.; Wargo, J.A. The microbiome, cancer, and cancer therapy. Nat. Med. 2019, 25, 377–388. [Google Scholar] [CrossRef]
- Zhang, T.; Joubert, P.; Ansari-Pour, N.; Zhao, W.; Hoang, P.H.; Lokanga, R.; Moye, A.L.; Rosenbaum, J.; Gonzalez-Perez, A.; Martínez-Jiménez, F.; et al. Genomic and Evolutionary Classification of Lung Cancer in Never Smokers. Nat. Genet. 2021, 53, 1348–1359. [Google Scholar] [CrossRef]
- Shiels, M.S.; Albanes, D.; Virtamo, J.; Engels, E.A. Increased Risk of Lung Cancer in Men with Tuberculosis in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study. Cancer Epidemiol. Biomark. Prev. 2011, 20, 672–678. [Google Scholar] [CrossRef]
- Littman, A.J.; White, E.; Jackson, L.A.; Thornquist, M.D.; Gaydos, C.A.; Goodman, G.E.; Vaughan, T.L. Chlamydia pneumoniae Infection and Risk of Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1624–1630. [Google Scholar] [CrossRef]
- Chellappan, D.K.; Ning, Q.L.S.; Min, S.K.S.; Bin, S.Y.; Chern, P.J.; Shi, T.P.; Mei, S.W.E.; Yee, T.H.; Qi, O.J.; Thangavelu, L.; et al. Interactions between Microbiome and Lungs: Paving New Paths for microbiome Based Bio-Engineered Drug Delivery Systems in Chronic Respiratory Diseases. Chem.-Biol. Interact. 2019, 310, 108732. [Google Scholar] [CrossRef]
- Sommariva, M.; Le Noci, V.; Bianchi, F.; Camelliti, S.; Balsari, A.; Tagliabue, E.; Sfondrini, L. The Lung Microbiota: Role in Maintaining Pulmonary Immune Homeostasis and Its Implications in Cancer Development and Therapy. Cell. Mol. Life Sci. 2020, 77, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Dickson, R.P.; Huffnagle, G.B. The Lung Microbiome: New Principles for Respiratory Bacteriology in Health and Disease. PLoS Pathog. 2015, 11, e1004923. [Google Scholar] [CrossRef]
- Yu, G.; Gail, M.H.; Consonni, D.; Carugno, M.; Humphrys, M.; Pesatori, A.C.; Caporaso, N.E.; Goedert, J.J.; Ravel, J.; Landi, M.T. Characterizing Human Lung Tissue Microbiota and Its Relationship to Epidemiological and Clinical Features. Genome Biol. 2016, 17, 163. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sung, J.Y.; Yong, D.; Chun, J.; Kim, S.Y.; Song, J.H.; Chung, K.S.; Kim, E.Y.; Jung, J.Y.; Kang, Y.A.; et al. Characterization of Microbiome in Bronchoalveolar Lavage Fluid of Patients with Lung Cancer Comparing with Benign Mass like Lesions. Lung Cancer 2016, 102, 89–95. [Google Scholar] [CrossRef]
- Laroumagne, S.; Lepage, B.; Hermant, C.; Plat, G.; Phelippeau, M.; Bigay-Game, L.; Lozano, S.; Guibert, N.; Segonds, C.; Mallard, V.; et al. Bronchial Colonisation in Patients with Lung Cancer: A Prospective Study. Eur. Respir. J. 2013, 42, 220–229. [Google Scholar] [CrossRef]
- Stasiewicz, M.; Karpiński, T.M. The Oral Microbiota and Its Role in Carcinogenesis. Semin. Cancer Biol. 2022, 86, 633–642. [Google Scholar] [CrossRef]
- Dickson, R.P.; Erb-Downward, J.R.; Freeman, C.M.; McCloskey, L.; Falkowski, N.R.; Huffnagle, G.B.; Curtis, J.L. Bacterial Topography of the Healthy Human Lower Respiratory Tract. mBio 2017, 8, e02287-16. [Google Scholar] [CrossRef]
