
Glycosylation and Characterization of Human Transferrin in an End-Stage Kidney Disease

 , , , ,
, , , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Hematological and Biochemical Parameters in Patients on Peritoneal Dialysis
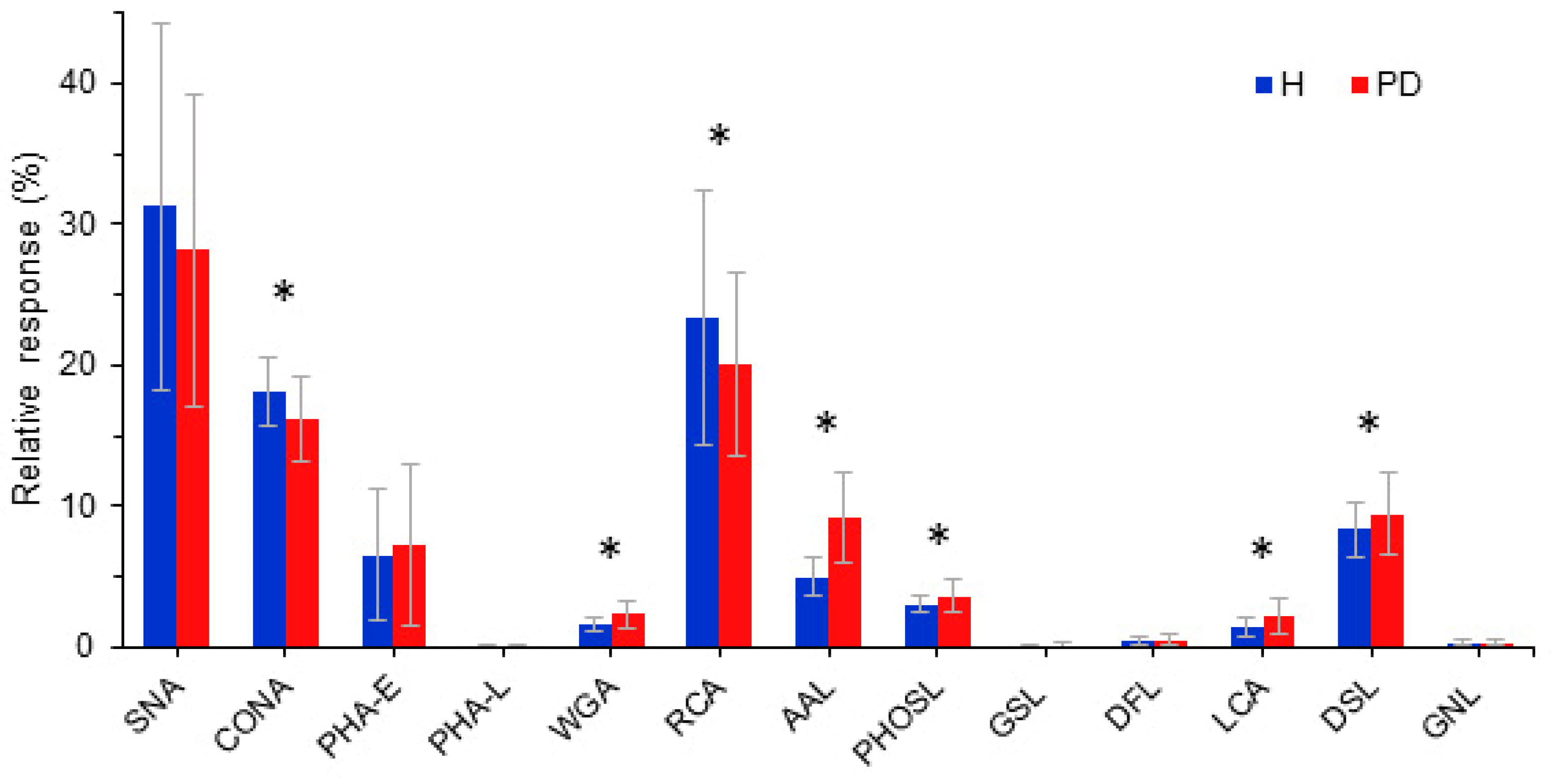
2.2. Glycosylation of PD Patients Transferrin
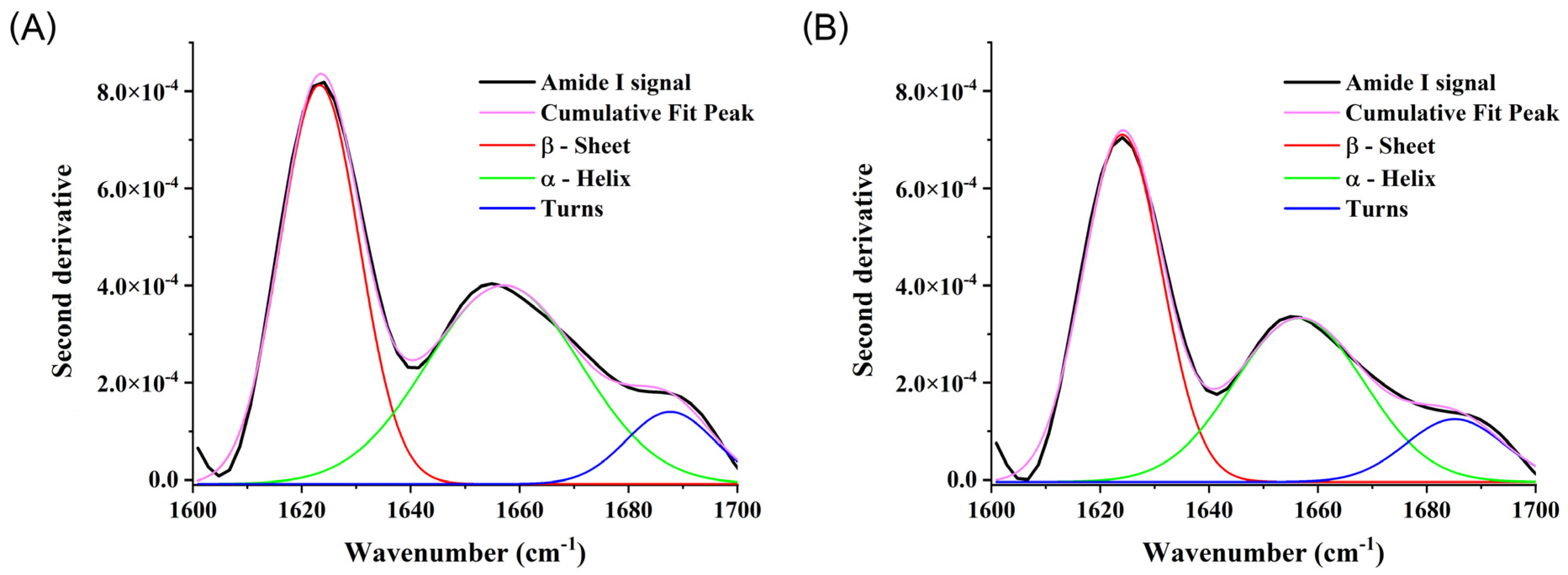
2.3. Secondary and Tertiary Structures of PD Transferrin
2.4. Functional Analysis of PD Transferrin
3. Discussion
4. Materials and Methods
4.1. Blood Samples
4.2. Isolation of Transferrin
4.3. Lectin-Based Microarray Analysis of Transferrin
4.4. Fourier-Transform Infrared Spectroscopic (FTIR) Analysis of Transferrin
4.5. Fluorescence Emission Spectroscopic Analysis of Transferrin
4.6. UV-VIS Spectrophotometric Analysis of Transferrin
4.7. Functional Analysis of Transferrin via Immunocytochemistry
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalantar-Zadeh, K.; Jafar, T.H.; Nitsch, D.; Neuen, B.L.; Perkovic, V. Chronic kidney disease. Lancet 2021, 398, 786–802. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, U.; Toto, R.D. Anemia, Diabetes, and Chronic Kidney Disease. Diabetes Care 2009, 32, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Bikbov, B.; Purcell, C.A.; Levey, A.S.; Smith, M.; Abdoli, A.; Abebe, M.; Adebayo, O.M.; Afarideh, M.; Agarwal, S.K.; Agudelo-Botero, M.; et al. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Hill, N.R.; Fatoba, S.T.; Oke, J.L.; Hirst, J.A.; O’Callaghan, C.A.; Lasserson, D.S.; Hobbs, F.D.R. Global prevalence of chronic kidney disease—A systematic review and meta-analysis. PLoS ONE 2016, 11, e0158765. [Google Scholar] [CrossRef]
- Vanholder, R.; Annemans, L.; Brown, E.; Gansevoort, R.; Gout-Zwart, J.J.; Lameire, N.; Morton, R.L.; Oberbauer, R.; Postma, M.J. Reducing the costs of chronic kidney disease while delivering quality health care: A call to action. Nat. Rev. Nephrol. 2017, 13, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Carney, E.F. The impact of chronic kidney disease on global health. Nat. Rev. Nephrol. 2020, 16, 251. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, P.; Remuzzi, G.; Glassock, R.; Levin, A.; Jager, K.J.; Tonelli, M.; Massy, Z.; Wanner, C.; Anders, H.-J. Chronic kidney disease. Nat. Rev. Dis. Primers 2017, 3, 17088. [Google Scholar] [CrossRef] [PubMed]
- Levey, A.S.; Eckardt, K.-U.; Dorman, N.M.; Christiansen, S.L.; Cheung, M.; Jadoul, M.; Winkelmayer, W.C. Nomenclature for kidney function and disease: Report of a Kidney Disease: Improving Global Outcomes (KDIGO) consensus conference. Kidney Int. Rep. 2020, 97, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Zoccali, C.; Kramer, A.; Jager, K.J. Chronic kidney disease and end-stage renal disease—A review produced to contribute to the report ‘the status of health in the European Union: Towards a healthier Europe’. NDT Plus 2010, 3, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Teitelbaum, I. Peritoneal dialysis. New Engl. J. Med. 2021, 385, 1786–1795. [Google Scholar] [CrossRef] [PubMed]
