


ERECTA Modulates Seed Germination and Fruit Development via Auxin Signaling in Tomato

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. sler Exhibited Compact Traits
2.2. sler Altered Fruit and Seed Formation
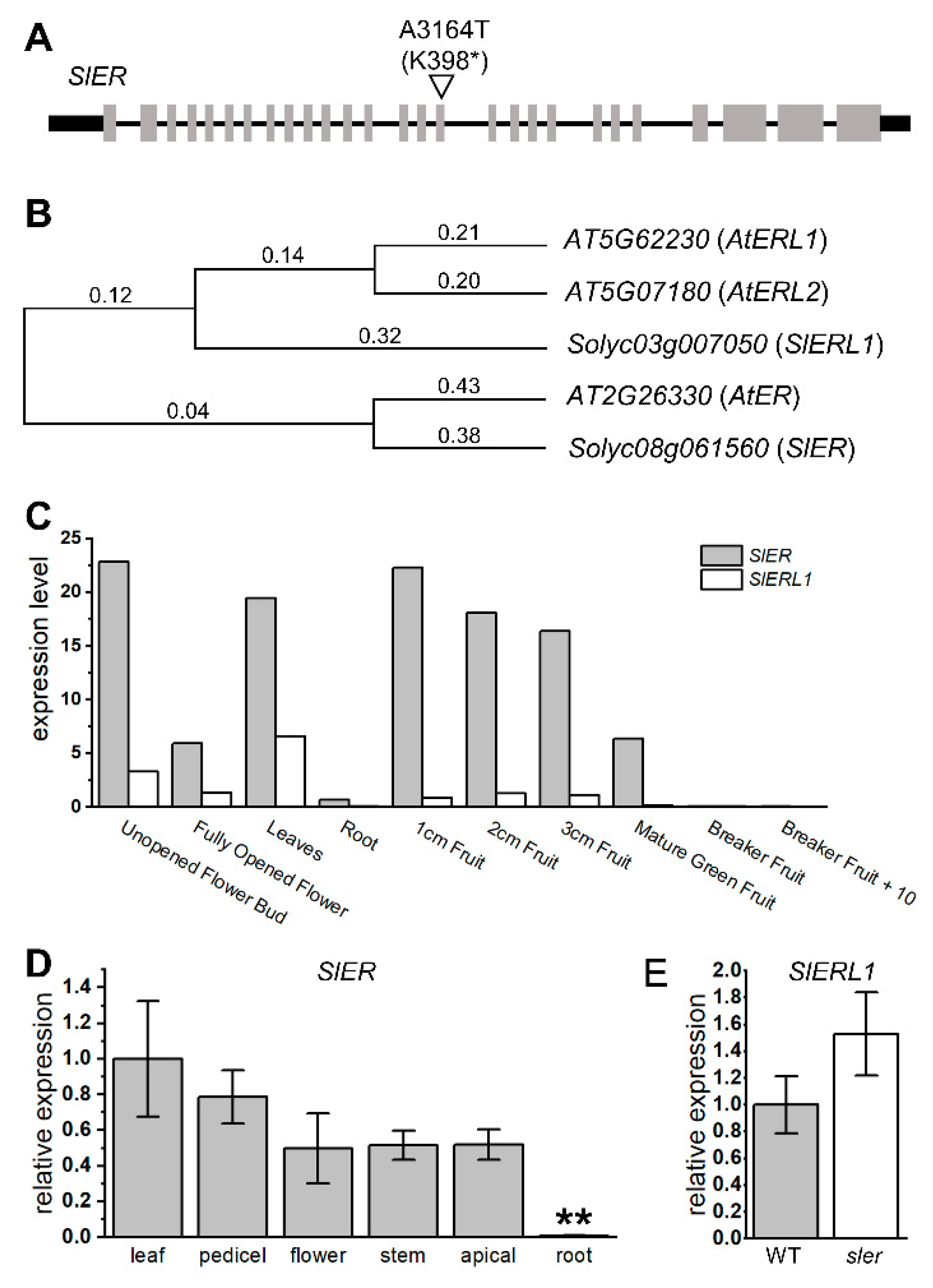
2.3. Genetic Redundancy between SlER and AtER
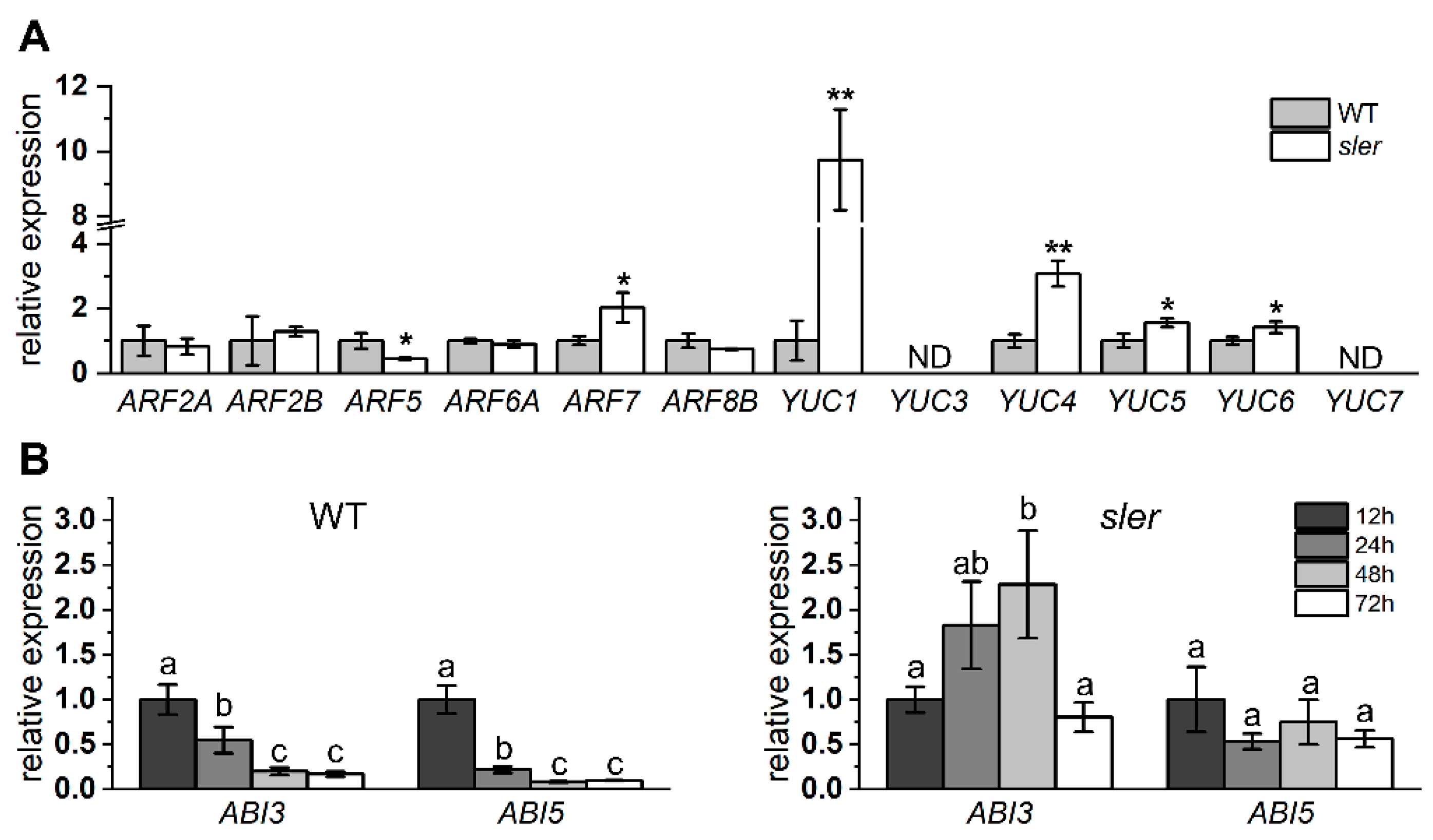
2.4. Enhanced Auxin Signal in the Seed and Ovary
3. Discussion
3.1. Gene Duplication and Functional Divergence of ERECTA
3.2. ERECTA Regulates Seed Germination via Auxin Signaling
3.3. ERECTA Regulates Fruit Set
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Phenotypic Measurement
4.3. Seed Germination Test
4.4. Aniline Blue Staining of Pollen Tubes
4.5. DNA Extraction and Sequencing
4.6. Comparative Analysis of ER Family Genes
4.7. Immunolocalization of IAA
4.8. RNA Extraction and qRT-PCR
4.9. Statistical Method
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed Dormancy and the Control of Germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Graeber, K.; Linkies, A.; Müller, K.; Wunchova, A.; Rott, A.; Leubner-Metzger, G. Cross-Species Approaches to Seed Dormancy and Germination: Conservation and Biodiversity of ABA-Regulated Mechanisms and the Brassicaceae DOG1 Genes. Plant Mol. Biol. 2010, 73, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Voegele, A.; Linkies, A.; Müller, K.; Leubner-Metzger, G. Members of the Gibberellin Receptor Gene Family GID1 (GIBBERELLIN INSENSITIVE DWARF1) Play Distinct Roles during Lepidium Sativum and Arabidopsis Thaliana Seed Germination. J. Exp. Bot. 2011, 62, 5131–5147. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.-Q.; Luan, S.; Li, J.; He, Z.-H. Auxin Controls Seed Dormancy through Stimulation of Abscisic Acid Signaling by Inducing ARF-Mediated ABI3 Activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Mei, S.; Zhang, M.; Ye, J.; Du, J.; Jiang, Y.; Hu, Y. Auxin Contributes to Jasmonate-Mediated Regulation of Abscisic Acid Signaling during Seed Germination in Arabidopsis. Plant Cell 2023, 35, 1110–1133. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Luo, X.; Li, M.; Joldersma, D.; Plunkert, M.; Liu, Z. Mechanism of Fertilization-Induced Auxin Synthesis in the Endosperm for Seed and Fruit Development. Nat. Commun. 2022, 13, 3985. [Google Scholar] [CrossRef]