- Zeineldin, M.M.; Lowe, J.F.; Grimmer, E.D.; de Godoy, M.R.C.; Ghanem, M.M.; El-Raof, Y.M.A.; Aldridge, B.M. Relationship between Nasopharyngeal and Bronchoalveolar Microbial Communities in Clinically Healthy Feedlot Cattle. BMC Microbiol. 2017, 17, 138. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, T.; Xu, Q. Therapeutic Advances in Non-Small Cell Lung Cancer: Focus on Clinical Development of Targeted Therapy and Immunotherapy. MedComm 2020, 2, 692–729. [Google Scholar] [CrossRef]
- Roviello, G.; Iannone, L.F.; Bersanelli, M.; Mini, E.; Catalano, M. The Gut Microbiome and Efficacy of Cancer Immunotherapy. Pharmacol. Ther. 2022, 231, 107973. [Google Scholar] [CrossRef]
- Jang, H.J.; Choi, J.Y.; Kim, K.; Yong, S.H.; Kim, Y.W.; Kim, S.Y.; Kim, E.Y.; Jung, J.Y.; Kang, Y.A.; Park, M.S.; et al. Relationship of the Lung Microbiome with PD-L1 Expression and Immunotherapy Response in Lung Cancer. Respir. Res. 2021, 22, 322. [Google Scholar] [CrossRef]
- McQuade, J.L.; Ologun, G.O.; Arora, R.; Wargo, J.A. Gut Microbiome Modulation Via Fecal Microbiota Transplant to Augment Immunotherapy in Patients with Melanoma or Other Cancers. Curr. Oncol. Rep. 2020, 22, 74. [Google Scholar] [CrossRef] [PubMed]
- Borgers, J.S.W.; Burgers, F.H.; Terveer, E.M.; van Leerdam, M.E.; Korse, C.M.; Kessels, R.; Flohil, C.C.; Blank, C.U.; Schumacher, T.N.; van Dijk, M.; et al. Conversion of Unresponsiveness to Immune Checkpoint Inhibition by Fecal Microbiota Transplantation in Patients with Metastatic Melanoma: Study Protocol for a Randomized Phase Ib/IIa Trial. BMC Cancer 2022, 22, 1366. [Google Scholar] [CrossRef]
- Xia, L.; Liu, Y.; Wang, Y. PD-1/PD-L1 Blockade Therapy in Advanced Non-Small-Cell Lung Cancer: Current Status and Future Directions. Oncologist 2019, 24, S31–S41. [Google Scholar] [CrossRef] [PubMed]
- Zuo, B.; Li, T.; Liu, X.; Wang, S.; Cheng, J.; Liu, X.; Cui, W.; Shi, H.; Ling, C. Dipeptidyl Peptidase 4 Inhibitor Reduces Tumor-associated Macrophages and Enhances Anti-PD-L1-Mediated Tumor Suppression in Non-Small Cell Lung Cancer. Clin. Transl. Oncol. 2023, 25, 3188–3202. [Google Scholar] [CrossRef]
- Peters, B.A.; Hayes, R.B.; Goparaju, C.; Reid, C.; Pass, H.I.; Ahn, J. The Microbiome in Lung Cancer Tissue and Recurrence-Free Survival. Cancer Epidemiol. Biomark. Prev. 2019, 28, 731–740. [Google Scholar] [CrossRef]
- Liu, H.; Tao, L.; Zhang, J.; Zhu, Y.; Zheng, Y.; Liu, D.; Zhou, M.; Ke, H.; Shi, M.; Qu, J. Difference of Lower Airway Microbiome in Bilateral Protected Specimen Brush between Lung Cancer Patients with Unilateral Lobar Masses and Control subjects. Int. J. Cancer 2018, 142, 769–778. [Google Scholar] [CrossRef]
- Bello, S.; Vengoechea, J.J.; Ponce-Alonso, M.; Figueredo, A.L.; Mincholé, E.; Rezusta, A.; Gambó, P.; Pastor, J.M.; Galeano, J.; del Campo, R. Core Microbiota in Central Lung Cancer with Streptococcal Enrichment as a Possible Diagnostic Marker. Arch. Bronconeumol. 2020, 57, 681–689. [Google Scholar] [CrossRef]
