
Advances in Gluten Hypersensitivity: Novel Dietary-Based Therapeutics in Research and Development

 ,
,
Abstract
:1. Introduction
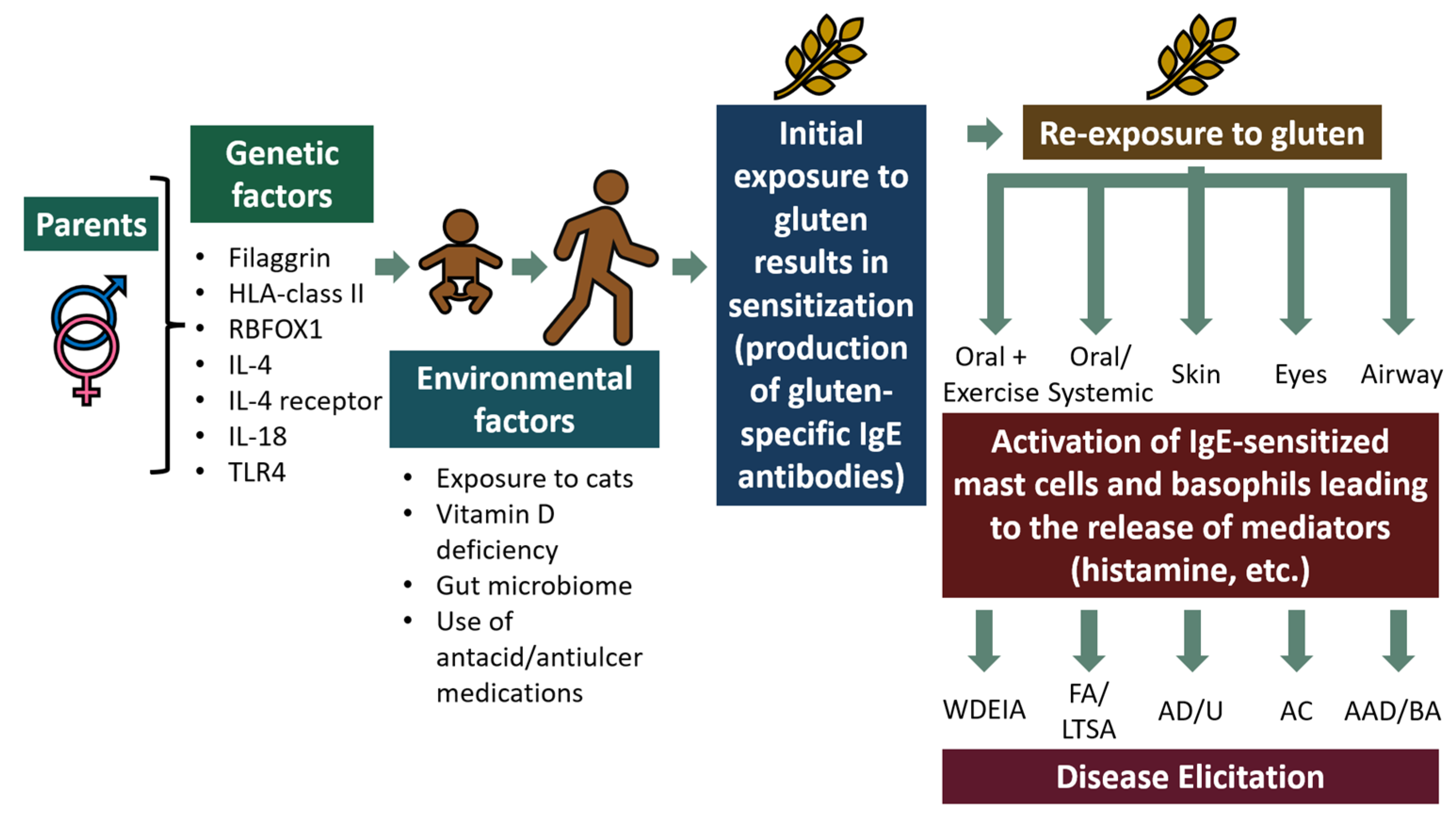
2. Mechanism of Gluten Hypersensitivity
3. Preclinical Development of Novel Dietary-Based Therapeutics Using Animal Models of Gluten Hypersensitivity
3.1. Animal Testing
3.1.1. Canine Model: Potential of Thioredoxin and Heat-Killed Listeria monocytogenes to Reduce Gluten Allergenicity
3.1.2. Evidence from Rat Models: Potential of Gluten Genetic Deletion, Deamidation, and Enzymatic Digestion to Reduce Gluten Allergenicity
3.1.3. Guinea Pig Model: Potential of Gluten Genetic Deletion to Reduce Gluten Allergenicity
3.1.4. Mouse Models: Potential of Probiotics, Enzymatic Digestion, L-arabinose, Deamidation, Polyphenols from Fruits, and Phosphorylation of Gluten to Reduce Wheat Allergenicity
4. Other Approaches to Produce Hypoallergenic Gluten Products
5. Effects of Fermentation, Gluten Gene Targeting, Deamidation, Thioredoxin, and Enzyme Treatment on Gluten Allergenicity in Humans
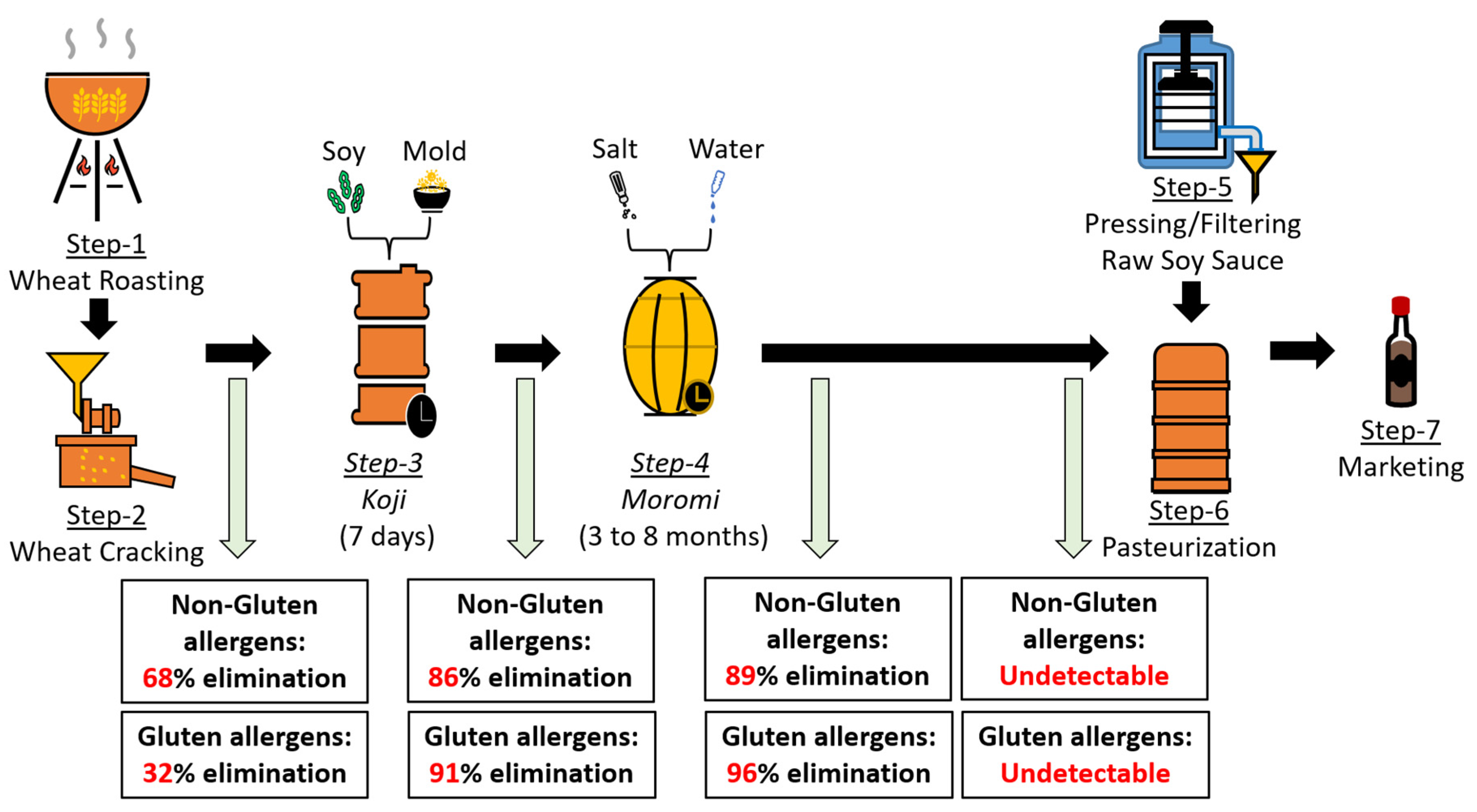
5.1. Soy Sauce Fermentation Can Reduce/Eliminate Human Gluten Allergenicity: In Vitro Evidence
5.2. Gluten Gene Targeting Can Be Used to Develop Hypoallergenic Wheat Lines: In Vitro Evidence from Human Studies
5.3. Gluten Gene Targeting and Enzyme Hydrolysis to Develop Hypoallergenic Wheat Products: Clinical Evidence from Testing in Gluten Hypersensitive Subjects
5.4. Optimized Thermal Processing Methods May Be Used to Produce Potentially Hypoallergenic Gluten Products
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- OECD; FAO. OECD-FAO Agricultural Outlook 2021–2030; FAO: Rome, Italy; OECD Publishing: Paris, France, 2021. [Google Scholar]
- Sievers, S.; Rohrbach, A.; Beyer, K. Wheat-Induced Food Allergy in Childhood: Ancient Grains Seem No Way Out. Eur. J. Nutr. 2020, 59, 2693–2707. [Google Scholar] [CrossRef] [PubMed]
- Pasha, I.; Saeed, F.; Sultan, M.T.; Batool, R.; Aziz, M.; Ahmed, W. Wheat Allergy and Intolerence; Recent Updates and Perspectives. Crit. Rev. Food Sci. Nutr. 2016, 56, 13–24. [Google Scholar] [CrossRef] [PubMed]
- García-Manzanares, Á.; Lucendo, A.J. Nutritional and Dietary Aspects of Celiac Disease. Nutr. Clin. Pract. 2011, 26, 163–173. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, T.; Ribeiro, M.; Sabença, C.; Igrejas, G. The 10,000-Year Success Story of Wheat! Foods 2021, 10, 2124. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, B. Gluten-Related Disorders: Celiac Disease, Wheat Allergy, and Nonceliac Gluten Sensitivity. Crit. Rev. Food Sci. Nutr. 2020, 60, 2606–2621. [Google Scholar] [CrossRef] [PubMed]
- Ricci, G.; Andreozzi, L.; Cipriani, F.; Giannetti, A.; Gallucci, M.; Caffarelli, C. Wheat Allergy in Children: A Comprehensive Update. Medicina 2019, 55, 400. [Google Scholar] [CrossRef] [PubMed]
- Pastorello, E.A.; Farioli, L.; Conti, A.; Pravettoni, V.; Bonomi, S.; Iametti, S.; Fortunato, D.; Scibilia, J.; Bindslev-Jensen, C.; Ballmer-Weber, B.; et al. Wheat IgE-Mediated Food Allergy in European Patients: α-Amylase Inhibitors, Lipid Transfer Proteins and Low-Molecular-Weight Glutenins—Allergenic Molecules Recognized by Double-Blind, Placebo-Controlled Food Challenge. Int. Arch. Allergy Immunol. 2007, 144, 10–22. [Google Scholar] [CrossRef]
