
Polyallylamine Binds to Aβ Amyloid and Inhibits Antibody Recognition


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
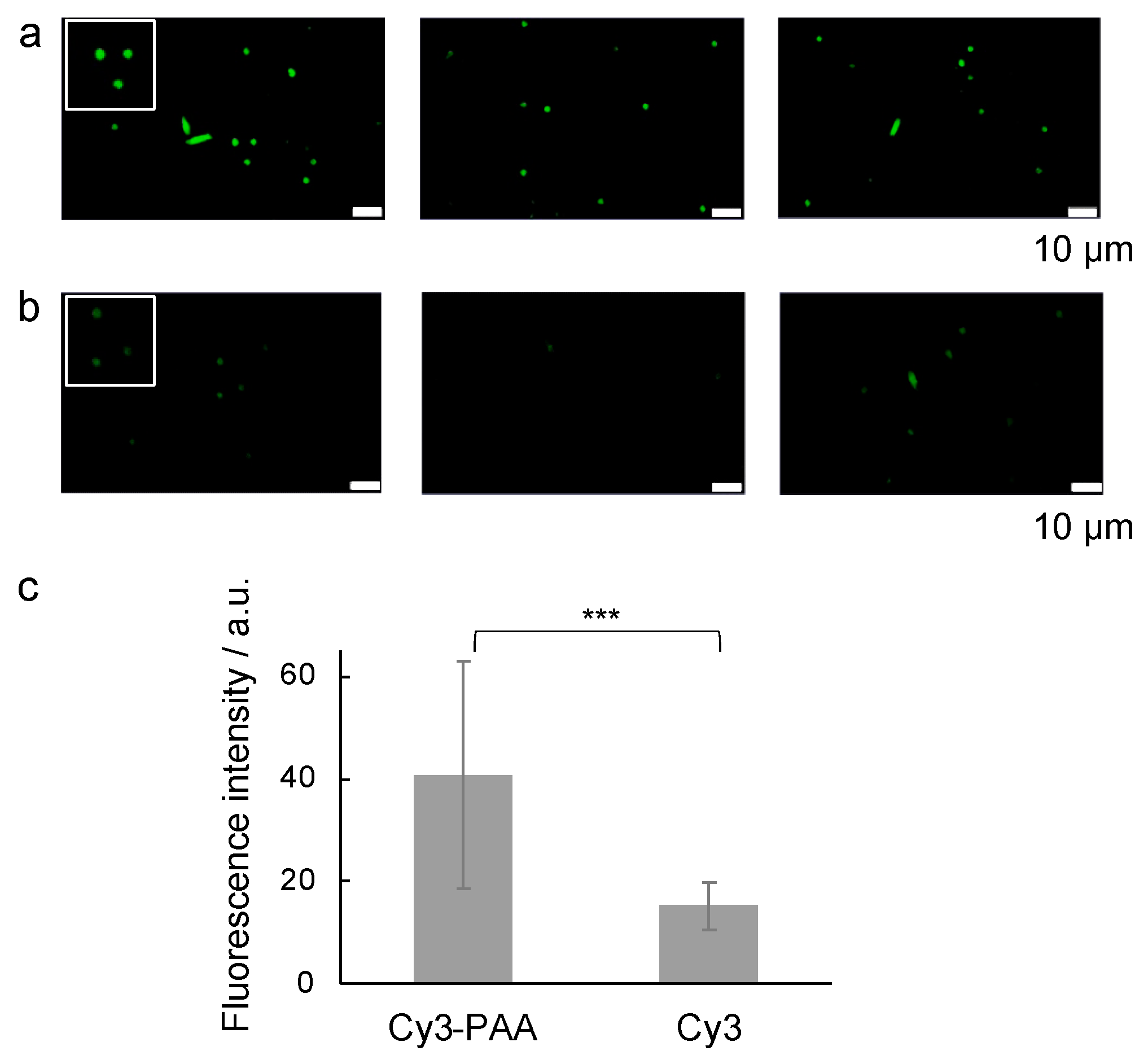
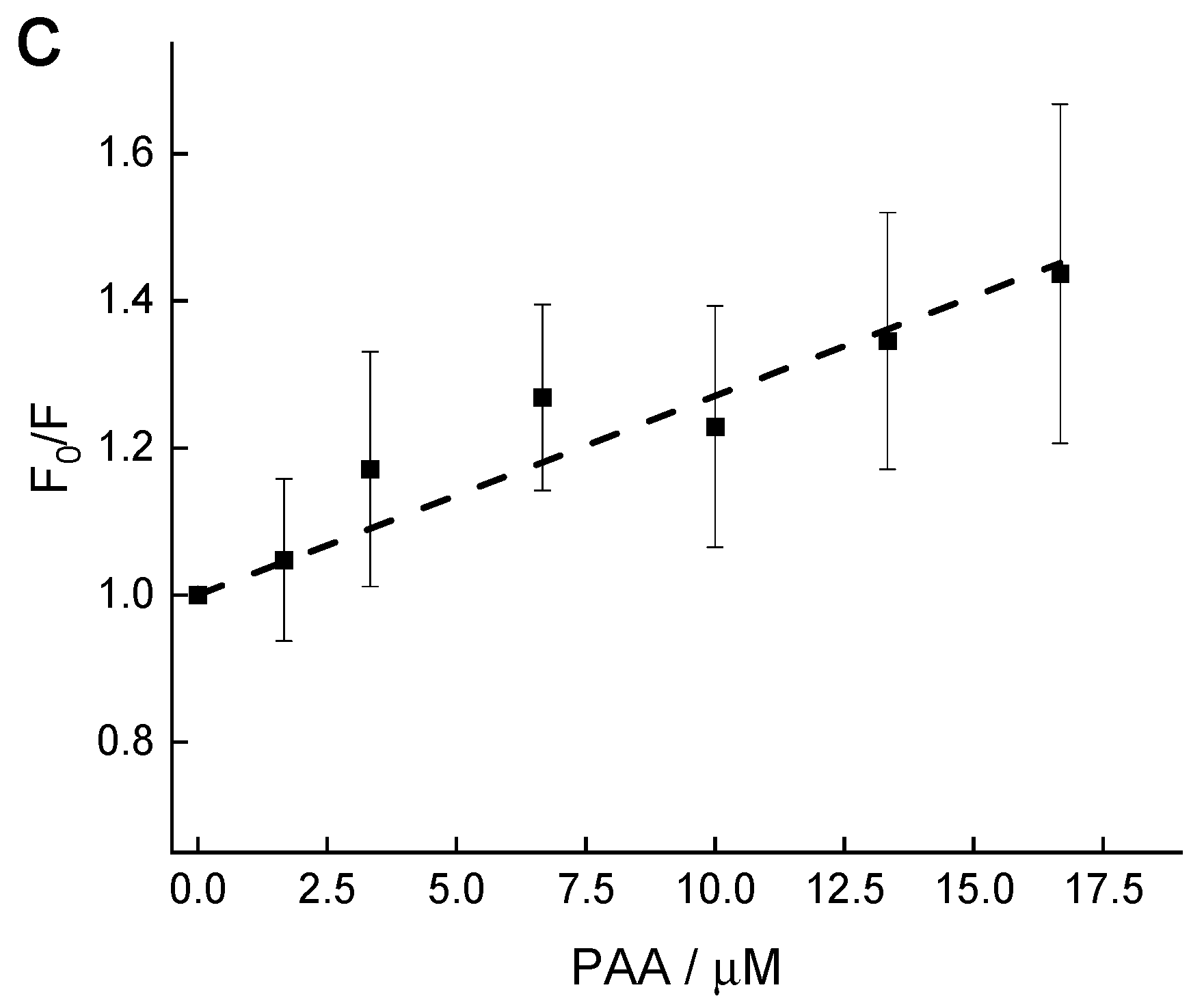
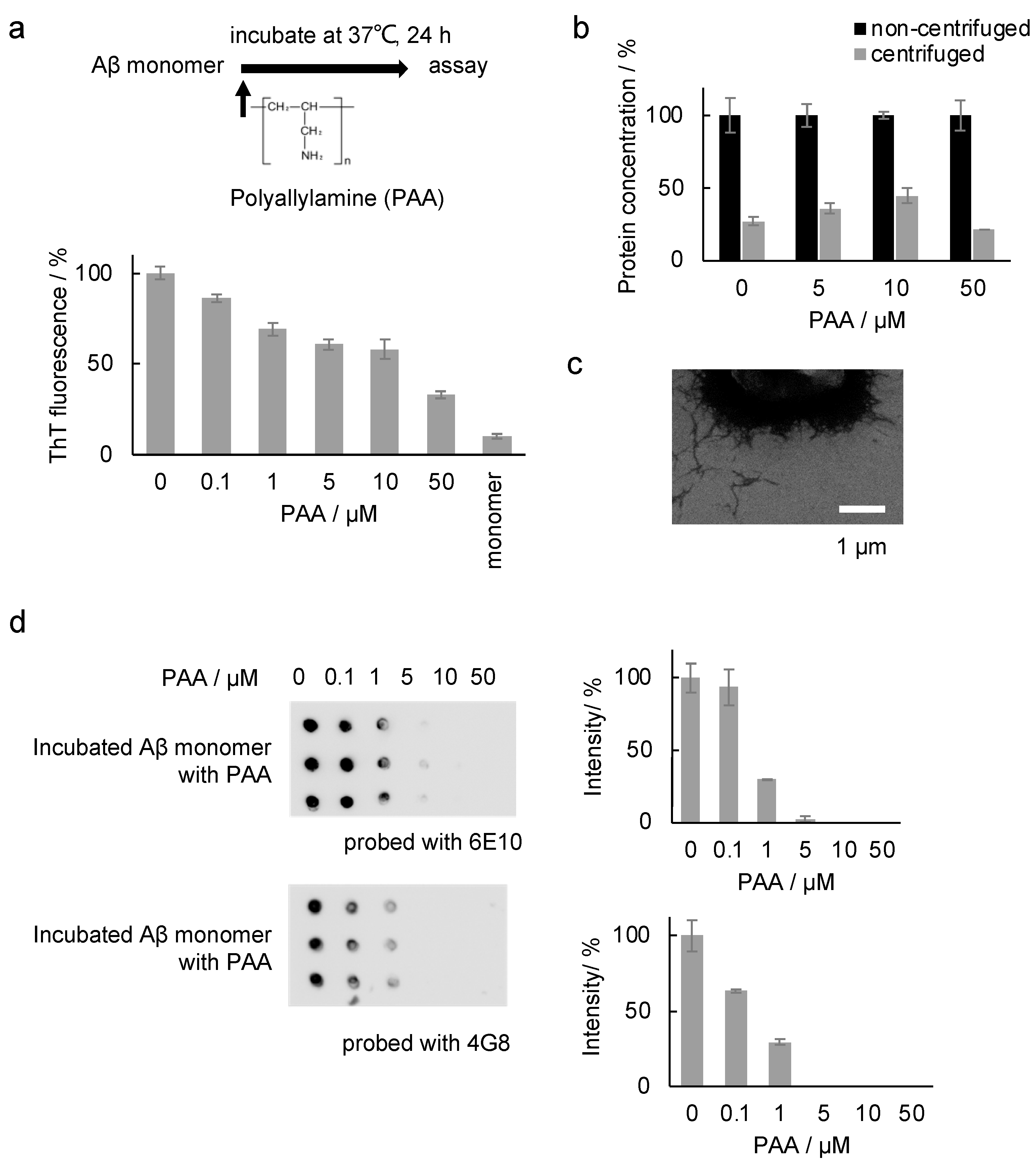
2.1. Effect of PAA on the Aβ Amyloid
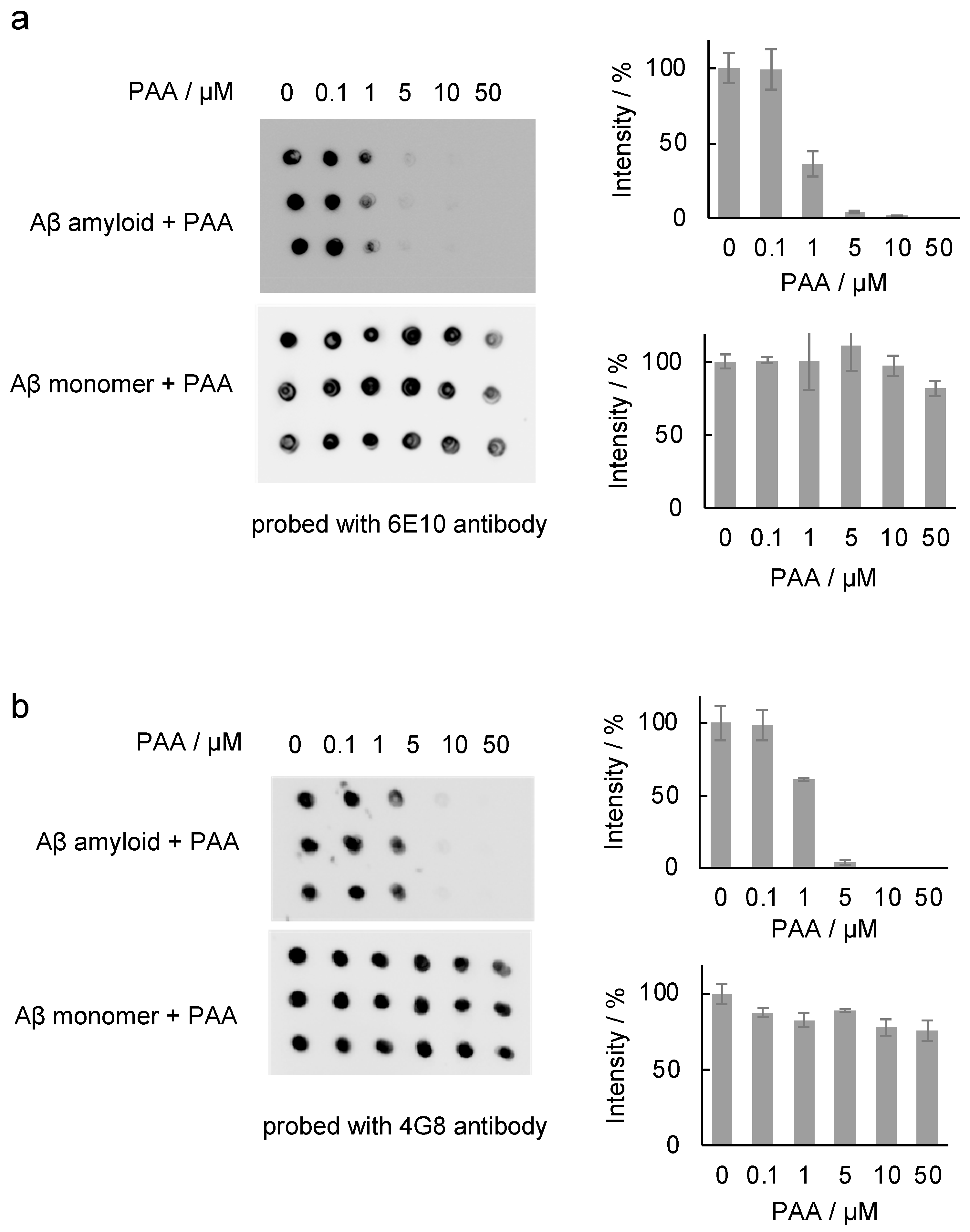
2.2. Effect of PAA on the Binding of Antibody to the Aβ42 Amyloid
2.3. Effect of PAA during the Incubation of the Aβ Sample
3. Materials and Methods
3.1. Materials
3.2. Formation of Aβ42 Amyloid and Effect of PAA
3.3. ThT Assay
3.4. Fluorescence Microscopy
3.5. Tyrosine Fluorescence
3.6. Scanning Electron Microscope (SEM)
3.7. Dot Blot Assay
3.8. Cytotoxicity Assay
3.9. Measurement of the Protein Concentration in the Supernatant
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ross, C.A.; Poirier, M.A. Protein Aggregation and Neurodegenerative Disease. Nat. Med. 2004, 10, S10. [Google Scholar] [CrossRef] [PubMed]
- Stefani, M.; Dobson, C.M. Protein Aggregation and Aggregate Toxicity: New Insights into Protein Folding, Misfolding Diseases and Biological Evolution. J. Mol. Med. 2003, 81, 678–699. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.L.; Guijarro, J.I.; Orlova, E.; Zurdo, J.; Dobson, C.M.; Sunde, M.; Saibil, H.R. Cryo-Electron Microscopy Structure of an SH3 Amyloid Fibril and Model of the Molecular Packing. EMBO J. 1999, 18, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Biancalana, M.; Koide, S. Molecular Mechanism of Thioflavin-T Binding to Amyloid Fibrils. Biochim. Biophys. Acta Proteins Proteom. 