Selenium Derivatization of Nucleic Acids for Phase and Structure Determination in Nucleic Acid X-ray Crystallography

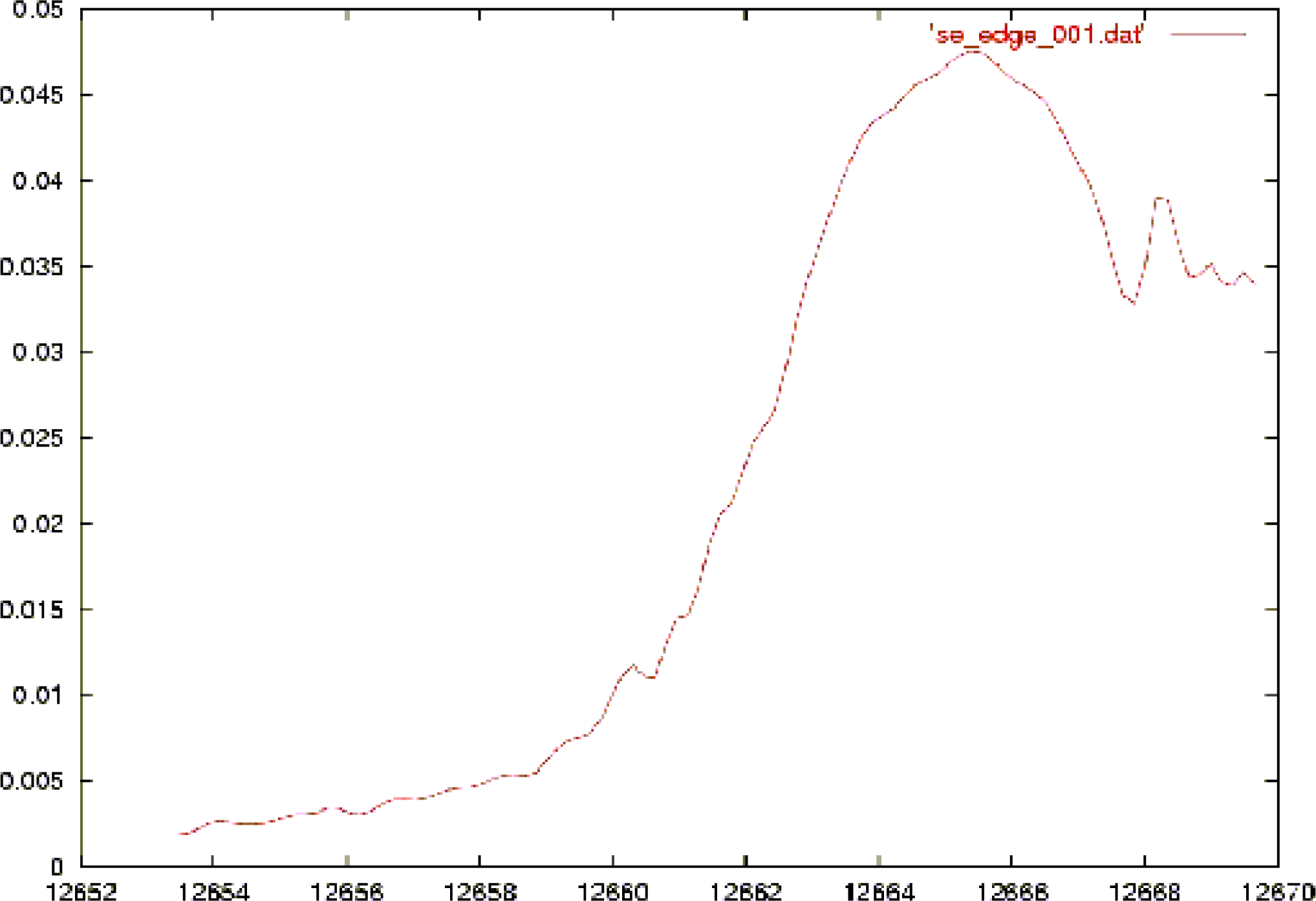{kind=link}
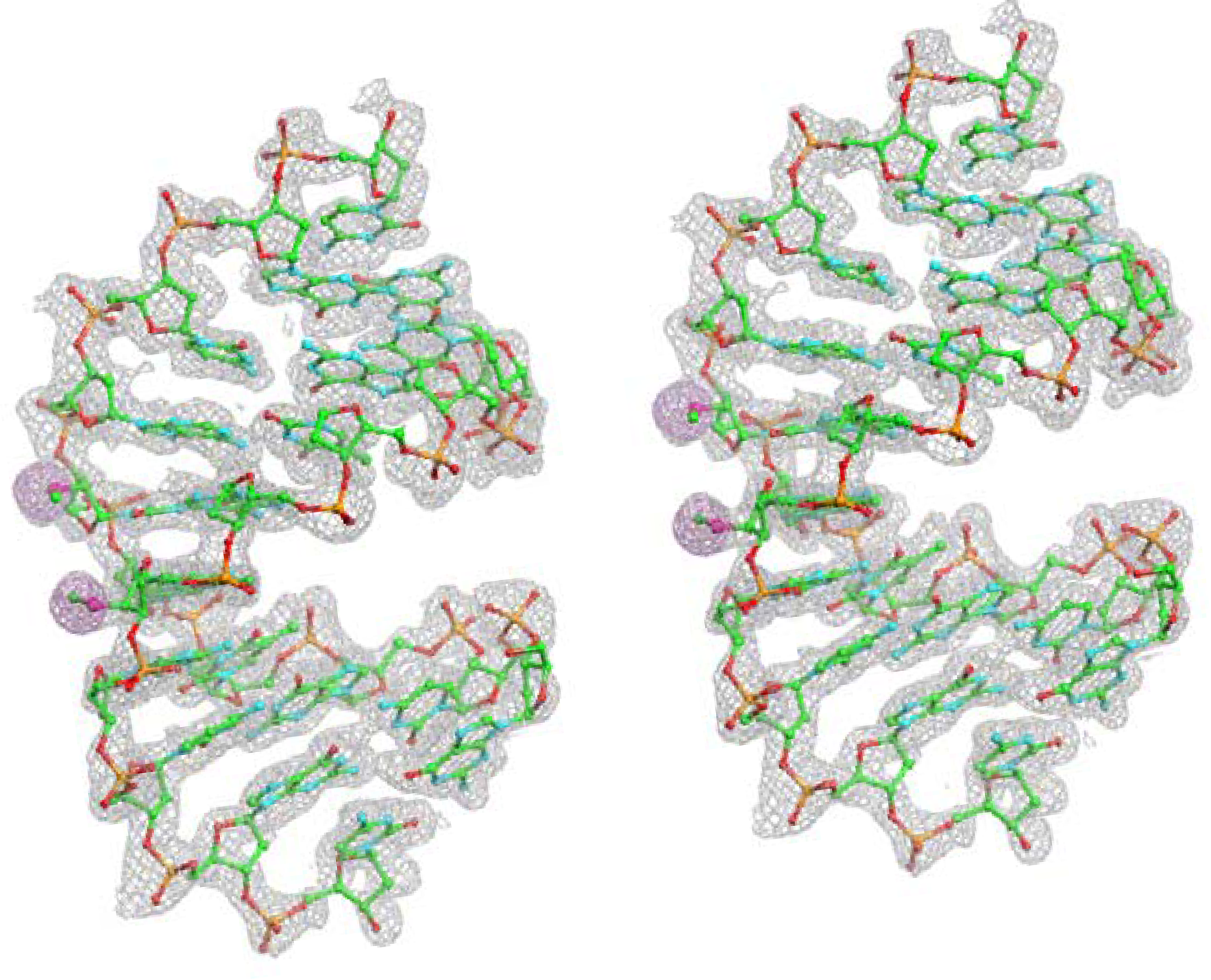{kind=link}
