


Single-Cell Transcriptional Profile Construction of Rat Pituitary Glands before and after Sexual Maturation and Identification of Novel Marker Spp1 in Gonadotropes

 , and
, and {kind=link}
{kind=link}
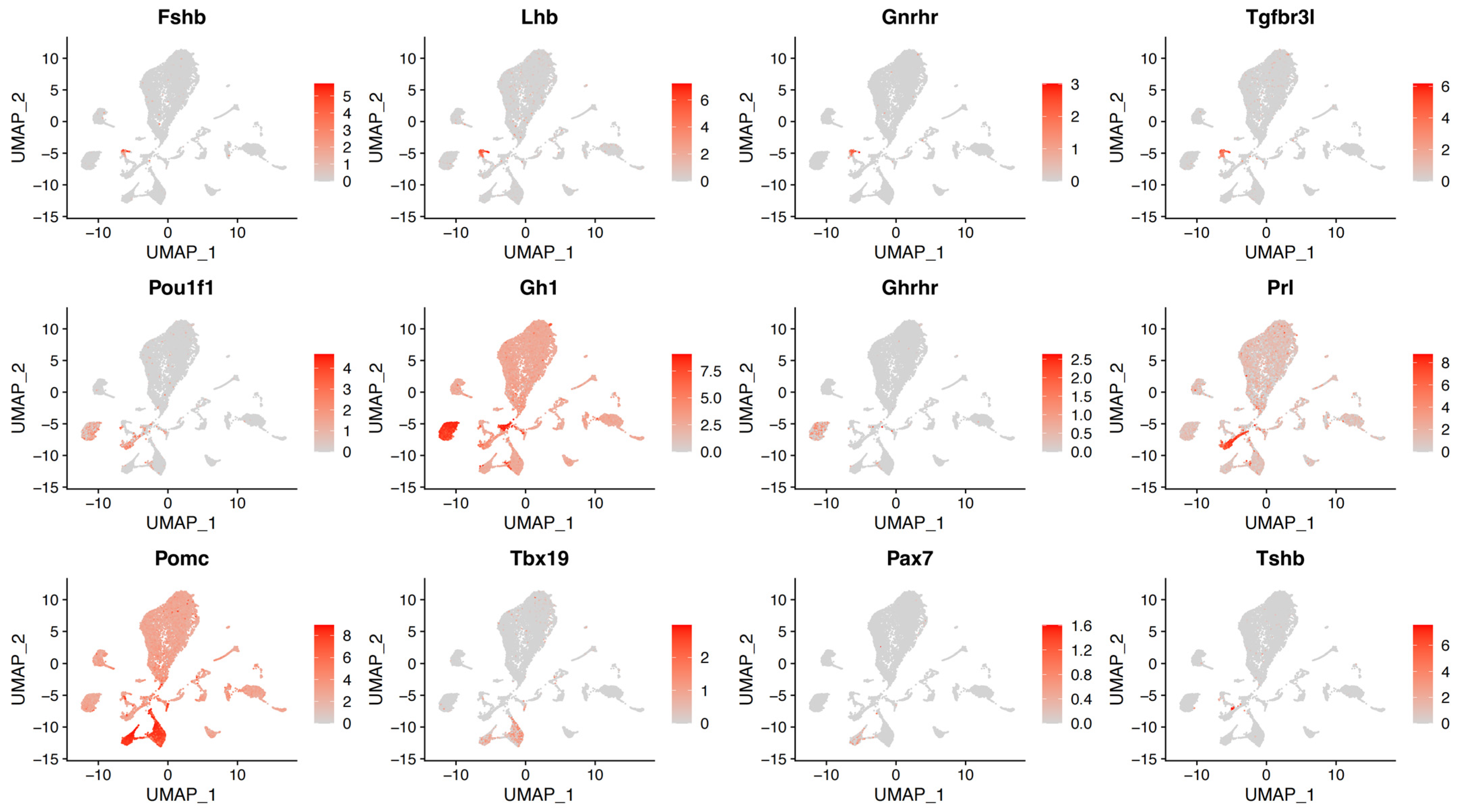{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Analysis of Pituitary Hormone Secretion Levels before and after Sexual Maturation in Rats
2.2. Construction of a Single-Cell Transcriptome Atlas of the Rat Pituitary Gland before and after Sexual Maturation
2.3. Enrichment Analysis of Differentially Expressed Genes in Gonadotrope Cells before and after Sexual Maturation
2.4. Validation of Differentially Expressed Genes in Different Cell Types
2.5. Identification and Validation of Spp1: A Novel Marker for Gonadotrope Cell Types’ Mining and Analysis of Novel Markers of Pituitary Hormone-Secreting Cells
3. Discussion
4. Materials and Methods
4.1. Animals and Cells
4.2. Cell Suspension Preparation
4.3. Cell Capture and RNA Library Construction
4.4. Bioinformatics Analysis of scRNA-seq Data
4.5. Cell Transfection
4.6. RT-qPCR
4.7. ELISA
4.8. Immunofluorescence
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropic hormone |
| ATP | Adeosine triphosphate |
| cAMP | Cyclic adenosine monophosphate |
| CREB | cAMP response element binding |
| FSH | Follicle-stimulating hormone |
| GH | growth hormone |
| GnRH | Gonadotropin-releasing hormone |
| LH | Luteinizing hormone |
| MAPK | Mitogen-activated protein kinase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| PKA | Protein kinase A |
| PRL | Prolactin |
| SPP1 | Secreted phosphoprotein 1 |
| TRβ | Thyroid hormone receptor β |
| TSH | Thyroid stimulating hormone |
References
- Alatzoglou, K.S.; Gregory, L.C.; Dattani, M.T. Development of the Pituitary Gland. Compr. Physiol. 2020, 10, 389–413. [Google Scholar] [CrossRef]
- Sheng, H.Z.; Moriyama, K.; Yamashita, T.; Li, H.; Potter, S.S.; Mahon, K.A.; Westphal, H. Multistep control of pituitary organogenesis. Science 1997, 278, 1809–1812. [Google Scholar] [CrossRef] [PubMed]
- Pottin, K.; Hyacinthe, C.; Retaux, S. Conservation, development, and function of a cement gland-like structure in the fish Astyanax mexicanus. Proc. Natl. Acad. Sci. USA 2010, 107, 17256–17261. [Google Scholar] [CrossRef]
- Kato, Y.; Yoshida, S.; Kato, T. Missing pieces of the pituitary puzzle: Participation of extra-adenohypophyseal placode-lineage cells in the adult pituitary gland. Cell Tissue Res. 2023, 394, 487–496. [Google Scholar] [CrossRef]
- Harris, J.; Gouhier, A.; Drouin, J. Mechanisms in endocrinology: Pioneer transcription factors in pituitary development and tumorigenesis. Eur. J. Endocrinol. 2021, 184, R1–R15. [Google Scholar] [CrossRef]
- Olson, L.E.; Tollkuhn, J.; Scafoglio, C.; Krones, A.; Zhang, J.; Ohgi, K.A.; Wu, W.; Taketo, M.M.; Kemler, R.; Grosschedl, R.; et al. Homeodomain-mediated beta-catenin-dependent switching events dictate cell-lineage determination. Cell 2006, 125, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Simmons, D.M.; Voss, J.W.; Ingraham, H.A.; Holloway, J.M.; Broide, R.S.; Rosenfeld, M.G.; Swanson, L.W. Pituitary cell phenotypes involve cell-specific Pit-1 mRNA translation and synergistic interactions with other classes of transcription factors. Genes Dev. 1990, 4, 695–711. [Google Scholar] [CrossRef] [PubMed]