2010, 1804, 1405–1412. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth Pryor, N.; Moss, M.A.; Hestekin, C.N. Unraveling the Early Events of Amyloid-ß Protein (Aß) Aggregation: Techniques for the Determination of Aβ Aggregate Size. Int. J. Mol. Sci. 2012, 13, 3038–3072. [Google Scholar] [CrossRef] [PubMed]
- Kayed, R.; Head, E.; Thompson, J.L.; McIntire, T.M.; Milton, S.C.; Cotman, C.W.; Glabel, C.G. Common Structure of Soluble Amyloid Oligomers Implies Common Mechanism of Pathogenesis. Science 2003, 300, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Sörgjerd, K.M.; Zako, T.; Sakono, M.; Stirling, P.C.; Leroux, M.R.; Saito, T.; Nilsson, P.; Sekimoto, M.; Saido, T.C.; Maeda, M. Human Prefoldin Inhibits Amyloid-β (Aβ) Fibrillation and Contributes to Formation of Nontoxic Aβ Aggregates. Biochemistry 2013, 52, 3532–3542. [Google Scholar] [CrossRef]
- Otzen, D.; Riek, R. Functional Amyloids. Cold Spring Harb. Perspect. Biol. 2019, 11, a033860. [Google Scholar] [CrossRef]
- Knowles, T.P.J.; Mezzenga, R. Amyloid Fibrils as Building Blocks for Natural and Artificial Functional Materials. Adv. Mater. 2016, 28, 6546–6561. [Google Scholar] [CrossRef]
- Wei, G.; Su, Z.; Reynolds, N.P.; Arosio, P.; Hamley, I.W.; Gazit, E.; Mezzenga, R. Self-Assembling Peptide and Protein Amyloids: From Structure to Tailored Function in Nanotechnology. Chem. Soc. Rev. 2017, 46, 4661–4708. [Google Scholar] [CrossRef]
