
Modulation of the Circadian Rhythm and Oxidative Stress as Molecular Targets to Improve Vascular Dementia: A Pharmacological Perspective

 , , , and
, , , and
Abstract
:1. Introduction
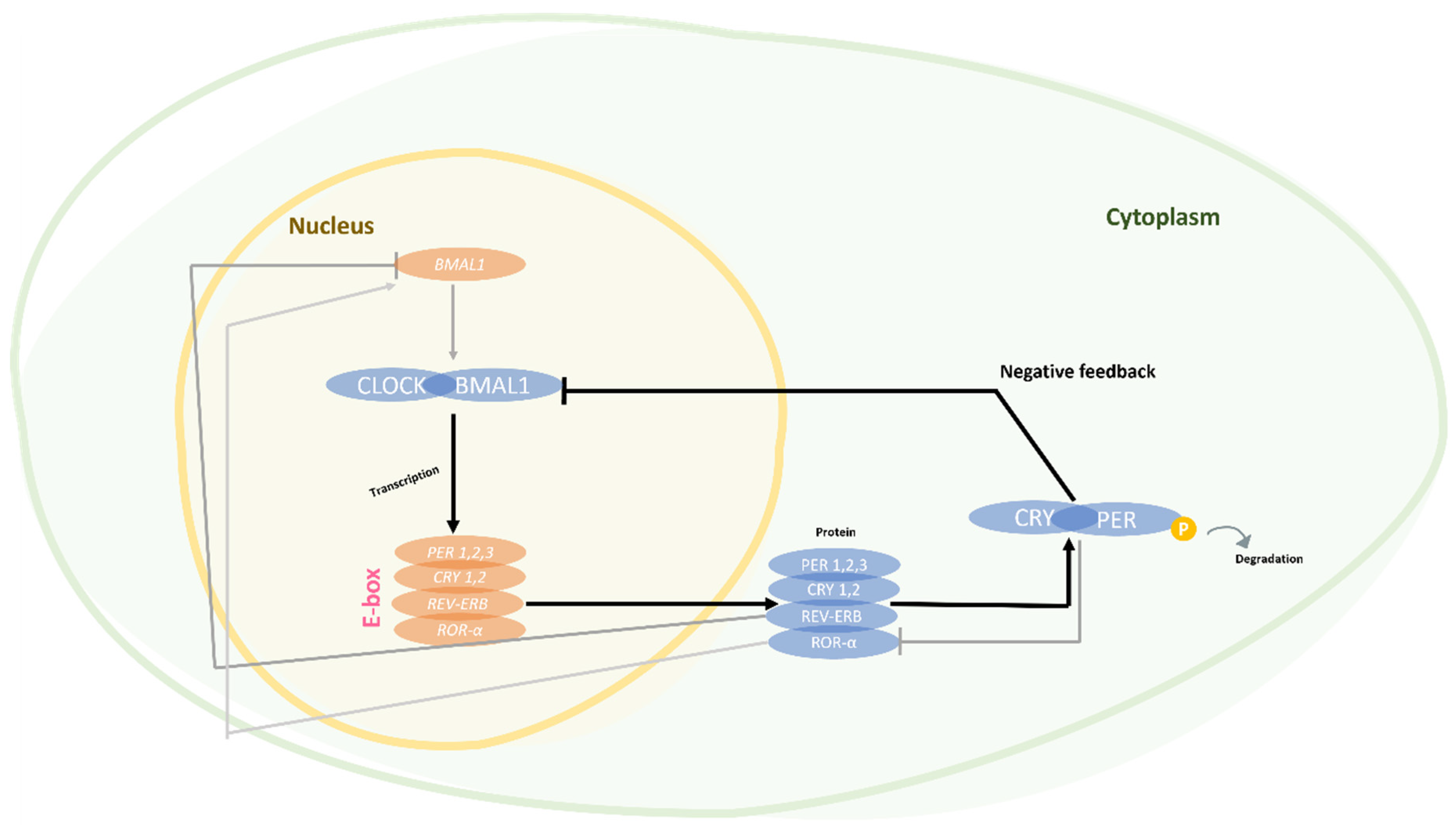
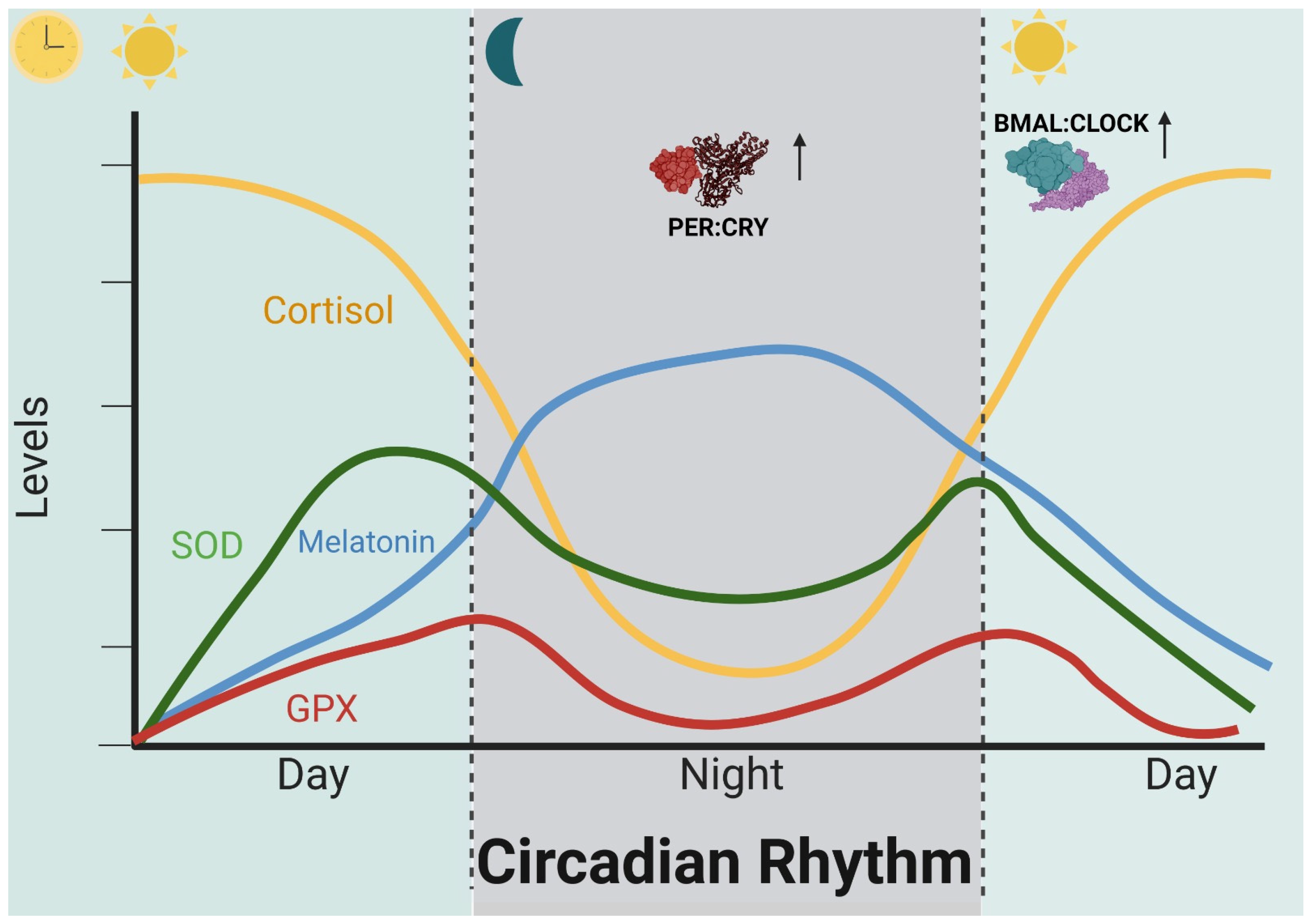
2. Circadian Rhythm and Vascular Dementia
3. Oxidative Stress and Vascular Dementia
Mitochondrial Dysfunction and Neuronal Cell Death in Vascular Dementia
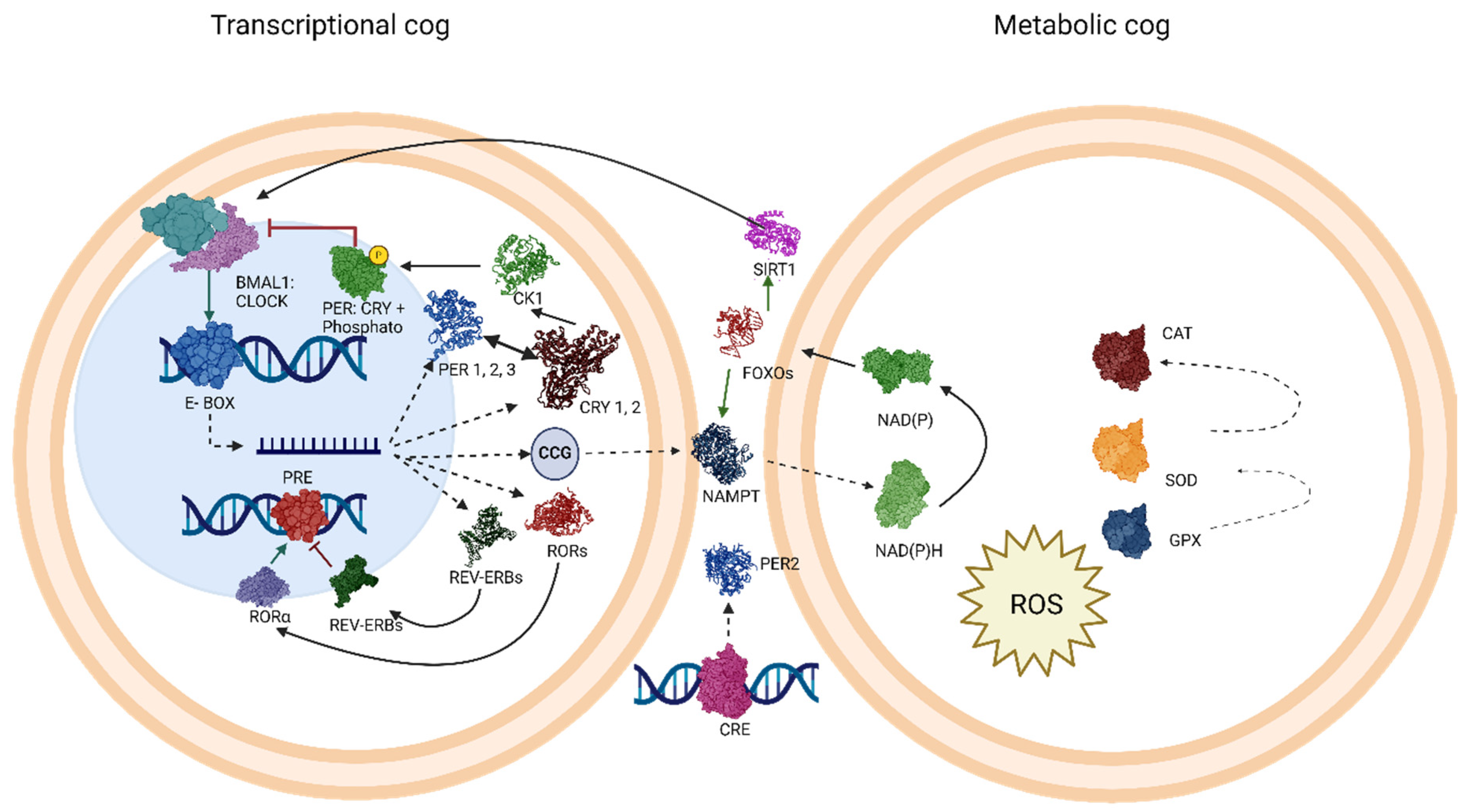
4. Molecular Targets Related to the Circadian Rhythm and Oxidative Stress in Vascular Dementia
5. Mechanism of Action: Molecular Targets Related to the Circadian Cycle and Oxidative Stress in Vascular Dementia
5.1. Pharmacological Interventions in the Circadian Cycle and Oxidative Stress
5.1.1. Melatonin
5.1.2. Nrf2 Activators
5.1.3. GLT2 Inhibitors
5.1.4. Nicotinamide
5.1.5. Phenolic Compounds
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garre-Olmo, J. Epidemiología de La Enfermedad de Alzheimer y Otras Demencias. Rev. Neuro 2018, 66, 377–386. [Google Scholar] [CrossRef]
- Rodríguez García, P.L.; Rodríguez García, D. Diagnóstico Del Deterioro Cognitivo Vascular y Sus Principales Categorías. Neurología 2015, 30, 223–239. [Google Scholar] [CrossRef]
- Vayas Abascal, R.; Carrera Romero, L. Disfunción Ejecutiva. Síntomas y Relevancia de Su Detección Desde Atención Primaria. Rev. Clin. Med. Fam. 2012, 5, 191. [Google Scholar] [CrossRef]
- Xu, Y.; Gong, C.; Liao, J.; Ge, Z.; Tan, Y.; Jiang, Y.; Liu, M.; Zhong, W.; Zhang, X.; Dong, N.; et al. Absence of Fluctuation and Inverted Circadian Rhythm of Blood Pressure Increase the Risk of Cognitive Dysfunction in Cerebral Small Vessel Disease Patients. BMC Neurol. 2023, 23, 73. [Google Scholar] [CrossRef]
- Nagai, M.; Hoshide, S.; Kario, K. Hypertension and Dementia. Am. J. Hypertens. 2010, 23, 116–124. [Google Scholar] [CrossRef]
- Thosar, S.S.; Berman, A.M.; Herzig, M.X.; McHill, A.W.; Bowles, N.P.; Swanson, C.M.; Clemons, N.A.; Butler, M.P.; Clemons, A.A.; Emens, J.S.; et al. Circadian Rhythm of Vascular Function in Midlife Adults. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1203–1211. [Google Scholar] [CrossRef]
- Maiese, K. Cognitive Impairment and Dementia: Gaining Insight through Circadian Clock Gene Pathways. Biomolecules 2021, 11, 1002. [Google Scholar] [CrossRef]
- Sharma, A.; Goyal, R. Long-Term Exposure to Constant Light Induces Dementia, Oxidative Stress and Promotes Aggregation of Sub-Pathological Aβ42 in Wistar Rats. Pharmacol. Biochem. Behav. 2020, 192, 172892. [Google Scholar] [CrossRef]
- Maiese, K. Neurodegeneration, Memory Loss, and Dementia: The Impact of Biological Clocks and Circadian Rhythm. Front. Biosci.—Landmark 2021, 26, 614–627. [Google Scholar] [CrossRef]
- Lax, P.; Ortuño-Lizarán, I.; Maneu, V.; Vidal-Sanz, M.; Cuenca, N. Molecular Sciences Photosensitive Melanopsin-Containing Retinal Ganglion Cells in Health and Disease: Implications for Circadian Rhythms. Int. J. Mol. Sci. 2019, 20, 3164. [Google Scholar] [CrossRef]