- Ferrantelli, E.; Farhat, K.; Ederveen, A.L.H.; Reiding, K.R.; Beelen, R.H.J.; van Ittersum, F.J.; Wuhrer, M.; Dotz, V. Effluent and serum protein N-glycosylation is associated with inflammation and peritoneal membrane transport characteristics in peritoneal dialysis patients. Sci. Rep. 2018, 8, 979. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.M. Peritoneal protein loss, leakage or clearance in peritoneal dialysis, where do we stand? Periton Dial. Int. 2019, 39, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Wada, Y. Mass spectrometry of transferrin glycoforms to detect congenital disorders of glycosylation: Site-specific profiles and pitfalls. Proteomics 2016, 16, 3105–3110. [Google Scholar] [CrossRef] [PubMed]
- Nagae, M.; Morita-Matsumoto, K.; Arai, S.; Wada, I.; Matsumoto, Y. Structural change of N-glycan exposes hydrophobic surface of human transferrin. Glycobiology 2014, 24, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Satomi, Y.; Shimonishi, Y.; Hase, T.; Takao, T. Site-specific carbohydrate profiling of human transferrin by nano-flow liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass. Spectrom. 2004, 18, 2983–2988. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Andrews, N.C.; Schmidt, P.J. Iron homeostasis. Annu. Rev. Physiol. 2007, 69, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.R.; Ponka, P. The molecular mechanisms of the metabolism and transport of iron in normal and neoplastic cells. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1997, 1331, 1–40. [Google Scholar] [CrossRef]
- Jansen, J.C.; van Hoek, B.; Metselaar, H.J.; van den Berg, A.P.; Zijlstra, F.; Huijben, K.; van Scherpenzeel, M.; Drenth, J.P.H.; Lefeber, D.J. Screening for abnormal glycosylation in a cohort of adult liver disease patient. J. Inherit. Metab. Dis. 2020, 43, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Hülsmeier, A.J.; Tobler, M.; Burda, P.; Hennet, T. Glycosylation site occupancy in health, congenital disorder of glycosylation and fatty liver disease. Sci. Rep. 2016, 6, 33927. [Google Scholar] [CrossRef] [PubMed]
- Rysz, J.; Franczyk, B.; Ławiński, J.; Gluba-Brzózka, A. Oxidative stress in ESRD patients on dialysis and the risk of cardiovascular diseases. Antioxidants 2020, 9, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Lange, R.; Balny, C. UV-visible derivative spectroscopy under high pressure. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzym. 2002, 1595, 80–93. [Google Scholar] [CrossRef]