- Maji, S.K.; Perrin, M.H.; Sawaya, M.R.; Jessberger, S.; Vadodaria, K.; Rissman, R.A.; Singru, P.S.; Nilsson, K.P.R.; Simon, R.; Schubert, D.; et al. Functional Amyloids as Natural Storage of Peptide Hormones in Pituitary Secretory Granules. Science 2009, 325, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Baxa, U.; Cheng, N.; Winkler, D.C.; Chiu, T.K.; Davies, D.R.; Sharma, D.; Inouye, H.; Kirschner, D.A.; Wickner, R.B.; Steven, A.C. Filaments of the Ure2p Prion Protein Have a Cross-β Core Structure. J. Struct. Biol. 2005, 150, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Tursi, S.A.; Tükel, Ç. Curli-Containing Enteric Biofilms inside and out: Matrix Composition, Immune Recognition, and Disease Implications. Microbiol. Mol. Biol. Rev. 2018, 82, e00028-18. [Google Scholar] [CrossRef] [PubMed]
- Gras, S.L.; Tickler, A.K.; Squires, A.M.; Devlin, G.L.; Horton, M.A.; Dobson, C.M.; MacPhee, C.E. Functionalised Amyloid Fibrils for Roles in Cell Adhesion. Biomaterials 2008, 29, 1553–1562. [Google Scholar] [CrossRef] [PubMed]
- Sakono, M.; Akiyama, S.; Zako, T.; Sakaki, S.; Waku, T.; Tanaka, N.; Maeda, M. Immobilized Insulin Amyloid Enhances Cell Adhesion and Proliferation Due to Interaction with Fibronectin. Chem. Lett. 2011, 40, 315–317. [Google Scholar] [CrossRef]