- Brown, T.M.; Piggins, H.D. Electrophysiology of the Suprachiasmatic Circadian Clock. Prog. Neurobiol. 2007, 82, 229–255. [Google Scholar] [CrossRef]
- Carter, B.; Justin, H.S.; Gulick, D.; Gamsby, J.J. The Molecular Clock and Neurodegenerative Disease: A Stressful Time. Front. Mol. Biosci. 2021, 8, 644747. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Zhang, T.; Lee, T.H. Cellular Mechanisms of Melatonin: Insight from Neurodegenerative Diseases. Biomolecules 2020, 10, 1158. [Google Scholar] [CrossRef] [PubMed]
- Downton, P.; Early, J.O.; Gibbs, J.E. Circadian Rhythms in Adaptive Immunity. Immunology 2020, 161, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Impacting Dementia and Cognitive Loss with Innovative Strategies: Mechanistic Target of Rapamycin, Clock Genes, Circular Non-Coding Ribonucleic Acids, and Rho/Rock. Neural Regen. Res. 2019, 14, 773–774. [Google Scholar] [CrossRef]
- Shi, L.; Chen, S.J.; Ma, M.Y.; Bao, Y.P.; Han, Y.; Wang, Y.M.; Shi, J.; Vitiello, M.V.; Lu, L. Sleep Disturbances Increase the Risk of Dementia: A Systematic Review and Meta-Analysis. Sleep Med. Rev. 2018, 40, 4–16. [Google Scholar] [CrossRef]
- Colwell, C.S. Defining Circadian Disruption in Neurodegenerative Disorders. J. Clin. Investig. 2021, 131, e148288. [Google Scholar] [CrossRef]
- Morton, A.J.; Wood, N.I.; Hastings, M.H.; Hurelbrink, C.; Barker, R.A.; Maywood, E.S. Disintegration of the Sleep-Wake Cycle and Circadian Timing in Huntington’s Disease. J. Neurosci. 2005, 25, 157–163. [Google Scholar] [CrossRef]
- Delgado-Lara, D.L.; González-Enríquez, G.V.; Torres-Mendoza, B.M.; González-Usigli, H.; Cárdenas-Bedoya, J.; Macías-Islas, M.A.; de la Rosa, A.C.; Jiménez-Delgado, A.; Pacheco-Moisés, F.; Cruz-Serrano, J.A.; et al. Effect of Melatonin Administration on the PER1 and BMAL1 Clock Genes in Patients with Parkinson’s Disease. Biomed. Pharmacother. 2020, 129, 110485. [Google Scholar] [CrossRef]
- Cronin, P.; McCarthy, M.J.; Lim, A.S.P.; Salmon, D.P.; Galasko, D.; Masliah, E.; De Jager, P.L.; Bennett, D.A.; Desplats, P. Circadian Alterations during Early Stages of Alzheimer’s Disease Are Associated with Aberrant Cycles of DNA Methylation in BMAL1. Alzheimer’s Dement. 2017, 13, 689–700. [Google Scholar] [CrossRef]
- Breen, D.P.; Vuono, R.; Nawarathna, U.; Fisher, K.; Shneerson, J.M.; Reddy, A.B.; Barker, R.A.; Van Geest, J. Sleep and Circadian Rhythm Regulation in Early Parkinson Disease. JAMA Neurol. 2014, 71, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Logan, R.W.; Ma, T.; Lewis, D.A.; Tseng, G.C.; Sibille, E.; McClung, C.A. Effects of Aging on Circadian Patterns of Gene Expression in the Human Prefrontal Cortex. Proc. Natl. Acad. Sci. USA 2016, 113, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.A.; Pijl, H.; Frö, M.; Van Der Graaf, A.W.M.; Roelfsema, F.; Roos, R.A.C. Increased Hypothalamic-Pituitary-Adrenal Axis Activity in Huntington’s Disease. J. Clin. Endocrinol. Metab. 