- Mehrotra, R.; Devuyst, O.; Davies, S.J.; Johnson, D.W. The current state of peritoneal dialysis. J. Am. Soc. Nephrol. 2016, 27, 3238–3252. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Reed, J.; Noble, S.; Cangiano, J.L.; Van Wyck, D.B. United States Iron Sucrose (Venofer) Clinical Trials Group. Effect of intravenous iron sucrose in peritoneal dialysis patients who receive erythropoiesis-stimulating agents for anemia: A randomized, controlled trial. Clin. J. Am. Soc. Nephrol. 2006, 1, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Zheng, Z.; Yi, C.; Cao, P.; Ye, H.; Liu, R.; Lin, J.; Chen, W.; Mao, H.; Huang, F.; et al. Association of Abnormal Iron Status with the Occurrence and Prognosis of Peritoneal Dialysis-Related Peritonitis: A Longitudinal Data-Based 10-Year Retrospective Study. Nutrients 2022, 14, 1613. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Y.; Chuang, C.L.; Lin, C.C.; Tsai, S.H.; Chen, J.Y. Peritoneal-Membrane Characteristics and Hypervolemia Management in Peritoneal Dialysis: A Randomized Control Trial. Membranes 2021, 11, 768. [Google Scholar] [CrossRef] [PubMed]
- Di-Gioia, M.C.; Gallar, P.; Rodriguez, I.; Laso, N.; Callejas, R.; Ortega, O.; Herrero, J.C.; Vigil, A. Changes in body composition parameters in patients on haemodialysis and peritoneal dialysis. Nefrologia 2012, 32, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.A.; Blake, P.G.; Boudville, N.; Davies, S.; de Arteaga, J.; Dong, J.; Finkelstein, F.; Foo, M.; Hurst, H.; Johnson, D.W.; et al. International Society for Peritoneal Dialysis practice recommendations: Prescribing high-quality goal-directed peritoneal dialysis. Periton Dial. Int. 2020, 40, 244–253. [Google Scholar] [CrossRef]
- Ghanbari, Z.; Housaindokht, M.R.; Bozorgmehr, M.R.; Izadyar, M. The effect of glycosylation on the transferrin structure: A molecular dynamic simulation analysis. J. Theor. Biol. 2016, 404, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, H. Transferrin and transferrin receptors update. Free Rad. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Formanowicz, D.; Formanowicz, P. Transferrin changes in haemodialysed patients. Int. Urol. Nephrol. 2012, 44, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Wish, J.B. Assessing iron status: Beyond serum ferritin and transferrin saturation. Clin. J. Am. Soc. Nephrol. 2006, 1, S4–S8. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.-L.; Regoeczi, E. Some observations on the carbohydrate composition of purified transferrin. Int. J. Pept. Protein Res. 1977, 9, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lang, M. New insight into protein glycosylation in the development of Alzheimer’s disease. Cell Death Discov. 2023, 9, 314. [Google Scholar] [CrossRef] [PubMed]