- Greathouse, K.L.; White, J.R.; Vargas, A.J.; Bliskovsky, V.V.; Beck, J.A.; Von Muhlinen, N.; Polley, E.C.; Bowman, E.D.; Khan, M.A.; Robles, A.I.; et al. Interaction between the Microbiome and TP53 in Human Lung Cancer. Genome Biol. 2018, 19, 123. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Zhao, C.; Yu, M.; Chen, H.; Pan, Y.; Wang, Y.; Bao, H.; Ma, H.; Ma, S. Alterations of Lung Microbiota in Patients with Non-Small Cell Lung Cancer. Bioengineered 2022, 13, 6665–6677. [Google Scholar] [CrossRef]
- Islam, Z.; Tran, M.; Xu, T.; Tierney, B.T.; Patel, C.; Kostic, A.D. Reproducible and Opposing Gut Microbiome Signatures Distinguish Autoimmune Diseases and Cancers: A Systematic Review and Meta-Analysis. Microbiome 2022, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; An, R.; Wang, L.; Shan, J.; Wang, X. Specific Gut Microbiome and Serum Metabolome Changes in Lung Cancer Patients. Front. Cell. Infect. Microbiol. 2021, 11, 725284. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, J.; Guan, Y.; Lou, Y.; Chen, H.; Xu, M.; Deng, D.; Chen, J.; Ni, B.; Zhao, L.; et al. Dysbiosis of the Gut Microbiome is associated with Tumor Biomarkers in Lung Cancer. Int. J. Biol. Sci. 2019, 15, 2381–2392. [Google Scholar] [CrossRef]
- Lu, H.; Gao, N.L.; Tong, F.; Wang, J.; Li, H.; Zhang, R.; Ma, H.; Yang, N.; Zhang, Y.; Wang, Y.; et al. Alterations of the Human Lung and Gut Microbiomes in Non-Small Cell Lung Carcinomas and Distant Metastasis. Microbiol. Spectr. 2021, 9, e0080221. [Google Scholar] [CrossRef]
- Dumont-Leblond, N.; Veillette, M.; Racine, C.; Joubert, P.; Duchaine, C. Non-Small Cell Lung Cancer Microbiota Characterization: Prevalence of Enteric and Potentially Pathogenic Bacteria in Cancer Tissues. PLoS ONE 2021, 16, e0249832. [Google Scholar] [CrossRef]
- Goto, T. Microbiota and Lung Cancer. Semin. Cancer Biol. 2022, 86, 1–10. [Google Scholar] [CrossRef]
- Zheng, Y.; Fang, Z.; Xue, Y.; Zhang, J.; Zhu, J.; Gao, R.; Yao, S.; Ye, Y.; Wang, S.; Lin, C.; et al. Specific Gut Microbiome Signature predicts the Early-Stage Lung Cancer. Gut Microbes 2020, 11, 1030–1042. [Google Scholar] [CrossRef]
- Qin, X.; Bi, L.; Yang, W.; He, Y.; Gu, Y.; Yang, Y.; Gong, Y.; Wang, Y.; Yan, X.; Xu, L.; et al. Dysbiosis of the Gut Microbiome Is Associated with Histopathology of Lung Cancer. Front. Microbiol. 2022, 13, 918823. [Google Scholar] [CrossRef] [PubMed]
- Kespohl, M.; Vachharajani, N.; Luu, M.; Harb, H.; Pautz, S.; Wolff, S.; Sillner, N.; Walker, A.; Schmitt-Kopplin, P.; Boettger, T.; et al. The Microbial Metabolite Butyrate Induces Expression of Th1-Associated Factors in CD4+ T Cells. Front. Immunol. 2017, 8, 1036. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H. Blautia—A New Functional Genus with Potential Probiotic Properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human Intestinal Lumen and Mucosa-Associated Microbiota in Patients with Colorectal Cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, M.; Liu, J.; Gao, R.; Hu, J.; Li, J.; Zhang, L.; Shi, Y.; Guo, H.; Cheng, J.; et al. Discovery and Validation of Potential Bacterial Biomarkers for Lung Cancer. Am. J. Cancer Res. 2015, 5, 3111–3122. [Google Scholar]