- Lamolet, B.; Pulichino, A.M.; Lamonerie, T.; Gauthier, Y.; Brue, T.; Enjalbert, A.; Drouin, J. A pituitary cell-restricted T box factor, Tpit, activates POMC transcription in cooperation with Pitx homeoproteins. Cell 2001, 104, 849–859. [Google Scholar] [CrossRef]
- Ingraham, H.A.; Lala, D.S.; Ikeda, Y.; Luo, X.; Shen, W.H.; Nachtigal, M.W.; Abbud, R.; Nilson, J.H.; Parker, K.L. The nuclear receptor steroidogenic factor 1 acts at multiple levels of the reproductive axis. Genes Dev. 1994, 8, 2302–2312. [Google Scholar] [CrossRef]
- Ruf-Zamojski, F.; Fribourg, M.; Ge, Y.; Nair, V.; Pincas, H.; Zaslavsky, E.; Nudelman, G.; Tuminello, S.J.; Watanabe, H.; Turgeon, J.L.; et al. Regulatory Architecture of the LbetaT2 Gonadotrope Cell Underlying the Response to Gonadotropin-Releasing Hormone. Front. Endocrinol. 2018, 9, 34. [Google Scholar] [CrossRef]
- Mayran, A.; Sochodolsky, K.; Khetchoumian, K.; Harris, J.; Gauthier, Y.; Bemmo, A.; Balsalobre, A.; Drouin, J. Pioneer and nonpioneer factor cooperation drives lineage specific chromatin opening. Nat. Commun. 2019, 10, 3807. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, P.A.; Smiljanic, K.; Maso Previde, R.; Iben, J.R.; Li, T.; Rokic, M.B.; Sherman, A.; Coon, S.L.; Stojilkovic, S.S. Cell Type- and Sex-Dependent Transcriptome Profiles of Rat Anterior Pituitary Cells. Front. Endocrinol. 2019, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Cui, Y.; Ma, X.; Yong, J.; Yan, L.; Yang, M.; Ren, J.; Tang, F.; Wen, L.; Qiao, J. Single-cell transcriptomics identifies divergent developmental lineage trajectories during human pituitary development. Nat. Commun. 2020, 11, 5275. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, J.S.; Eugster, E.A. History of Puberty: Normal and Precocious. Horm. Res. Paediatr. 2022, 95, 568–578. [Google Scholar] [CrossRef]
- Calcaterra, V.; Rossi, V.; Massini, G.; Regalbuto, C.; Hruby, C.; Panelli, S.; Bandi, C.; Zuccotti, G. Precocious puberty and microbiota: The role of the sex hormone-gut microbiome axis. Front. Endocrinol. 2022, 13, 1000919. [Google Scholar] [CrossRef]
- Hou, H.; Chan, C.; Yuki, K.E.; Sokolowski, D.; Roy, A.; Qu, R.; Uuskula-Reimand, L.; Faykoo-Martinez, M.; Hudson, M.; Corre, C.; et al. Postnatal developmental trajectory of sex-biased gene expression in the mouse pituitary gland. Biol. Sex Differ. 2022, 13, 57. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Zhang, W.D.; Yuan, B.; Zhang, J.B. Advances in the Regulation of Mammalian Follicle-Stimulating Hormone Secretion. Animals 2021, 11, 1134. [Google Scholar] [CrossRef] [PubMed]
- Al-Samerria, S.; Radovick, S. Exploring the Therapeutic Potential of Targeting GH and IGF-1 in the Management of Obesity: Insights from the Interplay between These Hormones and Metabolism. Int. J. Mol. Sci. 2023, 24, 9556. [Google Scholar] [CrossRef]