2009, 94, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Kalliolia, E.; Silajdzic, E.; Nambron, R.; Hill, N.R.; Doshi, A.; Frost, C.; Watt, H.; Hindmarsh, P.; Bj, M.; Warner, T.T. Plasma Melatonin Is Reduced in Huntington’s Disease. Mov. Disord. 2014, 29, 1511–1515. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.L.; Harshman, L.A.; Kamholz, J.A.; Nopoulos, P.C. Autonomic Dysregulation as an Early Pathologic Feature of Huntington Disease. Auton. Neurosci. 2021, 231, 102775. [Google Scholar] [CrossRef] [PubMed]
- Raupach, A.K.; Ehgoetz Martens, K.A.; Memarian, N.; Zhong, G.; Matar, E.; Halliday, G.M.; Grunstein, R.; Lewis, S.J.G. Assessing the Role of Nocturnal Core Body Temperature Dysregulation as a Biomarker of Neurodegeneration. J. Sleep Res. 2020, 29, e12939. [Google Scholar] [CrossRef] [PubMed]
- Satlin, A.; Volicer, L.; Stopa, E.G.; Harper, D. Circadian Locomotor Activity and Core-Body Temperature Rhythms in Alzheimer’s Disease. Neurobiol. Aging 1995, 16, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Rodriguez, A.; Abreu-Gonzalez, P.; Sanchez-Sanchez, J.J.; Kaski, J.C.; Reiter, R.J. Melatonin and Circadian Biology in Human Cardiovascular Disease. J. Pineal Res. 2010, 49, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Syahira, A.; Azmi, M.; Juliana, N.; Azmani, S.; Effendy, N.M.; Fahmy Abu, I.; Mohd, N.I.; Teng, F.; Das, S. Cortisol on Circadian Rhythm and Its Effect on Cardiovascular System. J. Environ. Res. Public Health 2021, 17, 676. [Google Scholar] [CrossRef]
- Uchiyama, M.; Isse, K.; Tanaka, K.; Yokota, N.; Hamamoto, M.; Aida, S.; Ito, Y.; Yoshimura, M.; Okawa, M. Incidental Lewy Body Disease in a Patient with REM Sleepbehavior Disorder. Neurology 1995, 45, 709–712. [Google Scholar] [CrossRef]
- Daniela, M.; Grigoras, C.; Cuciureanu, D.; Constantinescu, V. The Circadian Rhythm of Arterial Blood Pressure in Alzheimer’s Disease and Vascular Dementia. Acta Neurol. Belg. 2023, 123, 129–137. [Google Scholar] [CrossRef]
- Maemura, K.; Takeda, N.; Nagai, R. Forum Minireview Circadian Rhythms in the CNS and Peripheral Clock Disorders: Role of the Biological Clock in Cardiovascular Diseases. J. Pharmacol. Sci. 2007, 103, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Tarquini, R.; Mazzoccoli, G. Clock Genes, Metabolism, and Cardiovascular Risk. Heart Fail. Clin. 2017, 13, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Duong, S.; Patel, T.; Chang, F. Dementia: What Pharmacists Need to Know. Can. Pharm. J. 2017, 150, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhong, R.J.; Cheng, C.; Li, S.; Le, W.D. New Therapeutics beyond Amyloid-β and Tau for the Treatment of Alzheimer’s Disease. Acta Pharmacol. Sin. 2021, 42, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Liu, S.; Sothern, R.B.; Xu, S.; Chan, P. Expression of Clock Genes Per1 and Bmal1 in Total Leukocytes in Health and Parkinson’s Disease. Eur. J. Neurol. 2010, 17, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. The Lancet Commissions Dementia Prevention, Intervention, and Care: 2020 Report of the Lancet Commission. Lancet 2020, 396, 413–459. [Google Scholar] [CrossRef] [PubMed]