{kind=link}
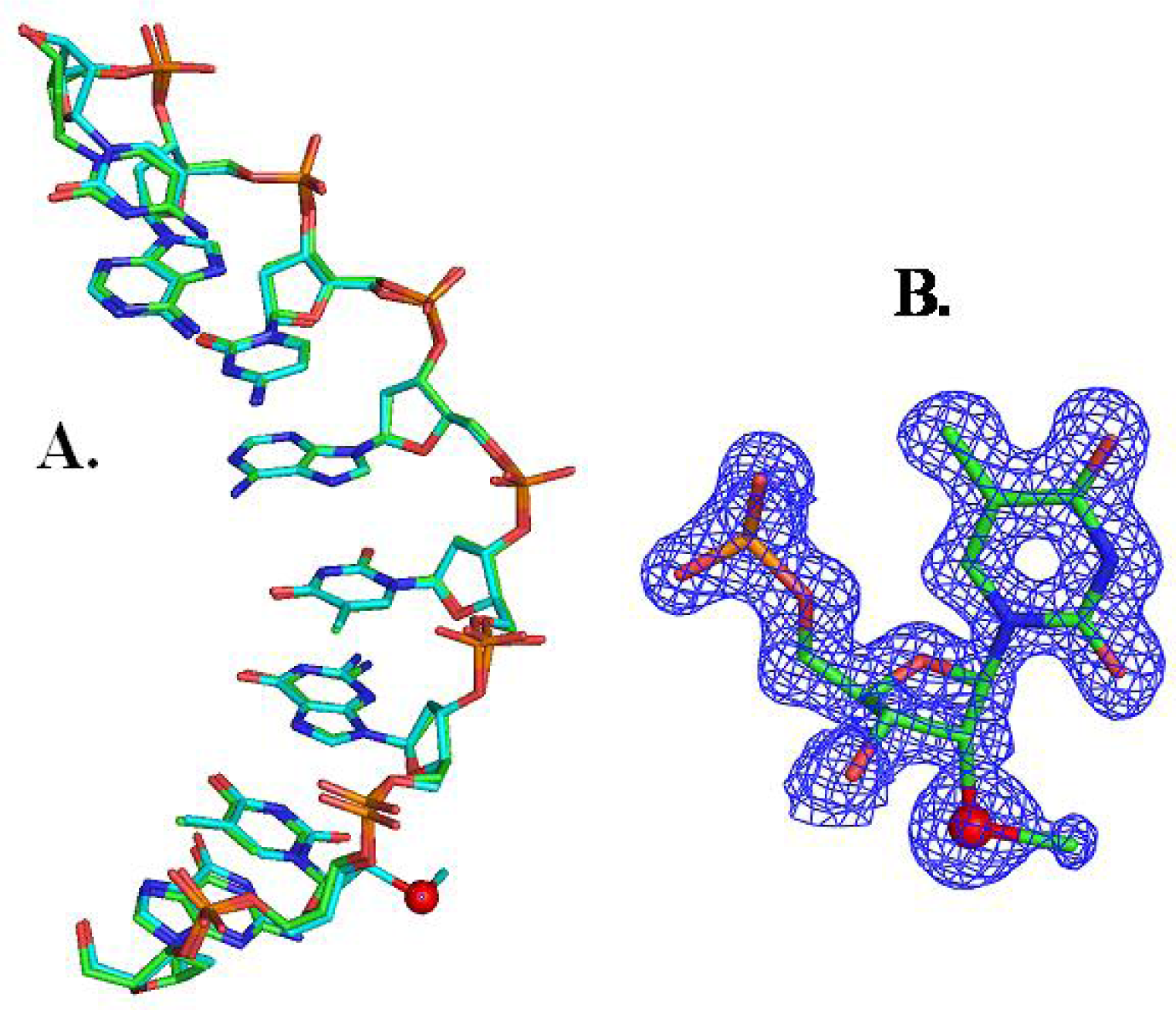{kind=link}
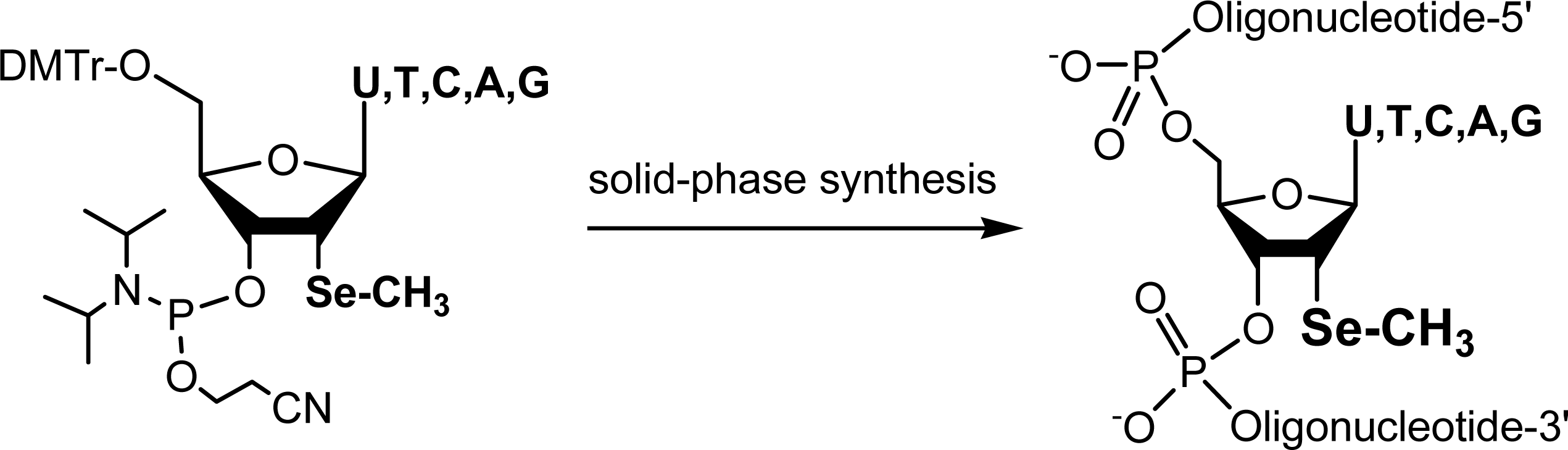