- Bang, P. Pediatric Implications of Normal Insulin-GH-IGF Axis Physiology. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; South Dartmouth: Dartmouth, MA, USA, 2000. [Google Scholar]
- Hage, C.; Salvatori, R. Growth Hormone and Aging. Endocrinol. Metab. Clin. 2023, 52, 245–257. [Google Scholar] [CrossRef]
- Krassas, G.E.; Poppe, K.; Glinoer, D. Thyroid function and human reproductive health. Endocr. Rev. 2010, 31, 702–755. [Google Scholar] [CrossRef]
- Kakoulidis, I.; Ilias, I.; Stergiotis, S.; Togias, S.; Michou, A.; Lekkou, A.; Mastrodimou, V.; Pappa, A.; Milionis, C.; Venaki, E.; et al. Study on the Interaction between Serum Thyrotropin and Semen Parameters in Men. Med. Sci. 2022, 10, 22. [Google Scholar] [CrossRef]
- Atabaki, B.; Mirazi, N.; Hosseini, A.; Sarihi, A.; Izadi, Z.; Nourian, A. Effect of Elettaria cardamomum L. on hormonal changes and spermatogenesis in the propylthiouracil-induced hypothyroidism male BALB/c mice. Endocrinol. Diabetes Metab. 2023, 6, e438. [Google Scholar] [CrossRef]
- Krajewska-Kulak, E.; Sengupta, P. Thyroid function in male infertility. Front. Endocrinol. 2013, 4, 174. [Google Scholar] [CrossRef] [PubMed]
- Abeledo-Machado, A.; Pena-Zanoni, M.; Bornancini, D.; Diaz-Torga, G. Revealing Sexual Dimorphism in Prolactin Regulation From Early Postnatal Development to Adulthood in Murine Models. J. Endocr. Soc. 2023, 8, bvad146. [Google Scholar] [CrossRef] [PubMed]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, function, and regulation of secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef] [PubMed]
- Ladyman, S.R.; Hackwell, E.C.R.; Brown, R.S.E. The role of prolactin in co-ordinating fertility and metabolic adaptations during reproduction. Neuropharmacology 2020, 167, 107911. [Google Scholar] [CrossRef]
- Becu-Villalobos, D.; Lacau-Mengido, I.M.; Diaz-Torga, G.S.; Libertun, C. Ontogenic studies of the neural control of adenohypophyseal hormones in the rat. II. Prolactin. Cell. Mol. Neurobiol. 1992, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Johnston, H.; King, P.J.; O’Shaughnessy, P.J. Effects of ACTH and expression of the melanocortin-2 receptor in the neonatal mouse testis. Reproduction 2007, 133, 1181–1187. [Google Scholar] [CrossRef]
- Hasenmajer, V.; Bonaventura, I.; Minnetti, M.; Sada, V.; Sbardella, E.; Isidori, A.M. Non-Canonical Effects of ACTH: Insights Into Adrenal Insufficiency. Front. Endocrinol. 2021, 12, 701263. [Google Scholar] [CrossRef]
- Brandt, Y.; Lang, A.; Madej, A.; Rodriguez-Martinez, H.; Einarsson, S. Impact of ACTH administration on the oviductal sperm reservoir in sows: The local endocrine environment and distribution of spermatozoa. Anim. Reprod. Sci. 2006, 92, 107–122. [Google Scholar] [CrossRef]
- Brandt, Y.; Lang, A.; Rodriguez-Martinez, H.; Madej, A.; Einarsson, S. Impact of ACTH during oestrus on the ultrastructure of the spermatozoa and their environment in the tubal reservoir of the postovulatory sow. Anim. Reprod. Sci. 2006, 93, 231–245. [Google Scholar] [CrossRef]