- Jensen, P.J.; Bandurski, R.S. Metabolism and Synthesis of Indole-3-Acetic Acid (IAA) in Zea Mays (Levels of IAA during Kernel Development and the Use of in Vitro Endosperm Systems for Studying IAA Biosynthesis). Plant Physiol. 1994, 106, 343–351. [Google Scholar] [CrossRef]
- Yue, J.; Hu, X.; Huang, J. Origin of Plant Auxin Biosynthesis. Trends Plant Sci. 2014, 19, 764–770. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, Z.; Lei, C.; Zheng, C.; Wang, J.; Shao, S.; Li, X.; Xia, X.; Cai, X.; Zhou, J.; et al. A Plant Phytosulfokine Peptide Initiates Auxin-Dependent Immunity through Cytosolic Ca2+ Signaling in Tomato. Plant Cell 2018, 30, 652–667. [Google Scholar] [CrossRef]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Gene Structure and Spatiotemporal Expression Profile of Tomato Genes Encoding YUCCA-like Flavin Monooxygenases: The ToFZY Gene Family. Plant Physiol. Biochem. 2011, 49, 782–791. [Google Scholar] [CrossRef]
- Chandler, J.W. Auxin Response Factors. Plant Cell Environ. 2016, 39, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Tyagi, A.K.; Sharma, A.K. Genome-Wide Analysis of Auxin Response Factor (ARF) Gene Family from Tomato and Analysis of Their Role in Flower and Fruit Development. Mol. Genet. Genom. 2011, 285, 245–260. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.; Mariani, C.; Vriezen, W.H. The Role of Auxin and Gibberellin in Tomato Fruit Set. J. Exp. Bot. 2009, 60, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Pandolfini, T.; Rotino, G.L.; Camerini, S.; Defez, R.; Spena, A. Optimisation of Transgene Action at the Post-Transcriptional Level: High Quality Parthenocarpic Fruits in Industrial Tomatoes. BMC Biotechnol. 2002, 2, 1. [Google Scholar] [CrossRef] [PubMed]
- Mazzucato, A.; Cellini, F.; Bouzayen, M.; Zouine, M.; Mila, I.; Minoia, S.; Petrozza, A.; Picarella, M.E.; Ruiu, F.; Carriero, F. A TILLING Allele of the Tomato Aux/IAA9 Gene Offers New Insights into Fruit Set Mechanisms and Perspectives for Breeding Seedless Tomatoes. Mol. Breed. 2015, 35, 22. [Google Scholar] [CrossRef]