- Sakono, M.; Akiyama, S.; Zako, T.; Sakaki, S.; Waku, T.; Tanaka, N.; Maeda, M. Application of Two Morphologically Different Fibrillar and Filamentous Insulin Amyloids as a Biomaterial for Cell Culture Surfaces. React. Funct. Polym. 2011, 71, 324–328. [Google Scholar] [CrossRef]
- Bolisetty, S.; Adamcik, J.; Heier, J.; Mezzenga, R. Amyloid Directed Synthesis of Titanium Dioxide Nanowires and Their Applications in Hybrid Photovoltaic Devices. Adv. Funct. Mater. 2012, 22, 3424–3428. [Google Scholar] [CrossRef]
- Osaka, T.; Matsunaga, T.; Nakanishi, T.; Arakaki, A.; Niwa, D.; Iida, H. Synthesis of Magnetic Nanoparticles and Their Application to Bioassays. Anal. Bioanal. Chem. 2006, 384, 593–600. [Google Scholar] [CrossRef]
- Oberhaus, F.V.; Frense, D.; Beckmann, D. Immobilization Techniques for Aptamers on Gold Electrodes for the Electrochemical Detection of Proteins: A Review. Biosensors 2020, 10, 45. [Google Scholar] [CrossRef]
- Gazit, E. Use of Biomolecular Templates for the Fabrication of Metal Nanowires. FEBS J. 2007, 274, 317–322. [Google Scholar] [CrossRef]
- Wicklein, B.; Salazar-Alvarez, G. Functional Hybrids Based on Biogenic Nanofibrils and Inorganic Nanomaterials. J. Mater. Chem. A 2013, 1, 5469–5478. [Google Scholar] [CrossRef]
- Diáz-Caballero, M.; Navarro, S.; Ventura, S. Functionalized Prion-Inspired Amyloids for Biosensor Applications. Biomacromolecules 2021, 22, 2822–2833. [Google Scholar] [CrossRef]
- Arad, E.; Jelinek, R. Catalytic Amyloids. Trends Chem. 2022, 4, 907–917. [Google Scholar] [CrossRef]