- Ferini-Strambi, L. Sleep Disorders and Increased Risk of Dementia. Eur. J. Neurol. 2022, 29, 3484–3485. [Google Scholar] [CrossRef]
- Insel, P.S.; Mohlenhoff, B.S.; Neylan, T.C.; Krystal, A.D.; Mackin, R. Scott Association of Sleep and β-Amyloid Pathology Among Older Cognitively Unimpaired Adults. JAMA Netw. Open 2021, 4, e2117573. [Google Scholar] [CrossRef]
- Damsgaard, L.; Janbek, J.; Laursen, T.M. Hospital-Diagnosed Sleep Disorders and Incident Dementia: A Nationwide Observational Cohort Study. Eur. J. Neurol. 2022, 29, 3528–3536. [Google Scholar] [CrossRef]
- Krohn, L.; Heilbron, K.; Blauwendraat, C.; Reynolds, R.H.; Yu, E.; Senkevich, K.; Rudakou, U.; Estiar, M.A.; Gustavsson, E.K.; Brolin, K.; et al. Genome-Wide Association Study of REM Sleep Behavior Disorder Identifies Polygenic Risk and Brain Expression Effects. Nat. Commun. 2022, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.; Grant, M.M.; Aldred, S. Oxidative Stress in Vascular Dementia and Alzheimer’s Disease: A Common Pathology. J. Alzheimer’s Dis. 2009, 17, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Sienes Bailo, P.; Llorente Martín, E.; Calmarza, P.; Montolio Breva, S.; Bravo Gómez, A.; Pozo Giráldez, A.; José Sánchez-Pascuala Callau, J.; Maria Vaquer Santamaría, J.; Dayaldasani Khialani, A.; Cerdá Micó, C.; et al. Implicación Del Estrés Oxidativo En Las Neurodegenerativas y Posibles Antioxidantes. Adv. Lab. Med./Av. Med. Lab. 2022, 3, 351–360. [Google Scholar] [CrossRef]
- Hosoki, S.; Hansra, G.K.; Jayasena, T.; Poljak, A.; Mather, K.A.; Catts, V.S.; Rust, R.; Sagare, A.; Kovacic, J.C.; Brodtmann, A.; et al. Molecular Biomarkers for Vascular Cognitive Impairment and Dementia. Nat. Rev. Neurol. 2023, 19, 737–753. [Google Scholar] [CrossRef]
- Kattoor, A.J.; Pothineni, N.V.K.; Palagiri, D.; Mehta, J.L. Oxidative Stress in Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 1–11. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Chen, X.; Wei, Y. Neuronal Injuries in Cerebral Infarction and Ischemic Stroke: From Mechanisms to Treatment (Review). Int. J. Mol. Med. 2022, 49, 15. [Google Scholar] [CrossRef]
- Sinclair, A.J.; Bayer, A.J.; Johnston, J.O.; Warner, C.; Maxwell, S.R.J. Altered plasma antioxidant status in subjects with Alzheimer’s Disease and vascular dementia. Int. J. Geriatr. Psychiatry 1999, 12, 840–845. [Google Scholar] [CrossRef]
- Joshi, Y.B.; Praticò, D. Vitamin E in Aging, Dementia, and Alzheimer’s Disease. BioFactors 2012, 38, 90–97. [Google Scholar] [CrossRef]