- Hu, J.; Israeli, A.; Ori, N.; Sun, T. The Interaction between DELLA and ARF/IAA Mediates Crosstalk between Gibberellin and Auxin Signaling to Control Fruit Initiation in Tomato. Plant Cell 2018, 30, 1710–1728. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Li, X.; Sun, T. Four Class A AUXIN RESPONSE FACTORs Promote Tomato Fruit Growth despite Suppressing Fruit Set. Nat. Plants 2023, 9, 706–719. [Google Scholar] [CrossRef]
- Varoquaux, F.; Blanvillain, R.; Delseny, M.; Gallois, P. Less Is Better: New Approaches for Seedless Fruit Production. Trends Biotechnol. 2000, 18, 233–242. [Google Scholar] [CrossRef]
- Santos, D.D.; Yamaguchi, M. Seed-Sprouting in Tomato Fruits. Sci. Hortic. 1979, 11, 131–139. [Google Scholar] [CrossRef]
- Zhang, M.; Yuan, B.; Leng, P. The Role of ABA in Triggering Ethylene Biosynthesis and Ripening of Tomato Fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef]
- Kohalmi, S.E.; Kunz, B.A. Role of Neighbouring Bases and Assessment of Strand Specificity in Ethylmethanesulphonate and N-Methyl-N′-Nitro-N-Nitrosoguanidine Mutagenesis in the SUP4-o Gene of Saccharomyces Cerevisiae. J. Mol. Biol. 1988, 204, 561–568. [Google Scholar] [CrossRef]
- Henikoff, S.; Till, B.J.; Comai, L. TILLING. Traditional Mutagenesis Meets Functional Genomics. Plant Physiol. 2004, 135, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Mizoguchi, T.; Aoki, K.; Kubo, Y.; Mori, H.; Imanishi, S.; Yamazaki, Y.; Shibata, D.; Ezura, H. Ethylmethanesulfonate (EMS) mutagenesis of Solanum lycopersicum cv. Micro-Tom for large-scale mutant screens. Plant Biotechnol. 2007, 24, 33–38. [Google Scholar] [CrossRef]
- Meng, X.; Chen, X.; Mang, H.; Liu, C.; Yu, X.; Gao, X.; Torii, K.U.; He, P.; Shan, L. Differential Function of Arabidopsis SERK Family Receptor-like Kinases in Stomatal Patterning. Curr. Biol. 2015, 25, 2361–2372. [Google Scholar] [CrossRef]
- Zhang, L.; DeGennaro, D.; Lin, G.; Chai, J.; Shpak, E.D. ERECTA Family Signaling Constrains CLAVATA3 and WUSCHEL to the Center of the Shoot Apical Meristem. Development 2021, 148, dev189753. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Huang, Y.; Liu, L.; Zhang, M.; Chai, M.; Xi, X.; Aslam, M.; Wang, L.; Ma, S.; Su, H.; et al. Signaling by the EPFL-ERECTA Family Coordinates Female Germline Specification through the BZR1 Family in Arabidopsis. Plant Cell 2023, 35, 1455–1473. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, X.; Zhang, B.; Wu, S.; Wang, R.; Li, N.; Li, Y.; Sun, Y.; Tang, W. ERECTA Regulates Seed Size Independently of Its Intracellular Domain via MAPK-DA1-UBP15 Signaling. Plant Cell 2022, 34, 3773–3789. [Google Scholar] [CrossRef]
- Villagarcia, H.; Morin, A.-C.; Shpak, E.D.; Khodakovskaya, M.V. Modification of Tomato Growth by Expression of Truncated ERECTA Protein from Arabidopsis Thaliana. J. Exp. Bot. 2012, 63, 6493–6504. [Google Scholar] [CrossRef]
- Kwon, C.-T.; Heo, J.; Lemmon, Z.H.; Capua, Y.; Hutton, S.F.; Van Eck, J.; Park, S.J.; Lippman, Z.B. Rapid Customization of Solanaceae Fruit Crops for Urban Agriculture. Nat. Biotechnol. 2020, 38, 182–188. [Google Scholar] [CrossRef]
- Shen, H.; Zhong, X.; Zhao, F.; Wang, Y.; Yan, B.; Li, Q.; Chen, G.; Mao, B.; Wang, J.; Li, Y.; et al. Overexpression of Receptor-like Kinase ERECTA Improves Thermotolerance in Rice and Tomato. Nat. Biotechnol. 2015, 33, 996–1003. [Google Scholar] [CrossRef]
- Nanda, A.K.; El Habti, A.; Hocart, C.H.; Masle, J. ERECTA Receptor-Kinases Play a Key Role in the Appropriate Timing of Seed Germination under Changing Salinity. J. Exp. Bot. 2019, 70, 6417–6435. [Google Scholar] [CrossRef]
- Qu, X.; Zhao, Z.; Tian, Z. ERECTA Regulates Cell Elongation by Activating Auxin Biosynthesis in Arabidopsis Thaliana. Front. Plant Sci. 2017, 8, 298299. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-K.; Wilson, R.L.; Palme, K.; Ditengou, F.A.; Shpak, E.D. ERECTA Family Genes Regulate Auxin Transport in the Shoot Apical Meristem and Forming Leaf Primordia. Plant Physiol. 2013, 162, 1978–1991. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Chai, M.; Chen, F.; Huang, Y.; Zhang, M.; He, Q.; Liu, L.; Yan, M.; Qin, Y. HBI1 Acts Downstream of ERECTA and SWR1 in Regulating Inflorescence Architecture through the Activation of the Brassinosteroid and Auxin Signaling Pathways. New Phytol. 2021, 229, 414–428. [Google Scholar] [CrossRef] [PubMed]