- Shewry, P.R.; Napier, J.A.; Tatham, A.S. Seed Storage Proteins: Structures and Biosynthesis. Plant Cell 1995, 7, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H. Chemistry of Gluten Proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Yamada, T.; Maruyama, N. Wheat α-Gliadin and High-Molecular-Weight Glutenin Subunit Accumulate in Different Storage Compartments of Transgenic Soybean Seed. Transgenic Res. 2022, 31, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Tosi, P.; D’Ovidio, R.; Napier, J.A.; Bekes, F.; Shewry, P.R. Expression of Epitope-Tagged LMW Glutenin Subunits in the Starchy Endosperm of Transgenic Wheat and Their Incorporation into Glutenin Polymers. Theor. Appl. Genet. 2004, 108, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Dong, Y.; An, X.; Wang, A.; Zhang, Y.; Li, X.; Gao, L.; Xia, X.; He, Z.; Yan, Y. Characterization of HMW Glutenin Subunits in Common Wheat and Related Species by Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF-MS). J. Cereal Sci. 2008, 47, 252–261. [Google Scholar] [CrossRef]
- Liu, L.; Wang, A.; Appels, R.; Ma, J.; Xia, X.; Lan, P.; He, Z.; Bekes, F.; Yan, Y.; Ma, W. A MALDI-TOF Based Analysis of High Molecular Weight Glutenin Subunits for Wheat Breeding. J. Cereal Sci. 2009, 50, 295–301. [Google Scholar] [CrossRef]
- Peng, Y.; Yu, K.; Zhang, Y.; Islam, S.; Sun, D.; Ma, W. Two Novel Y-Type High Molecular Weight Glutenin Genes in Chinese Wheat Landraces of the Yangtze-River Region. PLoS ONE 2015, 10, e0142348. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fu, J.; Shen, Q.; Yang, D. High-Molecular-Weight Glutenin Subunits: Genetics, Structures, and Relation to End Use Qualities. Int. J. Mol. Sci. 2021, 22, 184. [Google Scholar] [CrossRef] [PubMed]
- Scherf, K.A.; Brockow, K.; Biedermann, T.; Koehler, P.; Wieser, H. Wheat-Dependent Exercise-Induced Anaphylaxis. Clin. Exp. Allergy 2016, 46, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Kotaniemi-Syrjänen, A.; Palosuo, K.; Jartti, T.; Kuitunen, M.; Pelkonen, A.S.; Mäkelä, M.J. The Prognosis of Wheat Hypersensitivity in Children. Pediatr. Allergy Immunol. 2010, 21, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Baar, A.; Pahr, S.; Constantin, C.; Scheiblhofer, S.; Thalhamer, J.; Giavi, S.; Papadopoulos, N.G.; Ebner, C.; Mari, A.; Vrtala, S.; et al. Molecular and Immunological Characterization of Tri a 36, a Low Molecular Weight Glutenin, as a Novel Major Wheat Food Allergen. J. Immunol. 2012, 189, 3018–3025. [Google Scholar] [CrossRef] [PubMed]
- Cianferoni, A. Wheat Allergy: Diagnosis and Management. J. Asthma Allergy 2016, 9, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Lebwohl, B.; Sanders, D.S.; Green, P.H.R. Coeliac Disease. Lancet 2018, 391, 70–81. [Google Scholar] [CrossRef]
- Cárdenas-Torres, F.I.; Cabrera-Chávez, F.; Figueroa-Salcido, O.G.; Ontiveros, N. Non-Celiac Gluten Sensitivity: An Update. Medicina 2021, 57, 526. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Sapone, A.; Zevallos, V.; Schuppan, D. Nonceliac Gluten Sensitivity. Gastroenterology 2015, 148, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Jorgensen, R.; Raghunath, R.; Nagisetty, S.; Ng, P.K.W.; Gangur, V. Creating Hypo-/Nonallergenic Wheat Products Using Processing Methods: Fact or Fiction? Compr. Rev. Food Sci. Food Saf. 2021, 20, 6089–6115. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Acharya, H.G.; Acharya, D.; Jorgensen, R.; Gao, H.; Secord, J.; Ng, P.K.W.; Gangur, V. Advances in Molecular Mechanisms of Wheat Allergenicity in Animal Models: A Comprehensive Review. Molecules 2019, 24, 1142. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food Allergy: A Review and Update on Epidemiology, Pathogenesis, Diagnosis, Prevention, and Management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Iga, N.; Tanizaki, H.; Endo, Y.; Egawa, G.; Fujisawa, A.; Tanioka, M.; Miyachi, Y.; Kabashima, K. Hydrolyzed Wheat Protein-Containing Facial Soap-Induced Wheat-Dependent Exercise-Induced Anaphylaxis in a Patient without Filaggrin Mutations. J. Dermatol. 