- Manzano-León, N.; Mas-Oliva, J. Estrés Oxidativo, Péptido β-Amiloide Enfermedad de Alzheimer. Inst. Fisiol. Cel. 2006, 142, 229–238. [Google Scholar]
- Akbar, M.; Essa, M.M.; Daradkeh, G.; Abdelmegeed, M.A.; Choi, Y.; Mahmood, L.; Song, B.J. Mitochondrial Dysfunction and Cell Death in Neurodegenerative Diseases through Nitroxidative Stress. Brain Res. 2016, 1637, 34–55. [Google Scholar] [CrossRef]
- Yu, E.; Mercer, J.; Bennett, M. Mitochondria in Vascular Disease. Cardiovasc. Res. 2012, 95, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Mikhed, Y.; Daiber, A.; Steven, S.; Olson, L.; Ross, J.M.; Coppotelli, G. Mitochondrial Oxidative Stress, Mitochondrial DNA Damage and Their Role in Age-Related Vascular Dysfunction. Int. J. Mol. Sci. 2015, 16, 15918–15953. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Horke, S.; Förstermann, U. Oxidative Stress in Vascular Disease and Its Pharmacological Prevention. Trends Pharmacol. Sci. 2013, 34, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T. Oxidative Stress and Mitochondrial Dysfunction-Linked Neurodegenerative Disorders. Neurol. Res. 2017, 39, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative Stress, Mitochondrial Dysfunction and Neurodegenerative Diseases; a Mechanistic Insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Budkowska, M.; Zbieta Cecerska-Hery’chery´c, E.; Marcinowska, Z.; Siennicka, A.; Dołęgowska, B. The Influence of Circadian Rhythm on the Activity of Oxidative Stress Enzymes. Int. J. Mol. Sci. 2022, 2022, 14275. [Google Scholar] [CrossRef] [PubMed]
- McClean, C.; Davison, G.W. Circadian Clocks, Redox Homeostasis, and Exercise: Time to Connect the Dots? Antioxidants 2022, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Lananna, B.V.; Musiek, E.S. The Wrinkling of Time: Aging, Inflammation, Oxidative Stress, and the Circadian Clock in Neurodegeneration. Neurobiol. Dis. 2020, 139, 104832. [Google Scholar] [CrossRef] [PubMed]
- Musiek, E.S.; Lim, M.M.; Yang, G.; Bauer, A.Q.; Qi, L.; Lee, Y.; Roh, J.H.; Ortiz-Gonzalez, X.; Dearborn, J.T.; Culver, J.P.; et al. Circadian Clock Proteins Regulate Neuronal Redox Homeostasis and Neurodegeneration. J. Clin. Investig. 2013, 123, 5389. [Google Scholar] [CrossRef]
- Kress, G.J.; Liao, F.; Dimitry, J.; Cedeno, M.R.; FitzGerald, G.A.; Holtzman, D.M.; Musiek, E.S. Regulation of Amyloid-β Dynamics and Pathology by the Circadian Clock. J. Exp. Med. 2018, 215, 1059–1068. [Google Scholar] [CrossRef]
- Sharma, A.; Sethi, G.; Tambuwala, M.M.; Aljabali, A.A.A.; Chellappan, D.K.; Dua, K.; Goyal, R. Circadian Rhythm Disruption and Alzheimer’s Disease: The Dynamics of a Vicious Cycle. Curr. Neuropharmacol. 2021, 19, 248. [Google Scholar] [CrossRef]
- Ralph, M.R.; Menaker, M. A Mutation of the Circadian System in Golden Hamsters. Science 1988, 241, 1225–1227. [Google Scholar] [CrossRef]