- Parre, E.; Geitmann, A. More Than a Leak Sealant. The Mechanical Properties of Callose in Pollen Tubes. Plant Physiol. 2005, 137, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Raul, A.; Bernard, L. Imaging Callose at Plasmodesmata Using Aniline Blue: Quantitative Confocal Microscopy|SpringerLink. Available online: https://link.springer.com/protocol/10.1007/978-1-4939-1523-1_7 (accessed on 9 April 2024).
- Birchler, J.A.; Yang, H. The Multiple Fates of Gene Duplications: Deletion, Hypofunctionalization, Subfunctionalization, Neofunctionalization, Dosage Balance Constraints, and Neutral Variation. Plant Cell 2022, 34, 2466–2474. [Google Scholar] [CrossRef] [PubMed]
- Shpak, E.D.; Berthiaume, C.T.; Hill, E.J.; Torii, K.U. Synergistic Interaction of Three ERECTA-Family Receptor-like Kinases Controls Arabidopsis Organ Growth and Flower Development by Promoting Cell Proliferation. Development 2004, 131, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Liu, L.; Ma, S.; Aslam, M.; Qin, Y. Insights into the Role of Phytohormones in Plant Female Germline Cell Specification. Curr. Opin. Plant Biol. 2023, 75, 102439. [Google Scholar] [CrossRef] [PubMed]
- Leng, Y.-J.; Yao, Y.-S.; Yang, K.-Z.; Wu, P.-X.; Xia, Y.-X.; Zuo, C.-R.; Luo, J.-H.; Wang, P.; Liu, Y.-Y.; Zhang, X.-Q.; et al. Arabidopsis ERdj3B Coordinates with ERECTA-Family Receptor Kinases to Regulate Ovule Development and the Heat Stress Response. Plant Cell 2022, 34, 3665–3684. [Google Scholar] [CrossRef]
- Hu, Y.; Lin, Y.; Bai, J.; Xu, X.; Wang, Z.; Ding, C.; Ding, Y.; Chen, L. AMPK Activator 991 Specifically Activates SnRK1 and Thereby Affects Seed Germination in Rice. J. Exp. Bot. 2024, erae046. [Google Scholar] [CrossRef]
- Wang, Z.; Ren, Z.; Cheng, C.; Wang, T.; Ji, H.; Zhao, Y.; Deng, Z.; Zhi, L.; Lu, J.; Wu, X.; et al. Counteraction of ABA-Mediated Inhibition of Seed Germination and Seedling Establishment by ABA Signaling Terminator in Arabidopsis. Mol. Plant 2020, 13, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wu, J.; Gan, Y. The New Insight of Auxin Functions: Transition from Seed Dormancy to Germination and Floral Opening in Plants. Plant Growth Regul. 2020, 91, 169–174. [Google Scholar] [CrossRef]
- Xu, X.; E, Z.; Zhang, D.; Yun, Q.; Zhou, Y.; Niu, B.; Chen, C. OsYUC11-Mediated Auxin Biosynthesis Is Essential for Endosperm Development of Rice. Plant Physiol. 2021, 185, 934–950. [Google Scholar] [CrossRef] [PubMed]