2013, 40, 494–495. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, O.; Nomura, T.; Ohguchi, Y.; Suzuki, S.; Nomura, Y.; Hamade, Y.; Hoshina, D.; Sandilands, A.; Akiyama, M.; McLean, W.H.I.; et al. Loss-of-Function Mutations in the Gene Encoding Filaggrin Underlie a Japanese Family with Food-Dependent Exercise-Induced Anaphylaxis. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, K.; Chinuki, Y.; Hamada, Y.; Fukutomi, Y.; Sugiyama, A.; Kishikawa, R.; Fukunaga, A.; Oda, Y.; Ugajin, T.; Yokozeki, H.; et al. Genome-Wide Association Study Reveals an Association between the HLA-DPB1∗02:01:02 Allele and Wheat-Dependent Exercise-Induced Anaphylaxis. Am. J. Hum. Genet. 2021, 108, 1540–1548. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, E.; Akiyama, M.; Yagami, A.; Hirota, T.; Okada, Y.; Kato, Z.; Kishikawa, R.; Fukutomi, Y.; Hide, M.; Morita, E.; et al. HLA-DQ and RBFOX1 as Susceptibility Genes for an Outbreak of Hydrolyzed Wheat Allergy. J. Allergy Clin. Immunol. 2019, 144, 1354–1363. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.P.; Yin, J. Association between Single Nucleotide Polymorphisms and Wheat-Dependent Exercise-Induced Anaphylaxis in Chinese Population. Chin. Med. J. 2013, 126, 1159–1165. [Google Scholar] [CrossRef] [PubMed]
- Hur, G.Y.; Ye, Y.M.; Koh, D.H.; Kim, S.H.; Park, H.S. IL-4 Receptor a Polymorphisms May Be a Susceptible Factor for Work-Related Respiratory Symptoms in Bakery Workers. Allergy Asthma Immunol. Res. 2013, 5, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wen, L.; Li, H.; Wang, R.; Yin, J. Genetic Variation at the Interleukin-18 Locus Is Associated with Wheat-Dependent Exercise-Induced Anaphylaxis in the Han Chinese Population. Gene 2020, 737, 144462. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Kim, S.H.; Kim, J.H.; Choi, H.; Son, J.K.; Hur, G.Y.; Park, H.S. Effect of Toll-like Receptor 4 Gene Polymorphisms on Work-Related Respiratory Symptoms and Sensitization to Wheat Flour in Bakery Workers. Ann. Allergy Asthma Immunol. 2011, 107, 57–64. [Google Scholar] [CrossRef] [PubMed]
- von Mutius, E. Allergies, Infections and the Hygiene Hypothesis—The Epidemiological Evidence. Immunobiology 2007, 212, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Okabe, H.; Hashimoto, K.; Yamada, M.; Ono, T.; Yaginuma, K.; Kume, Y.; Chishiki, M.; Sato, A.; Ogata, Y.; Imaizumi, K.; et al. Associations between Fetal or Infancy Pet Exposure and Food Allergies: The Japan Environment and Children’s Study. PLoS ONE 2023, 18, e0282725. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Shin, Y.H.; Chung, I.H.; Kim, H.J.; Yoo, E.G.; Yoon, J.W.; Jee, H.M.; Chang, Y.E.; Han, M.Y. The Link between Serum Vitamin d Level, Sensitization to Food Allergens, and the Severity of Atopic Dermatitis in Infancy. J. Pediatr. 2014, 165, 849–854.e1. [Google Scholar] [CrossRef] [PubMed]
- Savage, J.H.; Lee-Sarwar, K.A.; Sordillo, J.; Bunyavanich, S.; Zhou, Y.; O’Connor, G.; Sandel, M.; Bacharier, L.B.; Zeiger, R.; Sodergren, E.; et al. A Prospective Microbiome-Wide Association Study of Food Sensitization and Food Allergy in Early Childhood. Allergy Eur. J. Allergy Clin. Immunol. 2018, 73, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Untersmayr, E.; Bakos, N.; Schöll, I.; Kundi, M.; Roth-Walter, F.; Szalai, K.; Riemer, A.B.; Ankersmit, H.J.; Scheiner, O.; Boltz-Nitulescu, G.; et al. Anti-ulcer Drugs Promote IgE Formation toward Dietary Antigens in Adult Patients. FASEB J. 2005, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- De Zorzi, M.; Curioni, A.; Simonato, B.; Giannattasio, M.; Pasini, G. Effect of Pasta Drying Temperature on Gastrointestinal Digestibility and Allergenicity of Durum Wheat Proteins. Food Chem. 2007, 104, 353–363. [Google Scholar] [CrossRef]