- Karolczak, K.; Watala, C. Melatonin as a Reducer of Neuro-and Vasculotoxic Oxidative Stress Induced by Homocysteine. Antioxidants 2021, 10, 1178. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Z.; Cai, Y.; Zeng, S.; Peng, B.; Ren, X.; Yan, Y.; Gong, Z. Rheostatic Balance of Circadian Rhythm and Autophagy in Metabolism and Disease. Front. Cell Dev. Biol. 2020, 8, 616434. [Google Scholar] [CrossRef]
- Rami, A.; Fekadu, J.; Rawashdeh, O. The Hippocampal Autophagic Machinery Is Depressed in the Absence of the Circadian Clock Protein PER1 That May Lead to Vulnerability During Cerebral Ischemia. Curr. Neurovasc Res. 2017, 14, 207–214. [Google Scholar] [CrossRef]
- Crnko, S.; Cour, M.; Van Laake, L.W.; Lecour, S. Vasculature on the Clock: Circadian Rhythm and Vascular Dysfunction. Vascul Pharmacol. 2018, 108, 1–7. [Google Scholar] [CrossRef]
- Elykova, A.V.; Orlova, V.S.; Batlutskaya, I.V.; Sorokina, I.N.; Efremova, O.A.; Rudyh, N.A.; Churnosov, M.I. Polymorphism of Vascular Homeostasis Genes as a Risk Factor for the Formation of Preeclampsia. Meditsinskiy Sov. 2023, 17, 98–105. [Google Scholar] [CrossRef]
- Mishima, K.; Okawa, M.; Satoh, K.; Shimizu, T.; Hozumi, S.; Hishikawa, Y. Different Manifestations of Circadian Rhythms in Senile Dementia of Alzheimer’s Type and Multi-Infarct Dementia. Neurobiol. Aging 1997, 18, 105–109. [Google Scholar] [CrossRef]
- Cermakian, N.; Waddington Lamont, E.; Boudreau, P.; Boivin, D.B. Circadian Clock Gene Expression in Brain Regions of Alzheimer’s Disease Patients and Control Subjects. J. Biol. Rhythms 2011, 26, 160–170. [Google Scholar] [CrossRef]
- Sari, A.; Davutoglu, V.; Bozkurt, E.; Taner, I.L.; Erciyas, K. Effect of Periodontal Disease on Oxidative Stress Markers in Patients with Atherosclerosis. Clin. Oral. Investig. 2022, 26, 1713–1724. [Google Scholar] [CrossRef]
- Hossain, M.F.; Wang, N.; Chen, R.; Li, S.; Roy, J.; Uddin, M.G.; Li, Z.; Lim, L.W.; Song, Y.Q. Exploring the Multifunctional Role of Melatonin in Regulating Autophagy and Sleep to Mitigate Alzheimer’s Disease Neuropathology. Ageing Res. Rev. 2021, 67, 101304. [Google Scholar] [CrossRef]
- Martín Giménez, V.M.; de las Heras, N.; Ferder, L.; Lahera, V.; Reiter, R.J.; Manucha, W. Potential Effects of Melatonin and Micronutrients on Mitochondrial Dysfunction during a Cytokine Storm Typical of Oxidative/Inflammatory Diseases. Diseases 2021, 9, 30. [Google Scholar] [CrossRef]
- Hossain, M.F.; Uddin, M.S.; Uddin, G.M.S.; Sumsuzzman, D.M.; Islam, M.S.; Barreto, G.E.; Mathew, B.; Ashraf, G.M. Melatonin in Alzheimer’s Disease: A Latent Endogenous Regulator of Neurogenesis to Mitigate Alzheimer’s Neuropathology. Mol. Neurobiol. 2019, 56, 8255–8276. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, H.; Cao, Y.; Ma, L.; Pei, H.; Rausch, W.D. Dysfunction of Cerebrovascular Endothelial Cells: Prelude to Vascular Dementia. Front. Aging Neurosci. 2018, 10, 376. [Google Scholar] [CrossRef]
- Wongprayoon, P.; Govitrapong, P. Melatonin Receptor as a Drug Target for Neuroprotection. Curr. Mol. Pharmacol. 2020, 14, 150–164. [Google Scholar] [CrossRef] [PubMed]