- Zhao, H.; Chu, M.; Huang, Z.; Yang, X.; Ran, S.; Hu, B.; Zhang, C.; Liang, J. Variations in Oral Microbiota Associated with Oral Cancer. Sci. Rep. 2017, 7, 11773. [Google Scholar] [CrossRef]
- Furuya, R.; Onoye, Y.; Kanayama, A.; Saika, T.; Iyoda, T.; Tatewaki, M.; Matsuzaki, K.; Kobayashi, I.; Tanaka, M. Antimicrobial Resistance in Clinical Isolates of Neisseria Subflava from the oral Cavities of a Japanese Population. J. Infect. Chemother. 2007, 13, 302–304. [Google Scholar] [CrossRef]
- Li, L.; Mac Aogáin, M.; Xu, T.; Jaggi, T.K.; Chan, L.L.; Qu, J.; Wei, L.; Liao, S.; Cheng, H.S.; Keir, H.R.; et al. Neisseria Species as Pathobionts in Bronchiectasis. Cell Host Microbe 2022, 30, 1311–1327.e8. [Google Scholar] [CrossRef]
- Qin, S.; Clausen, E.; Lucht, L.; Michael, H.; Beck, J.M.; Curtis, J.L.; Freeman, C.M.; Morris, A. Presence of Tropheryma whipplei in Different Body Sites in a Cohort of Healthy Subjects. Am. J. Respir. Crit. Care Med. 2016, 194, 243–245. [Google Scholar] [CrossRef]
- Dolmans, R.A.V.; Boel, C.H.E.; Lacle, M.M.; Kusters, J.G. Clinical Manifestations, Treatment, and Diagnosis of Tropheryma whipplei Infections. Clin. Microbiol. Rev. 2017, 30, 529–555. [Google Scholar] [CrossRef]
- Lozupone, C.; Cota-Gomez, A.; Palmer, B.E.; Linderman, D.J.; Charlson, E.S.; Sodergren, E.; Mitreva, M.; Abubucker, S.; Martin, J.; Yao, G.; et al. Widespread Colonization of the Lung by Tropheryma whipplei in HIV Infection. Am. J. Respir. Crit. Care Med. 2013, 187, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Weyler, L.; Engelbrecht, M.; Forsberg, M.M.; Brehwens, K.; Vare, D.; Vielfort, K.; Wojcik, A.; Aro, H. Restriction Endonucleases from Invasive Neisseria gonorrhoeae Cause Double-Strand Breaks and Distort Mitosis in Epithelial Cells during Infection. PLoS ONE 2014, 9, e114208. [Google Scholar] [CrossRef] [PubMed]
- Khatun, S.; Appidi, T.; Rengan, A.K. The Role Played by Bacterial infections in the Onset and Metastasis of Cancer. Curr. Res. Microb. Sci. 2021, 2, 100078. [Google Scholar] [CrossRef] [PubMed]
- Chang, N.-W.; Pei, R.-J.; Tseng, H.-C.; Yeh, K.-T.; Chan, H.-C.; Lee, M.-R.; Lin, C.; Hsieh, W.-T.; Kao, M.-C.; Tsai, M.-H.; et al. Co-Treating with Arecoline and 4-Nitroquinoline 1-Oxide to Establish a Mouse Model Mimicking Oral Tumorigenesis. Chem.-Biol. Interact. 2010, 183, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.N.; Clemente, J.C.; Tsay, J.-C.J.; Koralov, S.B.; Keller, B.C.; Wu, B.G.; Li, Y.; Shen, N.; Ghedin, E.; Morris, A.; et al. Enrichment of the Lung Microbiome with Oral Taxa Is Associated with Lung Inflammation of a Th17 Phenotype. Nat. Microbiol. 2016, 1, 16031. [Google Scholar] [CrossRef]
- Cheng, M.; Hu, S. Lung-Resident γδ T Cells and Their Roles in Lung Diseases. Immunology 2017, 151, 375–384. [Google Scholar] [CrossRef]