- Pawankar, R.; Hayashi, M.; Yamanishi, S.; Igarashi, T. The Paradigm of Cytokine Networks in Allergic Airway Inflammation. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Finkelman, F.D.; Rothenberg, M.E.; Brandt, E.B.; Morris, S.C.; Strait, R.T. Molecular Mechanisms of Anaphylaxis: Lessons from Studies with Murine Models. J. Allergy Clin. Immunol. 2005, 115, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Leung, D.Y.M. Advances in Allergic Skin Disease, Anaphylaxis, and Hypersensitivity Reactions to Foods, Drugs, and Insects in 2014. J. Allergy Clin. Immunol. 2015, 135, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Stone, K.D.; Prussin, C.; Metcalfe, D.D. IgE, Mast Cells, Basophils, and Eosinophils. J. Allergy Clin. Immunol. 2010, 125, S73–S80. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B.; Adamidi, C.; Lozano, R.M.; Yee, B.C.; Momma, M.; Kobrehel, K.; Ermel, R.; Frick, O.L. Thioredoxin-Linked Mitigation of Allergic Responses to Wheat. Proc. Natl. Acad. Sci. USA 1997, 94, 5372–5377. [Google Scholar] [CrossRef] [PubMed]
- Frick, O.L.; Teuber, S.S.; Buchanan, B.B.; Morigasaki, S.; Umetsu, D.T. Allergen Immunotherapy with Heat-Killed Listeria monocytogenes Alleviates Peanut and Food-Induced Anaphylaxis in Dogs. Allergy Eur. J. Allergy Clin. Immunol. 2005, 60, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yokooji, T.; Ninomiya, N.; Taogoshi, T.; Morita, E.; Matsuo, H. Evaluation of the Allergenicity of Ω5-Gliadin-Deficient Hokushin Wheat (1BS-18) in a Wheat Allergy Rat Model. Biochem. Biophys. Rep. 2019, 20, 100702. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Yokooji, T.; Kunimoto, K.; Inoguchi, K.; Ogino, R.; Taogoshi, T.; Morita, E.; Matsuo, H. Hypoallergenic Wheat Line (1BS-18H) Lacking Ω5-Gliadin Induces Oral Tolerance to Wheat Gluten Proteins in a Rat Model of Wheat Allergy. Foods 2022, 11, 2181. [Google Scholar] [CrossRef]
- Kumagai, H.; Suda, A.; Sakurai, H.; Kumagai, H.; Arai, S.; Inomata, N.; Ikezawa, Z. Improvement of Digestibility, Reduction in Allergenicity, and Induction of Oral Tolerance of Wheat Gliadin by Deamidation. Biosci. Biotechnol. Biochem. 2007, 71, 977–985. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, J.; Tanabe, S.; Watanabe, M.; Kasai, T.; Sonoyama, K. Consumption of Hypoallergenic Flour Prevents Gluten-Induced Airway Inflammation in Brown Norway Rats. Biosci. Biotechnol. Biochem. 2001, 65, 1729–1735. [Google Scholar] [CrossRef]
- Kohno, K.; Takahashi, H.; Endo, T.R.; Matsuo, H.; Shiwaku, K.; Morita, E. Characterization of a Hypoallergenic Wheat Line Lacking ω-5 Gliadin. Allergol. Int. 2016, 65, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Chen, C.; Xie, Q.; Gu, S.; Tao, S.; Xue, W. Pediococcus Acidilactici Strain Alleviates Gluten-Induced Food Allergy and Regulates Gut Microbiota in Mice. Front. Cell. Infect. Microbiol. 2022, 12, 845142. [Google Scholar] [CrossRef] [PubMed]