- Thiruvengadam, M.; Venkidasamy, B.; Subramanian, U.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Girish, S.; Thangavel, S.; Dhanapal, A.R.; Fedoseeva, N.; et al. Antioxidants Bioactive Compounds in Oxidative Stress-Mediated Diseases: Targeting the NRF2/ARE Signaling Pathway and Epigenetic Regulation. Antioxidants 2021, 10, 1859. [Google Scholar] [CrossRef]
- Morroni, F.; Sita, G.; Graziosi, A.; Turrini, E.; Fimognari, C.; Tarozzi, A.; Hrelia, P. Neuroprotective Effect of Caffeic Acid Phenethyl Ester in a Mouse Model of Alzheimer’s Disease Involves Nrf2/HO-1 Pathway. Aging Dis. 2017, 9, 605–622. [Google Scholar] [CrossRef]
- Ren, P.; Chen, J.; Li, B.; Zhang, M.; Yang, B.; Guo, X.; Chen, Z.; Cheng, H.; Wang, P.; Wang, S.; et al. Nrf2 Ablation Promotes Alzheimer’s Disease-Like Pathology in APP/PS1 Transgenic Mice: The Role of Neuroinflammation and Oxidative Stress. Oxid. Med. Cell Longev. 2020, 2020, 3050971. [Google Scholar] [CrossRef]
- Pawlos, A.; Broncel, M.; Woźniak, E.; Gorzelak-Pabiś, P. Neuroprotective Effect of SGLT2 Inhibitors. Molecules 2021, 26, 7213. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Nohara, K.; Wirianto, M.; Escobedo, G.; Lim, J.Y.; Morales, R.; Yoo, S.H.; Chen, Z. Effects of the Clock Modulator Nobiletin on Circadian Rhythms and Pathophysiology in Female Mice of an Alzheimer’s Disease Model. Biomolecules 2021, 11, 1004. [Google Scholar] [CrossRef]
- Maiese, K. Nicotinamide: Oversight of Metabolic Dysfunction Through SIRT1, MTOR, and Clock Genes. Curr. Neurovasc Res. 2020, 17, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Figarola-Centurión, I.; Escoto-Delgadillo, M.; González-Enríquez, G.V.; Gutiérrez-Sevilla, J.E.; Vázquez-Valls, E.; Torres-Mendoza, B.M. Sirtuins Modulation: A Promising Strategy for HIV-Associated Neurocognitive Impairments. Int. J. Mol. Sci. 2022, 23, 643. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fuentes, C.; Suárez, M.; Aragonès, G.; Mulero, M.; Ávila-Román, J.; Arola-Arnal, A.; Salvadó, M.J.; Arola, L.; Bravo, F.I.; Muguerza, B. Cardioprotective Properties of Phenolic Compounds: A Role for Biological Rhythms. Mol. Nutr. Food Res. 2022, 66, e2100990. [Google Scholar] [CrossRef] [PubMed]
- Elgadir, M.A.; Chigurupati, S.; Mariod, A.A. Selected Potential Pharmaceutical and Medical Benefits of Phenolic Compounds: Recent Advances. Funct. Food Sci. 2023, 3, 108. [Google Scholar] [CrossRef]
- Hoyt, K.R.; Obrietan, K. Circadian Clocks, Cognition, and Alzheimer’s Disease: Synaptic Mechanisms, Signaling Effectors, and Chronotherapeutics. Mol. Neurodegener. 2022, 17, 35. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peripheral Markers | Molecular Markers | |
|---|---|---|