- Boussif, O.; Delair, T.; Brua, C.; Veron, L.; Pavirani, A.; Kolbe, H.V.J. Synthesis of Polyallylamine Derivatives and Their Use as Gene Transfer Vectors in Vitro. Bioconjug. Chem. 1999, 10, 877–883. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Huang, J.; Jiang, H.L.; Quan, J.S.; Cho, M.H.; Cho, C.S. Guanidinylated Poly (Allyl Amine) as a Gene Carrier. J. Appl. Polym. Sci. 2009, 112, 926–933. [Google Scholar] [CrossRef]
- Thompson, C.J.; Tetley, L.; Cheng, W.P. The Influence of Polymer Architecture on the Protective Effect of Novel Comb Shaped Amphiphilic Poly(Allylamine) against in vitro Enzymatic Degradation of Insulin-Towards Oral Insulin Delivery. Int. J. Pharm. 2010, 383, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Lakowicz, J.R. Energy Transfer. In Principles of Fluorescence Spectroscopy; Springer US: Boston, MA, USA, 1999; pp. 367–394. ISBN 978-1-4757-3061-6. [Google Scholar]
- Zhang, Y.; Yang, X.; Liu, L.; Huang, Z.; Pu, J.; Gaobo, L.; Zhang, L.; Liu, D.; Xu, B.; Liao, J.; et al. Comparison of Forster-Resonance-Energy-Transfer Acceptors for Tryptophan and Tyrosine Residues in Native Proteins as Donors. J. Fluoresc. 2013, 23, 147–157. [Google Scholar] [CrossRef]
- Fotouhi, L.; Tabatabaee, R. A Study of the Interaction Tyrosine and DNA Using Voltammetry and Spectroscopy Methods. Spectrochim. Acta Part. A Mol. Biomol. Spectrosc. 2014, 121, 152–156. [Google Scholar] [CrossRef]