- Mohan Kumar, B.V.; Vijaykrishnaraj, M.; Kurrey, N.K.; Shinde, V.S.; Prabhasankar, P. Prolyl Endopeptidase-Degraded Low Immunoreactive Wheat Flour Attenuates Immune Responses in Caco-2 Intestinal Cells and Gluten-Sensitized BALB/c Mice. Food Chem. Toxicol. 2019, 129, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, J.; Xue, L.; Liu, J.; Nie, C.; Fan, M.; Qian, H.; Zhang, D.; Ying, H.; Li, Y.; et al. L-Arabinose Attenuates Gliadin-Induced Food Allergy via Regulation of Th1/Th2 Balance and Upregulation of Regulatory T Cells in Mice. J. Agric. Food Chem. 2021, 69, 3638–3646. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Miyakawa, T.; Takano, T.; Nakajima-Adachi, H.; Tanokura, M.; Hachimura, S. Induction of Oral Tolerance by Pepsin-Digested Gliadin Retaining T Cell Reactivity in a Mouse Model of Wheat Allergy. Int. Arch. Allergy Immunol. 2020, 181, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Li, Y.; Li, T.; Pan, H.; Liu, J.; Fan, M.; Qian, H.; Zhang, H.; Ying, H.; Wang, L. Phosphorylation and Enzymatic Hydrolysis with Alcalase and Papain Effectively Reduce Allergic Reactions to Gliadins in Normal Mice. J. Agric. Food Chem. 2019, 67, 6313–6323. [Google Scholar] [CrossRef] [PubMed]
- Abe, R.; Shimizu, S.; Yasuda, K.; Sugai, M.; Okada, Y.; Chiba, K.; Akao, M.; Kumagai, H.; Kumagai, H. Evaluation of Reduced Allergenicity of Deamidated Gliadin in a Mouse Model of Wheat-Gliadin Allergy Using an Antibody Prepared by a Peptide Containing Three Epitopes. J. Agric. Food Chem. 2014, 62, 2845–2852. [Google Scholar] [CrossRef] [PubMed]
- Pérot, M.; Lupi, R.; Guyot, S.; Delayre-Orthez, C.; Gadonna-Widehem, P.; Thébaudin, J.Y.; Bodinier, M.; Larré, C. Polyphenol Interactions Mitigate the Immunogenicity and Allergenicity of Gliadins. J. Agric. Food Chem. 2017, 65, 6442–6451. [Google Scholar] [CrossRef] [PubMed]
- Tsurunaga, Y.; Arima, S.; Kumagai, S.; Morita, E. Low Allergenicity in Processed Wheat Flour Products Using Tannins from Agri-Food Wastes. Foods 2023, 12, 2722. [Google Scholar] [CrossRef]
- Jorgensen, R.; Gao, H.; Selvan, T.; Arasan, A.; Van Antwerp, C.; Sundar, V. Is Wheat Glutenin Extract Intrinsically Allergenic? Evaluation Using a Novel Adjuvant-Free Mouse Model of Systemic Anaphylaxis. Int. J. Mol. Sci. 2023, 24, 17247. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, R.; Gao, H.; Chandra, S.; Sundar, V.; Loy, J.; Van Antwerp, C.; Ng, P.K.W.; Gangur, V. Chronic Application of Alcohol-Soluble Gluten Extract over Undamaged Skin Causes Clinical Sensitization for Life-Threatening Anaphylaxis via Activation of Systemic Th2 Immune Responses in Mice. Front. Allergy 2023, 4, 1214051. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yunusbaev, U.; Fritz, A.; Tilley, M.; Akhunova, A.; Trick, H.; Akhunov, E. CRISPR-Based Editing of the ω- and γ-Gliadin Gene Clusters Reduces Wheat Immunoreactivity without Affecting Grain Protein Quality. Plant Biotechnol. J. 2023, 22, 892–903. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M. Immunological Functions of Soy Sauce: Hypoallergenicity and Antiallergic Activity of Soy Sauce. J. Biosci. Bioeng. 2005, 100, 144–151. [Google Scholar] [CrossRef]
- Denery-Papini, S.; Lauriére, M.; Branlard, G.; Morisset, M.; Pecquet, C.; Choudat, D.; Merlino, M.; Pineau, F.; Popineau, Y.; Boulenc, E.; et al. Influence of the Allelic Variants Encoded at the Gli-B1 Locus, Responsible for a Major Allergen of Wheat, on IgE Reactivity for Patients Suffering from Food Allergy to Wheat. J. Agric. Food Chem. 2007, 55, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Waga, J.; Skoczowski, A. Development and Characteristics of ω-Gliadin-Free Wheat Genotypes. Euphytica 2014, 195, 105–116. [Google Scholar] [CrossRef]
- Altenbach, S.B.; Tanaka, C.K.; Pineau, F.; Lupi, R.; Drouet, M.; Beaudouin, E.; Morisset, M.; Denery-Papini, S. Assessment of the Allergenic Potential of Transgenic Wheat (Triticum aestivum) with Reduced Levels of Ω5-Gliadins, the Major Sensitizing Allergen in Wheat-Dependent Exercise-Induced Anaphylaxis. J. Agric. Food Chem. 2015, 63, 9323–9332. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, S.R.; Park, J.H.; Park, K.H.; Jeong, K.Y.; Lee, J.H.; Kang, C.S.; Kim, K.H.; Park, J.W. Evaluation of Allergenicity on a ω-5 Gliadin-Deficient Cultivar in Wheat-Dependent Exercise-Induced Anaphylaxis. Allergy Asthma Immunol. Res. 2022, 14, 379–392. [Google Scholar] [CrossRef]