- Babitt, J.L.; Lin, H.Y. Mechanisms of anemia in CKD. J. Am. Soc. Nephrol. 2012, 23, 1631–1634. [Google Scholar] [CrossRef] [PubMed]
- Portolés, J.; Martín, L.; Broseta, J.J.; Cases, A. Anemia in chronic kidney disease: From pathophysiology and current treatments, to future agents. Front. Med. 2021, 8, 642296. [Google Scholar] [CrossRef]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- An, H.J.; Froehlich, J.W.; Lebrilla, C.B. Determination of glycosylation sites and site-specific heterogeneity in glycoproteins. Curr. Opin. Chem. Biol. 2009, 13, 421–426. [Google Scholar] [CrossRef]
- Butler, M.; Perreault, H. Protein Glycosylation: Methods for Determination. Protein Glycosylation: Methods for Determination. In Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology, 1st ed.; Flickinger, M.C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA. [CrossRef]
- Zhao, N.; Zhang, A.S.; Worthen, C.; Knutson, M.D.; Enns, C.A. An iron-regulated and glycosylation-dependent proteasomal degradation pathway for the plasma membrane metal transporter ZIP14. Proc. Natl. Acad. Sci. USA 2014, 111, 9175–9180. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kalantar-Zadeh, K. Haemodialysis-induced hypoglycaemia and glycaemic disarrays. Nat. Rev. Nephrol. 2015, 11, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Penezić, A.; Miljuš, G.; Milutinović, B.; Nedić, O. A microscale protocol for the isolation of transferrin directly from serum. Clin. Chim. Acta 2017, 471, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Baralić, M.; Gligorijević, N.; Brković, V.; Katrlík, J.; Pažitná, L.; Šunderić, M.; Miljuš, G.; Penezić, A.; Dobrijević, Z.; Laušević, M.; et al. Fibrinogen fucosylation as a prognostic marker of end-stage renal disease in patients on peritoneal dialysis. Biomolecules 2020, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Beniash, E.; Yamakoshi, Y.; Simmer, J.P.; Margolis, H.C. Protein phosphorylation and mineral binding affect the secondary structure of the Leucine-Rich Amelogenin Peptide. Front. Physiol. 2017, 8, 450. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration | Patients on PD (n = 75) | Reference Range |
|---|---|---|
| Erythrocytes (×1012/L) | 3.4 (0.3–4.3) | 4.3–5.7 |
| Hemoglobin (g/L) | 103 (72–129) | 138–175 |
| Hematocrit (L/L) | 0.32 (0.22–0.41) | 0.41–0.53 |
| Total protein (g/L) | 64.0 (54.9–77.0) | 62.0–81.0 |
| Albumin (g/L) | 37.0 (28.0–44.1) | 35.0–53.0 |
| TIBC (µmol/L) | 44.7 (27.2–57.8) | 44.8–75.1 |
| Transferrin (g/L) | 1.7 (0.9–2.6) | 1.7–3.8 |