- Qiu, L.; Lewis, A.; Como, J.; Vaughn, M.W.; Huang, J.; Somerharju, P.; Virtanen, J.; Kwan, H.C. Cholesterol Modulates the Interaction of β-Amyloid Peptide with Lipid Bilayers. Biophys. J. 2009, 96, 4299–4307. [Google Scholar] [CrossRef]
- Lührs, T.; Ritter, C.; Adrian, M.; Riek-Loher, D.; Bohrmann, B.; Döbeli, H.; Schubert, D.; Riek, R. 3D Structure of Alzheimer’s Amyloid-β(1-42) Fibrils. Proc. Natl. Acad. Sci. USA 2005, 102, 17342–17347. [Google Scholar] [CrossRef]
- Ballet, T.; Brukert, F.; Mangiagalli, P.; Bureau, C.; Boulangé, L.; Nault, L.; Perret, T.; Weidenhaupt, M. DnaK Prevents Human Insulin Amyloid Fiber Formation on Hydrophobic Surfaces. Biochemistry 2012, 51, 2172–2180. [Google Scholar] [CrossRef] [PubMed]
- Moores, B.; Drolle, E.; Attwood, S.J.; Simons, J.; Leonenko, Z. Effect of Surfaces on Amyloid Fibril Formation. PLoS ONE 2011, 6, e25954. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, H.; Ikawa, S.; Kitano, K.; Zako, T. Peroxynitric Acid Inhibits Amyloid β Aggregation. Biochem. Biophys. Res. Commun. 2023, 660, 1–5. [Google Scholar] [CrossRef]
- Kunitomi, R.; Pradipta, A.R.; Kawabe, H.; Lobsiger, N.; Tanaka, K.; Zako, T. Inhibition of Amyloid Formation of Amyloid β (1–42), Amylin and Insulin by 1,5-Diazacyclooctanes, a Spermine-Acrolein Conjugate. Bioorganic Med. Chem. 2021, 46, 116391. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.; Zako, T.; Zeltner, M.; Sörgjerd, K.M.; Schumacher, C.M.; Hofer, C.J.; Stark, W.J.; Maeda, M. Adsorption and Separation of Amyloid Beta Aggregates Using Ferromagnetic Nanoparticles Coated with Charged Polymer Brushes. J. Mater. Chem. B 2015, 3, 3351–3357. [Google Scholar] [CrossRef]