- Waga, J.; Zientarski, J.; Obtułowicz, K.; Bilo, B.; Stachowicz, M. Gliadin Immunoreactivity and Dough Rheological Properties of Winter Wheat Genotypes Modified by Thioredoxin. Cereal Chem. 2008, 85, 488–494. [Google Scholar] [CrossRef]
- Lombardo, C.; Bolla, M.; Chignola, R.; Senna, G.; Rossin, G.; Caruso, B.; Tomelleri, C.; Cecconi, D.; Brandolini, A.; Zoccatelli, G. Study on the Immunoreactivity of Triticum monococcum (Einkorn) Wheat in Patients with Wheat-Dependent Exercise-Induced Anaphylaxis for the Production of Hypoallergenic Foods. J. Agric. Food Chem. 2015, 63, 8299–8306. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S. Analysis of Food Allergen Structures and Development of Foods for Allergic Patients. Biosci. Biotechnol. Biochem. 2008, 72, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.R.; Jeong, D.Y.; Baik, S.H. Effects of Indigenous Yeasts on Physicochemical and Microbial Properties of Korean Soy Sauce Prepared by Low-Salt Fermentation. Food Microbiol. 2015, 51, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, J.; Gan, H.M.; Yin, W.F.; Chan, K.G. Microbial Succession and the Functional Potential during the Fermentation of Chinese Soy Sauce Brine. Front. Microbiol. 2014, 5, 112131. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Watanabe, J.; Mogi, Y. Monitoring of the Microbial Communities Involved in the Soy Sauce Manufacturing Process by PCR-Denaturing Gradient Gel Electrophoresis. Food Microbiol. 2012, 31, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, H.; Chen, Z.; Lv, Z.; Xie, Y.; Lu, F. Profiling of Dynamic Changes in the Microbial Community during the Soy Sauce Fermentation Process. Appl. Microbiol. Biotechnol. 2013, 97, 9111–9119. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.Z.; Qian, Y.L.; Ji, F.D.; Chen, J.Y.; Han, B.Z. Microbial Composition during Chinese Soy Sauce Koji-Making Based on Culture Dependent and Independent Methods. Food Microbiol. 2013, 34, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Deng, Y.; Jin, Y.; Liu, Y.; Xia, B.; Sun, Q. Dynamics of Microbial Community during the Extremely Long-term Fermentation Process of a Traditional Soy Sauce. J. Sci. Food Agric. 2017, 97, 3220–3227. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Watanabe, J.; Sonoyama, K.; Tanabe, S. Novel Method for Producing Hypoallergenic Wheat Flour by Enzymatic Fragmentation of the Constituent Allergens and Its Application to Food Processing. Biosci. Biotechnol. Biochem. 2000, 64, 2663–2667. [Google Scholar] [CrossRef] [PubMed]
- Lupi, R.; Denery-Papini, S.; Claude, M.; Tranquet, O.; Drouet, M.; Masci, S.; Larré, C. Thermal Treatment Reduces Gliadin Recognition by IgE, but a Subsequent Digestion and Epithelial Crossing Permits Recovery. Food Res. Int. 2019, 118, 22–31. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Rizzello, C.G.; Scala, E.; De Simone, C.; Farris, G.A.; Turrini, F.; Gobbetti, M. Probiotic Preparation Has the Capacity to Hydrolyze Proteins Responsible for Wheat Allergy. J. Food Prot. 2007, 70, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Gao, H.; Jorgensen, R.; Salloum, J.; Jian, D.I.; Ng, P.K.W.; Gangur, V. Mechanisms of Wheat Allergenicity in Mice: Comparison of Adjuvant-Free vs. Alum-Adjuvant Models. Int. J. Mol. Sci. 2020, 21, 3205. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Model and Therapy | Protein Used | Sensitization Phenotype | Disease Phenotype |