- Wang, J.; Jia, H. Metagenome-Wide Association Studies: Fine-Mining the Microbiome. Nat. Rev. Microbiol. 2016, 14, 508–522. [Google Scholar] [CrossRef]
- He, J.-Q.; Chen, Q.; Wu, S.-J.; Wang, D.-Q.; Zhang, S.-Y.; Zhang, S.-Z.; Chen, R.-L.; Wang, J.-F.; Wang, Z.; Yu, C.-H. Potential Implications of the Lung Microbiota in Patients with Chronic Obstruction Pulmonary Disease and Non-Small Cell Lung Cancer. Front. Cell. Infect. Microbiol. 2022, 12, 937864. [Google Scholar] [CrossRef]
- Millares, L.; Pérez-Brocal, V.; Ferrari, R.; Gallego, M.; Pomares, X.; García-Núñez, M.; Montón, C.; Capilla, S.; Monsó, E.; Moya, A. Functional Metagenomics of the Bronchial Microbiome in COPD. PLoS ONE 2015, 10, e0144448. [Google Scholar] [CrossRef] [PubMed]
- Downes, J.; Dewhirst, F.E.; Tanner, A.C.R.; Wade, W.G. Description of Alloprevotella rava gen. nov., sp. nov., isolated from the human oral cavity, and reclassification of Prevotella tannerae Moore et al. 1994 as Alloprevotella tannerae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1214–1218. [Google Scholar] [CrossRef]
- Santagati, M.; Scillato, M.; Patanè, F.; Aiello, C.; Stefani, S. Bacteriocin-Producing Oral Streptococci and Inhibition of Respiratory Pathogens. FEMS Immunol. Med. Microbiol. 2012, 65, 23–31. [Google Scholar] [CrossRef]
- Srikham, K.; Daengprok, W.; Niamsup, P.; Thirabunyanon, M. Characterization of Streptococcus salivarius as New Probiotics Derived from Human Breast Milk and Their Potential on Proliferative Inhibition of Liver and Breast Cancer Cells and Antioxidant Activity. Front. Microbiol. 2021, 12, 797445. [Google Scholar] [CrossRef]
- Page, M.J.; Kell, D.B.; Pretorius, E. The Role of Lipopolysaccharide-Induced Cell Signalling in Chronic Inflammation. Chronic Stress 2022, 6, 24705470221076390. [Google Scholar] [CrossRef] [PubMed]
- Brewster, W.R.; Burkett, W.C.; Ko, E.M.; Bae-Jump, V.; McCoy, A.N.; Keku, T.O. An Evaluation of the Microbiota of the Upper Reproductive Tract of Women with and without Epithelial Ovarian Cancer. Gynecol. Oncol. Rep. 2022, 42, 101017. [Google Scholar] [CrossRef] [PubMed]
- Heidler, S.; Lusuardi, L.; Madersbacher, S.; Freibauer, C. The Microbiome in Benign Renal Tissue and in Renal Cell Carcinoma. Urol. Int. 2020, 104, 247–252. [Google Scholar] [CrossRef]
- Chen, Q.; Hou, K.; Tang, M.; Ying, S.; Zhao, X.; Li, G.; Pan, J.; He, X.; Xia, H.; Li, Y.; et al. Screening of Potential Microbial Markers for Lung Cancer Using Metagenomic Sequencing. Cancer Med. 2023, 12, 7127–7139. [Google Scholar] [CrossRef]
- Lamaudière, M.T.F.; Arasaradnam, R.; Weedall, G.D.; Morozov, I.Y. The Colorectal Cancer Gut Environment Regulates Activity of the Microbiome and Promotes the Multidrug Resistant Phenotype of ESKAPE and Other Pathogens. mSphere 2023, 8, e0062622. [Google Scholar] [CrossRef]