| Huntington’s disease | Melatonin: reduced levels, delayed phase of secretion, and becomes more delayed with disease progression [17,23,24] Cortisol: some patients have altered levels in the morning. There is a higher amplitude of the rhythm. The greater the progression of the disease, the greater the phase advance, and the higher the plasma levels [17]. CBT: elevated during daytime in presymptomatic HD patients [25]. | Disruption of the mPer2 and mBmal1 circadian clock genes in the suprachiasmatic nucleus of R6/2 mice [18]. |
| Parkinson’s disease | Melatonin: reduction in amplitude of secretion [17]. Cortisol: higher levels in plasma compared to healthy patients of the same age, and a decreased amplitude of the secretion rhythm [17]. CBT: amplitude of the rhythm was reduced in patients with RBD and dementia with Lewy bodies [26]. | Loss of rhythms in BMAL1 and altered expression of PER2 and Rev-Erbα [19]. |
| Alzheimer’s disease | Melatonin: decreased levels, delayed phase, and reduced amplitude [17]. Cortisol: higher levels in plasma compared to healthy patients of the same age, and phase-advanced rhythm. CBT: delayed phase [27]. | Diurnal rhythm pattern was lost in pineal gland. Rhythmic methylation of BMAL1 was altered in fibroblasts [20]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillo-Rangel, W.Á.; Acuña-Vaca, S.; Padilla-Ponce, D.J.; García-Mercado, F.G.; Torres-Mendoza, B.M.; Pacheco-Moises, F.P.; Escoto-Delgadillo, M.; García-Benavides, L.; Delgado-Lara, D.L.C. Modulation of the Circadian Rhythm and Oxidative Stress as Molecular Targets to Improve Vascular Dementia: A Pharmacological Perspective. Int. J. Mol. Sci. 2024, 25, 4401. https://doi.org/10.3390/ijms25084401
Trujillo-Rangel WÁ, Acuña-Vaca S, Padilla-Ponce DJ, García-Mercado FG, Torres-Mendoza BM, Pacheco-Moises FP, Escoto-Delgadillo M, García-Benavides L, Delgado-Lara DLC. Modulation of the Circadian Rhythm and Oxidative Stress as Molecular Targets to Improve Vascular Dementia: A Pharmacological Perspective. International Journal of Molecular Sciences. 2024; 25(8):4401. https://doi.org/10.3390/ijms25084401
Chicago/Turabian StyleTrujillo-Rangel, Walter Ángel, Sofía Acuña-Vaca, Danna Jocelyn Padilla-Ponce, Florencia Guillermina García-Mercado, Blanca Miriam Torres-Mendoza, Fermín P. Pacheco-Moises, Martha Escoto-Delgadillo, Leonel García-Benavides, and Daniela L. C. Delgado-Lara. 2024. "Modulation of the Circadian Rhythm and Oxidative Stress as Molecular Targets to Improve Vascular Dementia: A Pharmacological Perspective" International Journal of Molecular Sciences 25, no. 8: 4401. https://doi.org/10.3390/ijms25084401
APA StyleTrujillo-Rangel, W. Á., Acuña-Vaca, S., Padilla-Ponce, D. J., García-Mercado, F. G., Torres-Mendoza, B. M., Pacheco-Moises, F. P., Escoto-Delgadillo, M., García-Benavides, L., & Delgado-Lara, D. L. C. (2024). Modulation of the Circadian Rhythm and Oxidative Stress as Molecular Targets to Improve Vascular Dementia: A Pharmacological Perspective. International Journal of Molecular Sciences, 25(8), 4401. https://doi.org/10.3390/ijms25084401