|---|---|---|---|
| Mitigation of gluten (wheat) allergenicity using thioredoxin treated wheat flour [45] | Wheat gluten | Specific IgE | Wheal and flair reaction upon SPT |
| Mitigation of gluten (wheat) allergenicity using HKL treatment of dogs [46] | Wheat flour | Specific IgE | Increase in minimum dose require to elicit positive SPT reaction |
| Model and Therapy | Protein Used | Sensitization Phenotype | Disease Phenotype |
|---|---|---|---|
| Injection of 1BS-18 gluten lacking ω-5 gliadin reduced anaphylaxis in BN rats [47] | Gluten from Hokushin and 1BS-18 wheat | Specific IgE | HSR decreased |
| Induction of oral tolerance by early ingestion of 1BS-18 gluten that is lacking ω-5 gliadin in BN rats [48] | Commercial gluten, gluten prepared from Hokushin, and 1BS-18 wheat flours | Specific IgE | HSR decreased |
| Induction of oral tolerance using deamidated gliadin in BN rats [49] | Gliadin and deamidated gliadin | Specific IgE | None |
| Enzyme-treated gluten reduced airway allergenicity in BN rats [50] | Gluten and hydrolyzed gluten with cellulase and actinase | Specific IgE | Reduced BAL immune cells (neutrophils, lymphocytes, and eosinophils) |
| Mitigation of oral allergy using 1BS-18 gluten lacking ω-5 gliadin in Guinea Pigs [51] | Commercial gluten and 1BS-18 gluten | None | Significant decrease in allergy scores |
| Model and Therapy | Protein Used | Sensitization Phenotype | Disease Phenotype |
|---|---|---|---|
| Mitigation of gluten anaphylaxis using oral administration of Pediococcus acidilactici XZ31 in Balb/c mice [52] | Commercial wheat gluten | Specific IgE | Clinical symptom scores of systemic anaphylaxis |
| Enzyme-hydrolyzed gluten reduced sensitization in Balb/c mice [53] | Hydrolyzed wheat gluten with AN-PEP | Specific IgE | None |
| Oral treatment with L-arabinose reduced anaphylaxis symptoms in Balb/c mice [54] | Commercial gliadin | Total IgE | Clinical symptom scores of systemic anaphylaxis, allergic enteritis (histology, jejunum) |
| Hydrolysis and deamidation of gliadin reduced sensitization in Balb/c mice [55] | Hydrolyzed and deamidated gliadin | Specific IgE | None |
| Phosphorylated gliadin, and enzyme treatment of gliadin reduced sensitization in Balb/c mice [56] | Gliadins, phosphorylated gliadins, hydrolyzed gliadins with alcalase or papain | Specific and total IgE | None |
| Repeated oral administration of deamidated gliadin reduced allergenicity in native gliadin sensitized Balb/c mice [57] | Native gliadin and deamidated gliadin by carboxylation cation exchange resin | Specific IgE | Blood elevation of histamine |
| Model and Therapy | Protein Used | Sensitization Phenotype | Disease Phenotype |
|---|---|---|---|
| In vitro model, soy sauce fermentation reduced/eliminated gluten allergenicity [63] | Non-gluten and gluten | Specific IgE | None |
| In vitro model, gene targeting to remove conventional gluten reduced gluten allergenicity [64,65,66,67] | Gluten from gene targeted wheats using various techniques (Gene Translocation, Gene inactivation, gene deletion and gene silencing) | Specific IgE | None |
| In vitro model, thioredoxin treatment of gluten reduced its allergenicity [68] | Alcohol-soluble gluten extract was treated with thioredoxin | Specific IgE | None |
| In vivo and in vitro model, diploid genotype (AA) reduced gluten allergenicity [69] | Alcohol (40%) soluble gluten extract | Specific IgE | Negative SPT reaction in WDEIA 13/14 patients |