- Wu, X.; Qian, S.; Zhang, J.; Feng, J.; Luo, K.; Sun, L.; Zhao, L.; Ran, Y.; Sun, L.; Wang, J.; et al. Lipopolysaccharide Promotes Metastasis via Acceleration of Glycolysis by the Nuclear Factor-κB/Snail/Hexokinase3 Signaling Axis in Colorectal Cancer. Cancer Metab. 2021, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Kim, S.; Keam, B.; Kim, T.M.; Kim, D.-W.; Heo, D.S. Soluble PD-L1 Is a Predictive and Prognostic Biomarker in Advanced Cancer Patients Who Receive Immune Checkpoint Blockade Treatment. Sci. Rep. 2021, 11, 19712. [Google Scholar] [CrossRef]
- Tian, P.; He, B.; Mu, W.; Liu, K.; Liu, L.; Zeng, H.; Liu, Y.; Jiang, L.; Zhou, P.; Huang, Z.; et al. Assessing PD-L1 Expression in Non-Small Cell Lung Cancer and Predicting Responses to Immune Checkpoint Inhibitors Using Deep Learning on Computed Tomography Images. Theranostics 2021, 11, 2098–2107. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti-PD-L1 Efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, F.; Gertz, R.E.; Park, S.H.; Kim, E.; Moura, I.; Milucky, J.; Rouphael, N.; Farley, M.M.; Harrison, L.H.; Bennett, N.M.; et al. Streptococcus infantis, Streptococcus mitis, and Streptococcus oralis Strains with Highly Similar cps5 Loci and Antigenic Relatedness to Serotype 5 Pneumococci. Front. Microbiol. 2018, 9, 3199. [Google Scholar] [CrossRef] [PubMed]
- Engen, S.A.; Schreurs, O.; Petersen, F.; Blix, I.J.S.; Baekkevold, E.S.; Schenck, K. The Regulatory Role of the Oral Commensal Streptococcus mitis on Human Monocytes. Scand. J. Immunol. 2018, 87, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, M.C.; Lefimil, C.; Rodas, P.I.; Vernal, R.; Lopez, M.; Acuña-Castillo, C.; Imarai, M.; Escobar, A. Neisseria gonorrhoeae Modulates Immunity by Polarizing Human Macrophages to a M2 Profile. PLoS ONE 2015, 10, e0130713. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, J.; Sun, Z.; Cao, Y.; Mu, Z.; Ji, X. Commensal Microbiota Contributes to Predicting the Response to Immune Checkpoint Inhibitors in Non-Small-Cell Lung Cancer Patients. Cancer Sci. 2021, 112, 3005–3017. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Trinchieri, G. Microbiota: A Key Orchestrator of Cancer Therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Dong, H.; Xia, L.; Yang, Y.; Zhu, Y.; Shen, Y.; Zheng, H.; Yao, C.; Wang, Y.; Lu, S. The Diversity of Gut Microbiome is Associated with Favorable Responses to Anti–Programmed Death 1 Immunotherapy in Chinese Patients With NSCLC. J. Thorac. Oncol. 2019, 14, 1378–1389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Bai, C.; Hu, T.; Luo, C.; Yu, H.; Ma, X.; Liu, T.; Gu, X. Emerging Trends and Hotspot in Gut–Lung Axis Research from 2011 to 2021: A Bibliometrics Analysis. Biomed. Eng. Online 2022, 21, 27. [Google Scholar] [CrossRef]
- Budden, K.F.; Gellatly, S.L.; Wood, D.L.A.; Cooper, M.A.; Morrison, M.; Hugenholtz, P.; Hansbro, P.M. Emerging Pathogenic Links between Microbiota and the Gut–Lung Axis. Nat. Rev. Microbiol. 2017, 15, 55–63. [Google Scholar] [CrossRef]
