
Apoptosis in Postmortal Tissues of Goat Spinal Cords and Survival of Resident Neural Progenitors


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
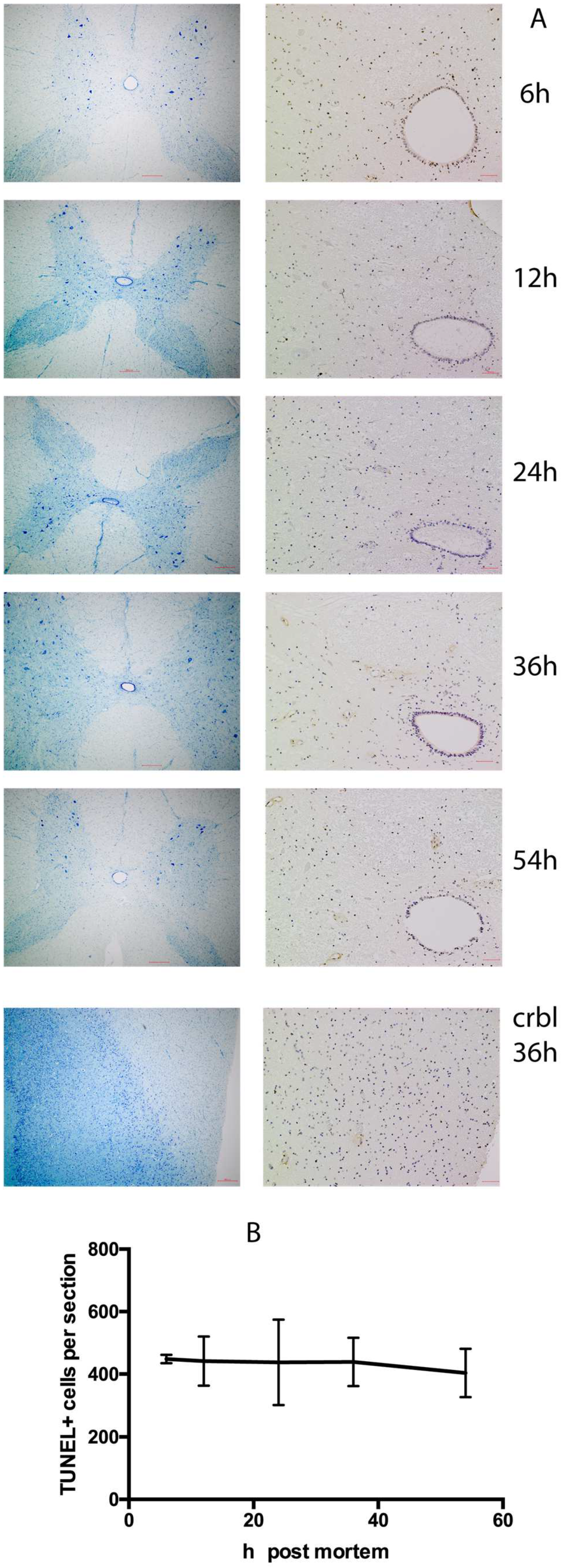
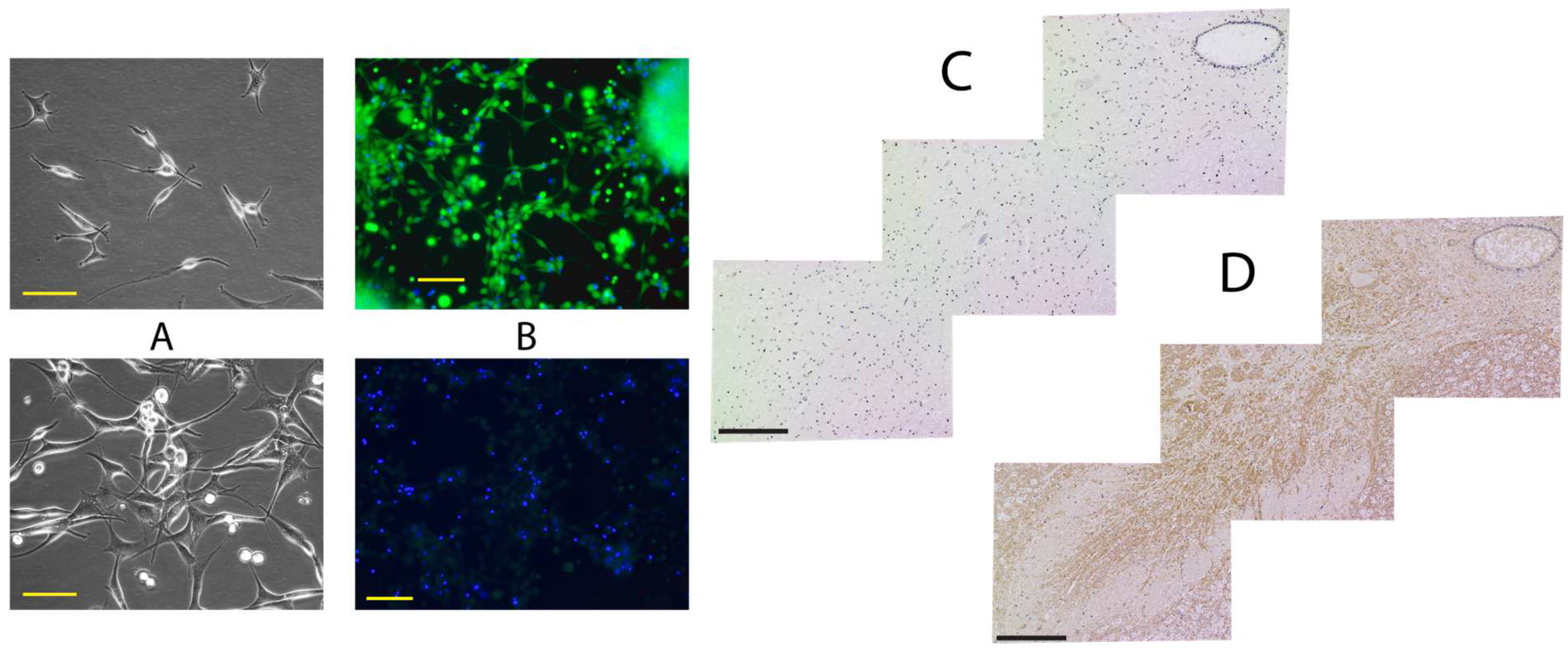
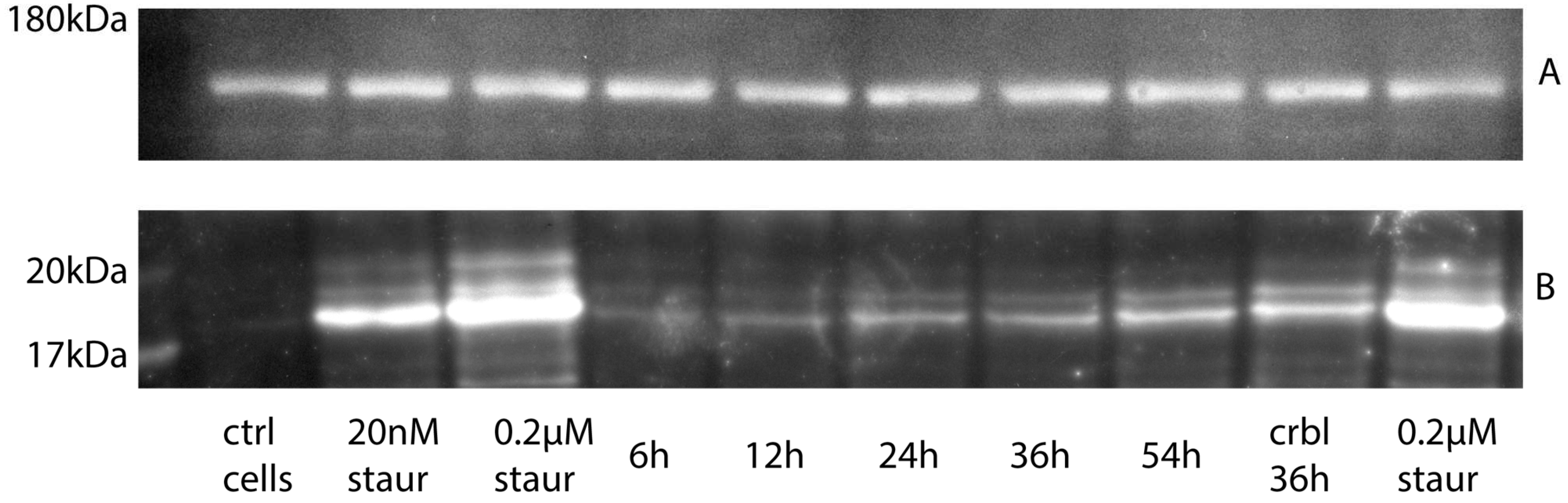
2. Results
2.1. Histological Analysis
2.2. Western Blot Analysis
2.3. Cytofluorometric Analysis
3. Discussion
4. Materials and Methods
4.1. Tissue Sampling
4.2. Histological Analysis
4.3. Cell Harvesting and Enrichment
4.4. Western Blot Analysis
4.5. Differentiation
4.6. Cytofluorometric Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| APC | allophycocyanin |
| CD24 | sialoglycoprotein antigen |
| CSCs | cadaveric stem cells |
| GD2 | disialoganglioside antigen |
| EDTA | ethylenediaminetetraacetic acid |
| FAM-VAD-FMK | fluorescent cell-permeable polycaspase inhibitor |
| FLICA | fluorochrome-labeled inhibitors of caspases |
| MSCs | mesenchymal stem cells |
| NSCs | neural stem cells |
| PVDF | polyvinylidene fluoride |
| PBS | phosphate buffer saline |
| TUNEL | terminal deoxynucleotidyl transferase mediated dUTP Nick End Labeling assay |
References
- Cieśla, J.; Tomsia, M. Cadaveric Stem Cells: Their Research Potential and Limitations. Front. Genet. 2021, 12, 798161. [Google Scholar] [CrossRef]
- Mansilla, E.; Mártire, K.; Roque, G.; Tau, J.M.; Marín, G.H.; Castuma, M.V.; Orlandi, G.; Tarditti, A. Salvage of Cadaver Stem Cells (CSCs) as a Routine Procedure: History or Future for Regenerative Medicine. J. Transplant. Technol. Res. 2013, 3, 118. [Google Scholar] [CrossRef]
- Sabrina, V.; Gianandrea, P. Phenotypic and Functional Mapping of Mesenchymal Stem Cells Harvested from Different Portions of the Human Arterial Tree. In Mesenchymal Stem Cells—Isolation, Characterization and Applications; InTech: Vienna, Austria, 2017. [Google Scholar] [CrossRef]