| In vivo and in vitro model, enzyme hydrolyzed wheat flour reduced gluten allergenicity [70] | Enzyme hydrolyzed wheat flour | Specific IgE | 13/15 AD patients tolerated cupcakes made from treated flour: 2/15 developed severe urticaria |
| Soy Sauce Country of Origin | Mold * | Yeast * | Gram-Positive Bacteria * | Gram-Negative Bacteria * |
|---|---|---|---|---|
| Korea [71] | Tetrapisispora Cryptococcus Penicillium Aspergillus sp., A. flavus: (6/6) | Wickerhamomyces | Not Studied | Not Studied |
| Torulaspora | ||||
| Tetrapisispora | ||||
| Rhodotorula | ||||
| Pichia | ||||
| Microbotryum | ||||
| Debaryomyces | ||||
| Candida: (6/6) | ||||
| Zygosaccharomyces | ||||
| China [72] | Aspergillus | Starmerella Wickerhamiella Saturnispora Candida | Weisella Bacillus: (3/5) Lactobacillus: (3/5) Leuconostoc Lactococcus Pediococcus Enterococcus Micrococcus Streptococcus Staphylococcus: (3/5) Propionibacteriacea | Xanthomonas Salmonella Pseudomonas Pantoea Lebsiella Dechloromonas Cupriavidus Arsenophonus Acidobacteriaceae |
| Japan [73] | Aspergillus: A. oryzae Geotrichum | Zygosacchormyces Candida etchellsii C. nodaensis C. versatilis C. catenulata Wickerhamomyces Pichia Trichosporon | Weisella Lactobacillus Staphylococcus gallinarum S. xylosus S.arlettae S. saprophyticus S. succinus S. cohnii S. caprae S. kloosii Pediococcus Tetragenococcus | Not studied |
| Not specified [74] | Aspergillus: Aspergillus sojae A. parasiticus Peronospora | Sacchoramycopsis Millerozyma Pichia Candida sp. C. rugosa C. orthopsilosis C. tropicalis | Staphylococcus Kurthia Bacillus Paenibacillus Corynebacterium | Klebsiella Enterobacter |
| China (LSSF) [75] | Aspergillus oryzae | Wickerhmomyces Saccharomycopsis Kluyveromyces Candida rugosa C. glabrata C. tropicalis Pichia Trichosporon | Weisella Bacillus subtilis B. licheniformis B. pumilus Staphylococcus sciuri S. gallinarum S. succinus S. aureus S. cohnii Corynebacterium Kurthia Enterococcus Lactobacillus Rothia Arhrobacter Pediococcus | Escherichia Enterobacter |
| China (Xianshi) [76] | Aspergillus niger Cladosporium Fusarium Lichtheimia Absidia | Meyerozyma Candida parapsilosis Sterigmatomyces | Bacillus amyloliquefaciens B. subtilis B.lincheniformis B. methylotrophicus B. aerius B. halmapalus B. flexus B. thuringiensis B. coagulan Scopulibacillus Shimwellia Weissella Lactococcus Clostridium Streptomyces Microlunatus | Klebsiella Pantoea Enterobacter Erwinia Trichodesmium |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jorgensen, R.; Devarahalli, S.S.; Shah, Y.; Gao, H.; Arul Arasan, T.S.; Ng, P.K.W.; Gangur, V. Advances in Gluten Hypersensitivity: Novel Dietary-Based Therapeutics in Research and Development. Int. J. Mol. Sci. 2024, 25, 4399. https://doi.org/10.3390/ijms25084399
Jorgensen R, Devarahalli SS, Shah Y, Gao H, Arul Arasan TS, Ng PKW, Gangur V. Advances in Gluten Hypersensitivity: Novel Dietary-Based Therapeutics in Research and Development. International Journal of Molecular Sciences. 2024; 25(8):4399. https://doi.org/10.3390/ijms25084399
Chicago/Turabian StyleJorgensen, Rick, Shambhavi Shivaramaiah Devarahalli, Yash Shah, Haoran Gao, Tamil Selvan Arul Arasan, Perry K. W. Ng, and Venugopal Gangur. 2024. "Advances in Gluten Hypersensitivity: Novel Dietary-Based Therapeutics in Research and Development" International Journal of Molecular Sciences 25, no. 8: 4399. https://doi.org/10.3390/ijms25084399
APA StyleJorgensen, R., Devarahalli, S. S., Shah, Y., Gao, H., Arul Arasan, T. S., Ng, P. K. W., & Gangur, V. (2024). Advances in Gluten Hypersensitivity: Novel Dietary-Based Therapeutics in Research and Development. International Journal of Molecular Sciences, 25(8), 4399. https://doi.org/10.3390/ijms25084399