- Kollmer, M.; Close, W.; Funk, L.; Rasmussen, J.; Bsoul, A.; Schierhorn, A.; Schmidt, M.; Sigurdson, C.J.; Jucker, M.; Fändrich, M. Cryo-EM Structure and Polymorphism of Aβ Amyloid Fibrils Purified from Alzheimer’s Brain Tissue. Nat. Commun. 2019, 10, 4760. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuchie, Y.; Kusuda, S.; Kawabe, H.; Mori, W.; Lindgren, M.; Watanabe, Y.; Zako, T. Polyallylamine Binds to Aβ Amyloid and Inhibits Antibody Recognition. Int. J. Mol. Sci. 2024, 25, 3112. https://doi.org/10.3390/ijms25063112
Tsuchie Y, Kusuda S, Kawabe H, Mori W, Lindgren M, Watanabe Y, Zako T. Polyallylamine Binds to Aβ Amyloid and Inhibits Antibody Recognition. International Journal of Molecular Sciences. 2024; 25(6):3112. https://doi.org/10.3390/ijms25063112
Chicago/Turabian StyleTsuchie, Yusuke, Soichiro Kusuda, Haruka Kawabe, Wakako Mori, Mikael Lindgren, Yutaka Watanabe, and Tamotsu Zako. 2024. "Polyallylamine Binds to Aβ Amyloid and Inhibits Antibody Recognition" International Journal of Molecular Sciences 25, no. 6: 3112. https://doi.org/10.3390/ijms25063112
APA StyleTsuchie, Y., Kusuda, S., Kawabe, H., Mori, W., Lindgren, M., Watanabe, Y., & Zako, T. (2024). Polyallylamine Binds to Aβ Amyloid and Inhibits Antibody Recognition. International Journal of Molecular Sciences, 25(6), 3112. https://doi.org/10.3390/ijms25063112