| Ferritin (µg/L) | 227 (47–1587) | 30–400 |
| Iron (µmol/L) | 11.5 (7.6–19.7) | 11.0–30.0 |
| Transferrin saturation (%) | 28.0 (17.9–49.3) | 20.0–50.0 |
| Lectin (Source) | Glycan Specificity | Reactivity with Tf |
|---|---|---|
| SNA (Sambucus nigra) | NeuNAcα2,6Gal/GalNAc | ++ |
| ConA (Conavalia ensiformis) | Manα1,6(Manα1,3)Man | ++ |
| MAL-I (Maackia amurensis) | NeuNAcα2,3Galβ1,4GlcNAc | - |
| MAL-II (Maackia amurensis) | NeuNAcα2,3Galβ1,3(±NeuNAc2,6)GalNAc | - |
| PHA-E (Phaseolus vulgaris) | Galβ1,4GlcNAcβ1,2Man with bisecting GlcNAc | ++ |
| PHA-L (Phaseolus vulgaris) | Tri/tetraantennary complex type N-glycans w/terminal Gal | + |
| WGA (Triticum vulgaris) | GlcNAcβ1,4GlcNAc; chitin oligomers; NeuAc | ++ |
| RCA (Ricinus communis) | Galβ1,4GlcNAc | ++ |
| AAL (Aleuria aurantia) | Fucα1,6GlcNAc; Fucα1,3(Galβ1,4)GlcNAc | ++ |
| PHOSL (Pholiota squarrosa) | Fucα1,6GlcNAc | ++ |
| GSL-I (Griffonia simplicifolia) | Galα1,3Gal; Galα1,3GalNAc | + |
| LCA (Lens culinaris) | αDGlc, αDMan in N-glycans with Fuca1,6GlcNAc | ++ |
| DSL (Datura stramonium) | GlcNAcβ1,4GlcNAc oligomers; Galβ1,4GlcNAc | ++ |
| DFL (Narcissus pseudonarcissus) | α-linked mannose, preferring polymannose structures containing (α-1,6) linkages | + |
| GNL (Galanthus nivalis) | structures containing (α-1,3) mannose residues | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miljuš, G.; Penezić, A.; Pažitná, L.; Gligorijević, N.; Baralić, M.; Vilotić, A.; Šunderić, M.; Robajac, D.; Dobrijević, Z.; Katrlík, J.; et al. Glycosylation and Characterization of Human Transferrin in an End-Stage Kidney Disease. Int. J. Mol. Sci. 2024, 25, 4625. https://doi.org/10.3390/ijms25094625
Miljuš G, Penezić A, Pažitná L, Gligorijević N, Baralić M, Vilotić A, Šunderić M, Robajac D, Dobrijević Z, Katrlík J, et al. Glycosylation and Characterization of Human Transferrin in an End-Stage Kidney Disease. International Journal of Molecular Sciences. 2024; 25(9):4625. https://doi.org/10.3390/ijms25094625
Chicago/Turabian StyleMiljuš, Goran, Ana Penezić, Lucia Pažitná, Nikola Gligorijević, Marko Baralić, Aleksandra Vilotić, Miloš Šunderić, Dragana Robajac, Zorana Dobrijević, Jaroslav Katrlík, and et al. 2024. "Glycosylation and Characterization of Human Transferrin in an End-Stage Kidney Disease" International Journal of Molecular Sciences 25, no. 9: 4625. https://doi.org/10.3390/ijms25094625
APA StyleMiljuš, G., Penezić, A., Pažitná, L., Gligorijević, N., Baralić, M., Vilotić, A., Šunderić, M., Robajac, D., Dobrijević, Z., Katrlík, J., & Nedić, O. (2024). Glycosylation and Characterization of Human Transferrin in an End-Stage Kidney Disease. International Journal of Molecular Sciences, 25(9), 4625. https://doi.org/10.3390/ijms25094625