- Ensslin, A.; Van de Vyver, A.; Vanderborght, T.; Godefroid, S. Ex Situ Cultivation Entails High Risk of Seed Dormancy Loss on Short-Lived Wild Plant Species. J. Appl. Ecol. 2018, 55, 1145–1154. [Google Scholar] [CrossRef]
- Hensel, L.; Grbić, V.; Baumgarten, D.; Bleecker, A. Developmental and Age-Related Processes That Influence the Longevity and Senescence of Photosynthetic Tissues in Arabidopsis.|The Plant Cell|Oxford Academic. Available online: https://academic.oup.com/plcell/article/5/5/553/5984515?login=true (accessed on 12 April 2024).
- Mori, K.; Lemaire-Chamley, M.; Jorly, J.; Carrari, F.; Conte, M.; Asamizu, E.; Mizoguchi, T.; Ezura, H.; Rothan, C. The Conserved Brassinosteroid-Related Transcription Factor BIM1a Negatively Regulates Fruit Growth in Tomato. J. Exp. Bot. 2021, 72, 1181–1197. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Jiang, C.; Liu, Y.; Gao, Y.; Lu, J.; Aiwaili, P.; Fei, Z.; Jiang, C.-Z.; Hong, B.; Ma, C.; et al. Auxin Regulates Sucrose Transport to Repress Petal Abscission in Rose (Rosa hybrida). Plant Cell 2020, 32, 3485–3499. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Xu, Y.; Li, J.; Shiba, H.; Ezura, H.; Wang, N. ERECTA Modulates Seed Germination and Fruit Development via Auxin Signaling in Tomato. Int. J. Mol. Sci. 2024, 25, 4754. https://doi.org/10.3390/ijms25094754
Chen D, Xu Y, Li J, Shiba H, Ezura H, Wang N. ERECTA Modulates Seed Germination and Fruit Development via Auxin Signaling in Tomato. International Journal of Molecular Sciences. 2024; 25(9):4754. https://doi.org/10.3390/ijms25094754
Chicago/Turabian StyleChen, Daoyun, Yuqing Xu, Jiawei Li, Hiroshi Shiba, Hiroshi Ezura, and Ning Wang. 2024. "ERECTA Modulates Seed Germination and Fruit Development via Auxin Signaling in Tomato" International Journal of Molecular Sciences 25, no. 9: 4754. https://doi.org/10.3390/ijms25094754
APA StyleChen, D., Xu, Y., Li, J., Shiba, H., Ezura, H., & Wang, N. (2024). ERECTA Modulates Seed Germination and Fruit Development via Auxin Signaling in Tomato. International Journal of Molecular Sciences, 25(9), 4754. https://doi.org/10.3390/ijms25094754