- Erker, L.; Azuma, H.; Lee, A.Y.; Guo, C.; Orloff, S.; Eaton, L.; Benedetti, E.; Jensen, B.; Finegold, M.; Willenbring, H.; et al. Therapeutic Liver Reconstitution With Murine Cells Isolated Long After Death. Gastroenterology 2010, 139, 1019–1029. [Google Scholar] [CrossRef]
- Mansilla, E.; Marín, G.H.; Berges, M.; Scafatti, S.; Rivas, J.; Núñez, A.; Menvielle, M.; Lamonega, R.; Gardiner, C.; Drago, H.; et al. Cadaveric bone marrow mesenchymal stem cells: First experience treating a patient with large severe burns. Burn. Trauma 2015, 3, 17. [Google Scholar] [CrossRef]
- Okonkwo, C.; Singh, M. Recovery of fibroblast-like cells from refrigerated goat skin up to 41 d of animal death. Vitr. Cell. Dev. Biol. Anim. 2015, 51, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Walcott, B.; Singh, M. Recovery of proliferative cells up to 15- and 49-day postmortem from bovine skin stored at 25 °C and 4 °C, respectively. Cogent Biol. 2017, 3, 1333760. [Google Scholar] [CrossRef]
- Alsook, M.K.S.; Gabriel, A.; Piret, J.; Waroux, O.; Tonus, C.; Connan, D.; Baise, E.; Antoine, N. Tissues from equine cadaver ligaments up to 72 hours of post-mortem: A promising reservoir of stem cells. Stem Cell Res. Ther. 2015, 6, 253. [Google Scholar] [CrossRef]
- Haring, G.; Zupan, J. Knee and Peri-Knee Tissues of Post Mortem Donors Are Strategic Sources of Mesenchymal Stem/Stromal Cells for Regenerative Procedures. Int. J. Mol. Sci. 2022, 23, 3170. [Google Scholar] [CrossRef]
- Dias, I.E.; Viegas, C.A.; Requicha, J.F.; Saavedra, M.J.; Azevedo, J.M.; Carvalho, P.P.; Dias, I.R. Mesenchymal Stem Cell Studies in the Goat Model for Biomedical Research. Biology 2022, 11, 1276. [Google Scholar] [CrossRef]
- Wright, A.; Arthaud-Day, M.L.; Weiss, M.L. Therapeutic Use of Mesenchymal Stromal Cells: The Need for Inclusive Characterization Guidelines to Accommodate All Tissue Sources and Species. Front. Cell Dev. Biol. 2021, 9, 632717. [Google Scholar] [CrossRef]