- Cheung, L.Y.M.; George, A.S.; McGee, S.R.; Daly, A.Z.; Brinkmeier, M.L.; Ellsworth, B.S.; Camper, S.A. Single-Cell RNA Sequencing Reveals Novel Markers of Male Pituitary Stem Cells and Hormone-Producing Cell Types. Endocrinology 2018, 159, 3910–3924. [Google Scholar] [CrossRef]
- Ho, Y.; Hu, P.; Peel, M.T.; Chen, S.; Camara, P.G.; Epstein, D.J.; Wu, H.; Liebhaber, S.A. Single-cell transcriptomic analysis of adult mouse pituitary reveals sexual dimorphism and physiologic demand-induced cellular plasticity. Protein Cell 2020, 11, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Bosch, E.; Alviggi, C.; Lispi, M.; Conforti, A.; Hanyaloglu, A.C.; Chuderland, D.; Simoni, M.; Raine-Fenning, N.; Crepieux, P.; Kol, S.; et al. Reduced FSH and LH action: Implications for medically assisted reproduction. Hum. Reprod. 2021, 36, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.R.; Ciccone, N.A.; Xu, S.; Zaytseva, S.; Carroll, R.S.; Kaiser, U.B. GnRH pulse frequency-dependent stimulation of FSHbeta transcription is mediated via activation of PKA and CREB. Mol. Endocrinol. 2013, 27, 606–618. [Google Scholar] [CrossRef]
- Ehrchen, J.; Heuer, H.; Sigmund, R.; Schafer, M.K.; Bauer, K. Expression and regulation of osteopontin and connective tissue growth factor transcripts in rat anterior pituitary. J. Endocrinol. 2001, 169, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Bjelobaba, I.; Janjic, M.M.; Previde, R.M.; Abebe, D.; Kucka, M.; Stojilkovic, S.S. Distinct Expression Patterns of Osteopontin and Dentin Matrix Protein 1 Genes in Pituitary Gonadotrophs. Front. Endocrinol. 2019, 10, 248. [Google Scholar] [CrossRef]
- Liu, K.; Hu, H.; Jiang, H.; Zhang, H.; Gong, S.; Wei, D.; Yu, Z. RUNX1 promotes MAPK signaling to increase tumor progression and metastasis via OPN in head and neck cancer. Carcinogenesis 2021, 42, 414–422. [Google Scholar] [CrossRef]
- Che, C.; Liu, J.; Yang, J.; Ma, L.; Bai, N.; Zhang, Q. Osteopontin is essential for IL-1beta production and apoptosis in peri-implantitis. Clin. Implant. Dent. Relat. Res. 2018, 20, 384–392. [Google Scholar] [CrossRef]
- Meller, R.; Stevens, S.L.; Minami, M.; Cameron, J.A.; King, S.; Rosenzweig, H.; Doyle, K.; Lessov, N.S.; Simon, R.P.; Stenzel-Poore, M.P. Neuroprotection by osteopontin in stroke. J. Cereb. Blood Flow Metab. 2005, 25, 217–225. [Google Scholar] [CrossRef]
- Han, H.; Ge, X.; Komakula, S.S.B.; Desert, R.; Das, S.; Song, Z.; Chen, W.; Athavale, D.; Gaskell, H.; Lantvit, D.; et al. Macrophage-derived Osteopontin (SPP1) Protects From Nonalcoholic Steatohepatitis. Gastroenterology 2023, 165, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Wen, W.; Ye, S. TSH-SPP1/TRbeta-TSH positive feedback loop mediates fat deposition of hepatocyte: Crosstalk between thyroid and liver. Front. Immunol. 2022, 13, 1009912. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ramos, R.; Phan, A.Q.; Yamaga, K.; Flesher, J.L.; Jiang, S.; Oh, J.W.; Jin, S.; Jahid, S.; Kuan, C.H.; et al. Signalling by senescent melanocytes hyperactivates hair growth. Nature 2023, 618, 808–817. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Zeng, F.; Su, J.; Zhao, S.; Hu, R.; Zhu, W.; Hu, S.; Chen, X.; Yin, M. BET inhibitor suppresses melanoma progression via the noncanonical NF-kappaB/SPP1 pathway. Theranostics 2020, 10, 11428–11443. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Fan, Y.; Zhang, Y.; Yan, X.; Li, W.; Yan, X.; He, L.; Wang, N.; Chen, O.; He, D.; et al. The regulation mechanism of different hair types in inner Mongolia cashmere goat based on PI3K-AKT pathway and FGF21. J. Anim. Sci. 2022, 100, skac292. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Tan, X.; Yuan, G.; Tang, Y.; Wang, Y.; Yang, C.; Luo, S.; Wu, Z.; Yao, K. SPP1 is associated with adverse prognosis and predicts immunotherapy efficacy in penile cancer. Hum. Genom. 2023, 17, 116. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Liu, D.; Sun, H.; Shao, Z.; Shi, P.; Li, T.; Yin, S.; Zhu, T. SPP1 is a prognostic related biomarker and correlated with tumor-infiltrating immune cells in ovarian cancer. BMC Cancer 2022, 22, 1367. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Guo, J.; Ai, L.; Wang, Y.; Wang, Y.; Chen, B.; Liu, M.; Zhuang, S.; Liu, K.; Zhao, Z.; et al. Up-regulated SPP1 increases the risk from IPF to lung cancer via activating the pro-tumor macrophages. Comput. Struct. Biotechnol. J. 2023, 21, 5751–5764. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Shi, Y.; Meng, L.; Ma, J.; Huang, S.; Zhang, J.; Wu, Y.; Li, J.; Lin, Y.; Yang, S.; et al. Publisher Correction: Single-cell transcriptomic analysis suggests two molecularly distinct subtypes of intrahepatic cholangiocarcinoma. Nat. Commun. 2022, 13, 2848. [Google Scholar] [CrossRef]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587.E29. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Q.-H.; Zhao, G.-K.; Wang, H.-Q.; Wei, F.-H.; Zhang, J.-Y.; Zhang, J.-B.; Gao, F.; Yuan, B. Single-Cell Transcriptional Profile Construction of Rat Pituitary Glands before and after Sexual Maturation and Identification of Novel Marker Spp1 in Gonadotropes. Int. J. Mol. Sci. 2024, 25, 4694. https://doi.org/10.3390/ijms25094694
Huang Q-H, Zhao G-K, Wang H-Q, Wei F-H, Zhang J-Y, Zhang J-B, Gao F, Yuan B. Single-Cell Transcriptional Profile Construction of Rat Pituitary Glands before and after Sexual Maturation and Identification of Novel Marker Spp1 in Gonadotropes. International Journal of Molecular Sciences. 2024; 25(9):4694. https://doi.org/10.3390/ijms25094694
Chicago/Turabian StyleHuang, Qing-Hua, Guo-Kun Zhao, Hao-Qi Wang, Fan-Hao Wei, Jin-Yu Zhang, Jia-Bao Zhang, Fei Gao, and Bao Yuan. 2024. "Single-Cell Transcriptional Profile Construction of Rat Pituitary Glands before and after Sexual Maturation and Identification of Novel Marker Spp1 in Gonadotropes" International Journal of Molecular Sciences 25, no. 9: 4694. https://doi.org/10.3390/ijms25094694
APA StyleHuang, Q. -H., Zhao, G. -K., Wang, H. -Q., Wei, F. -H., Zhang, J. -Y., Zhang, J. -B., Gao, F., & Yuan, B. (2024). Single-Cell Transcriptional Profile Construction of Rat Pituitary Glands before and after Sexual Maturation and Identification of Novel Marker Spp1 in Gonadotropes. International Journal of Molecular Sciences, 25(9), 4694. https://doi.org/10.3390/ijms25094694