- Dai, Z.; Coker, O.O.; Nakatsu, G.; Wu, W.K.K.; Zhao, L.; Chen, Z.; Chan, F.K.L.; Kristiansen, K.; Sung, J.J.Y.; Wong, S.H.; et al. Multi-Cohort Analysis of Colorectal Cancer Metagenome Identified Altered Bacteria across Populations and Universal Bacterial Markers. Microbiome 2018, 6, 70. [Google Scholar] [CrossRef]
- Liu, J.; Huang, X.; Chen, C.; Wang, Z.; Huang, Z.; Qin, M.; He, F.; Tang, B.; Long, C.; Hu, H.; et al. Identification of Colorectal Cancer Progression-Associated Intestinal Microbiome and Predictive Signature Construction. J. Transl. Med. 2023, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Ricaboni, D.; Mailhe, M.; Khelaifia, S.; Raoult, D.; Million, M. Romboutsia timonensis, a New Species Isolated from Human Gut. New Microbes New Infect. 2016, 12, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; De’angelis, N.; De’angelis, G.L.; van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal Microbiota of Intestinal Polyps Reveals Putative Biomarkers of Colorectal Cancer. Sci. Rep. 2018, 8, 13974. [Google Scholar] [CrossRef]
- Ankudavicius, V.; Miliauskas, S.; Poskiene, L.; Vajauskas, D.; Zemaitis, M. Diagnostic Yield of Transbronchial Cryobiopsy Guided by Radial Endobronchial Ultrasound and Fluoroscopy in the Radiologically Suspected Lung Cancer: A Single Institution Prospective Study. Cancers 2022, 14, 1563. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ankudavicius, V.; Nikitina, D.; Lukosevicius, R.; Tilinde, D.; Salteniene, V.; Poskiene, L.; Miliauskas, S.; Skieceviciene, J.; Zemaitis, M.; Kupcinskas, J. Detailed Characterization of the Lung–Gut Microbiome Axis Reveals the Link between PD-L1 and the Microbiome in Non-Small-Cell Lung Cancer Patients. Int. J. Mol. Sci. 2024, 25, 2323. https://doi.org/10.3390/ijms25042323
Ankudavicius V, Nikitina D, Lukosevicius R, Tilinde D, Salteniene V, Poskiene L, Miliauskas S, Skieceviciene J, Zemaitis M, Kupcinskas J. Detailed Characterization of the Lung–Gut Microbiome Axis Reveals the Link between PD-L1 and the Microbiome in Non-Small-Cell Lung Cancer Patients. International Journal of Molecular Sciences. 2024; 25(4):2323. https://doi.org/10.3390/ijms25042323
Chicago/Turabian StyleAnkudavicius, Vytautas, Darja Nikitina, Rokas Lukosevicius, Deimante Tilinde, Violeta Salteniene, Lina Poskiene, Skaidrius Miliauskas, Jurgita Skieceviciene, Marius Zemaitis, and Juozas Kupcinskas. 2024. "Detailed Characterization of the Lung–Gut Microbiome Axis Reveals the Link between PD-L1 and the Microbiome in Non-Small-Cell Lung Cancer Patients" International Journal of Molecular Sciences 25, no. 4: 2323. https://doi.org/10.3390/ijms25042323
APA StyleAnkudavicius, V., Nikitina, D., Lukosevicius, R., Tilinde, D., Salteniene, V., Poskiene, L., Miliauskas, S., Skieceviciene, J., Zemaitis, M., & Kupcinskas, J. (2024). Detailed Characterization of the Lung–Gut Microbiome Axis Reveals the Link between PD-L1 and the Microbiome in Non-Small-Cell Lung Cancer Patients. International Journal of Molecular Sciences, 25(4), 2323. https://doi.org/10.3390/ijms25042323