- Dunnett, S.B.; Björklund, A. Mechanisms and use of neural transplants for brain repair. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–51. [Google Scholar] [CrossRef]
- Schwartz, P.H.; Bryant, P.J.; Fuja, T.J.; Su, H.; O’Dowd, D.K.; Klassen, H. Isolation and characterization of neural progenitor cells from post-mortem human cortex. J. Neurosci. Res. 2003, 74, 838–851. [Google Scholar] [CrossRef] [PubMed]
- Moschella, M. Deconstructing the Brain Disconnection–Brain Death Analogy and Clarifying the Rationale for the Neurological Criterion of Death. J. Med. Philos. 2016, 41, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, T.; Hashizume, Y.; Sobue, G. Morphologic Features of the Normal Human Cadaveric Spinal Cord. Spine 1996, 21, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Berry, R.; Rodeheffer, M.S. Characterization of the adipocyte cellular lineage in vivo. Nat. Cell Biol. 2013, 15, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Garrett, S.H.; Sens, D.A.; Zhou, X.D.; Guyer, R.; Somji, S. Characterization and determination of cadmium resistance of CD133+/CD24+ and CD133−/CD24+ cells isolated from the immortalized human proximal tubule cell line, RPTEC/TERT1. Toxicol. Appl. Pharmacol. 2019, 375, 5–16. [Google Scholar] [CrossRef]
- Burmeister, D.M.; McIntyre, M.K.; Montgomery, R.K.; Gómez, B.I.; Dubick, M.A. Isolation and Characterization of Multipotent CD24+ Cells From the Renal Papilla of Swine. Front. Med. 2018, 5, 250. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Sui, X.; Zhang, D.; Liu, M.; Ding, M.; Shi, Y.; Deng, H. CD24: A Novel Surface Marker for PDX1-Positive Pancreatic Progenitors Derived from Human Embryonic Stem Cells. Stem Cells 2011, 29, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, C.; He, F.; Cai, Y.; Yang, H. Identification of CD44+CD24+ gastric cancer stem cells. J. Cancer Res. Clin. Oncol. 2011, 137, 1679–1686. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-J.; Lee, Y.-A.; Kim, K.-J.; Kim, Y.-H.; Jung, M.-S.; Ha, S.-J.; Kang, H.-G.; Jung, S.-E.; Kim, B.-G.; Choi, Y.-R.; et al. Effects of paracrine factors on CD24 expression and neural differentiation of male germline stem cells. Int. J. Mol. Med. 2015, 36, 255–262. [Google Scholar] [CrossRef]
- Pruszak, J.; Ludwig, W.; Blak, A.; Alavian, K.; Isacson, O. CD15, CD24, and CD29 Define a Surface Biomarker Code for Neural Lineage Differentiation of Stem Cells. Stem Cells 2009, 27, 2928–2940. [Google Scholar] [CrossRef]
- Pozhitkov, A.E.; Neme, R.; Domazet-Lošo, T.; Leroux, B.G.; Soni, S.; Tautz, D.; Noble, P.A. Tracing the dynamics of gene transcripts after organismal death. Open Biol. 2017, 7, 160267. [Google Scholar] [CrossRef] [PubMed]
- Ameisen, J.C. On the origin, evolution, and nature of programmed cell death: A timeline of four billion years. Cell Death Differ. 2002, 9, 367–393. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.-J.; Yang, B.-C.; Chen, J.-M.; Lin, Y.-H.; Huang, C.-L.; Cheng, Y.-Y.; Hsu, C.-Y.; Khoo, K.-H.; Shen, C.-N.; Yu, J. Changes in Glycosphingolipid Composition During Differentiation of Human Embryonic Stem Cells to Ectodermal or Endodermal Lineages. Stem Cells 2011, 29, 1995–2004. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Takamiya, K.; Furukawa, K. β1,4-N-acetylgalactosaminyltransferase—GM2/GD2 synthase: A key enzyme to control the synthesis of brain-enriched complex gangliosides. Biochim. Biophys. Acta—Gen. Subj. 2002, 1573, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Battula, V.L.; Shi, Y.; Evans, K.W.; Wang, R.-Y.; Spaeth, E.L.; Jacamo, R.O.; Guerra, R.; Sahin, A.A.; Marini, F.C.; Hortobagyi, G.; et al. Ganglioside GD2 identifies breast cancer stem cells and promotes tumorigenesis. J. Clin. Investig. 2012, 122, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Hosain, S.B.; Hill, R.A.; Liu, Y.-Y. The Role of Sphingolipids in Modulating Pluripotency of Stem Cells. In Trends in Stem Cell Proliferation and Cancer Research; Springer: Dordrecht, The Netherlands, 2013; pp. 167–191. [Google Scholar] [CrossRef]
- Madeja, Z.E.; Pawlak, P.; Piliszek, A. Beyond the mouse: Non-rodent animal models for study of early mammalian development and biomedical research. Int. J. Dev. Biol. 2019, 63, 187–201. [Google Scholar] [CrossRef]
- Llorente, V.; Velarde, P.; Desco, M.; Gómez-Gaviro, M.V. Current Understanding of the Neural Stem Cell Niches. Cells 2022, 11, 3002. [Google Scholar] [CrossRef]
- Paradiso, B.; Simonato, M.; Thiene, G.; Lavezzi, A. From fix to fit into the autoptic human brains. Eur. J. Histochem. 2018, 62, 257–265. [Google Scholar] [CrossRef]
- Mikhailov, A.; Sankai, Y. Donation of neural stem cells? Post mortal viability of spinal cord neuronal cells. In Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Honolulu, HI, USA, 18–21 July 2018; pp. 5333–5337. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikhailov, A.; Sankai, Y. Apoptosis in Postmortal Tissues of Goat Spinal Cords and Survival of Resident Neural Progenitors. Int. J. Mol. Sci. 2024, 25, 4683. https://doi.org/10.3390/ijms25094683
Mikhailov A, Sankai Y. Apoptosis in Postmortal Tissues of Goat Spinal Cords and Survival of Resident Neural Progenitors. International Journal of Molecular Sciences. 2024; 25(9):4683. https://doi.org/10.3390/ijms25094683
Chicago/Turabian StyleMikhailov, Andrey, and Yoshiyuki Sankai. 2024. "Apoptosis in Postmortal Tissues of Goat Spinal Cords and Survival of Resident Neural Progenitors" International Journal of Molecular Sciences 25, no. 9: 4683. https://doi.org/10.3390/ijms25094683
APA StyleMikhailov, A., & Sankai, Y. (2024). Apoptosis in Postmortal Tissues of Goat Spinal Cords and Survival of Resident Neural Progenitors. International Journal of Molecular Sciences, 25(9), 4683. https://doi.org/10.3390/ijms25094683