{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
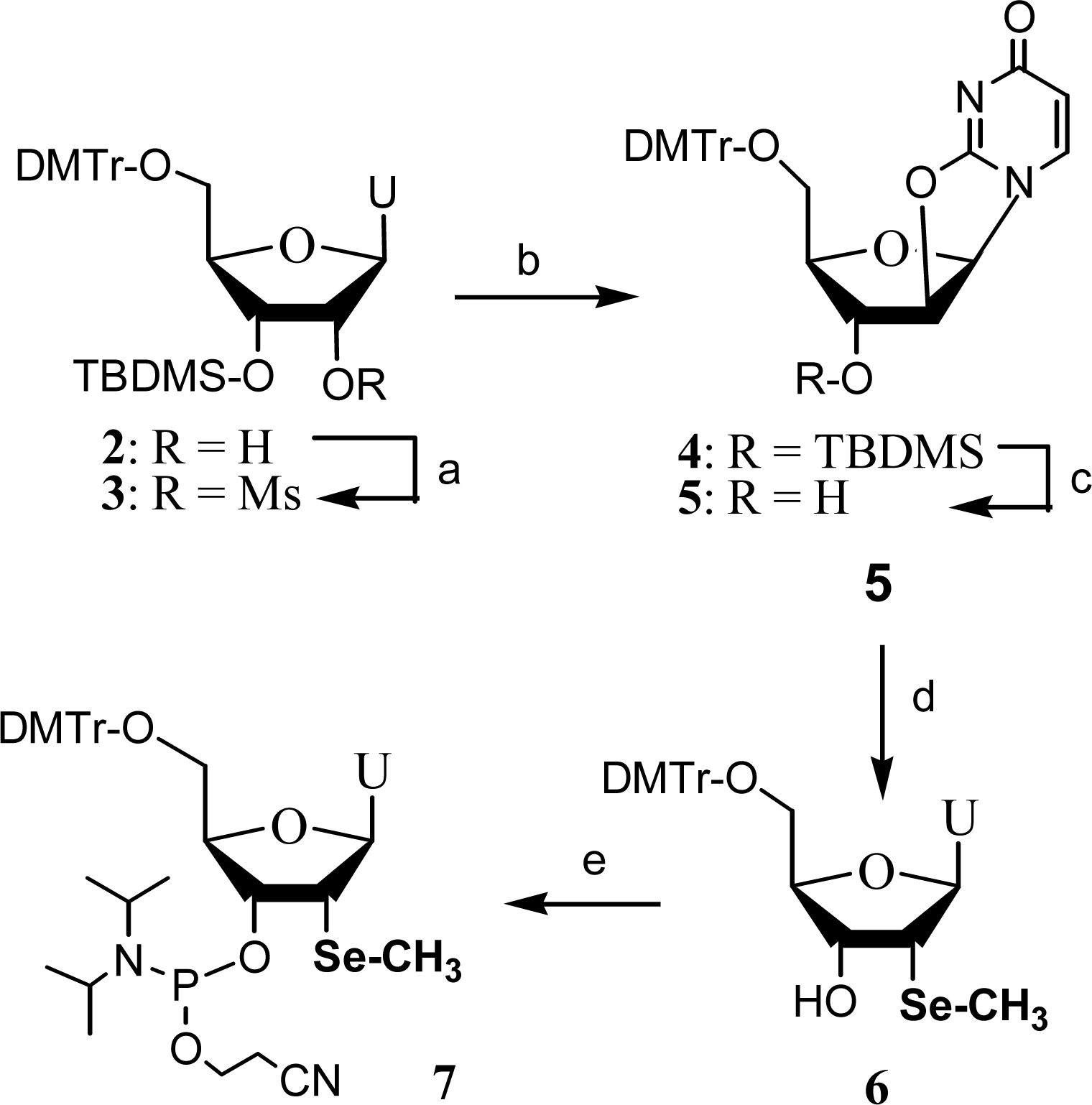
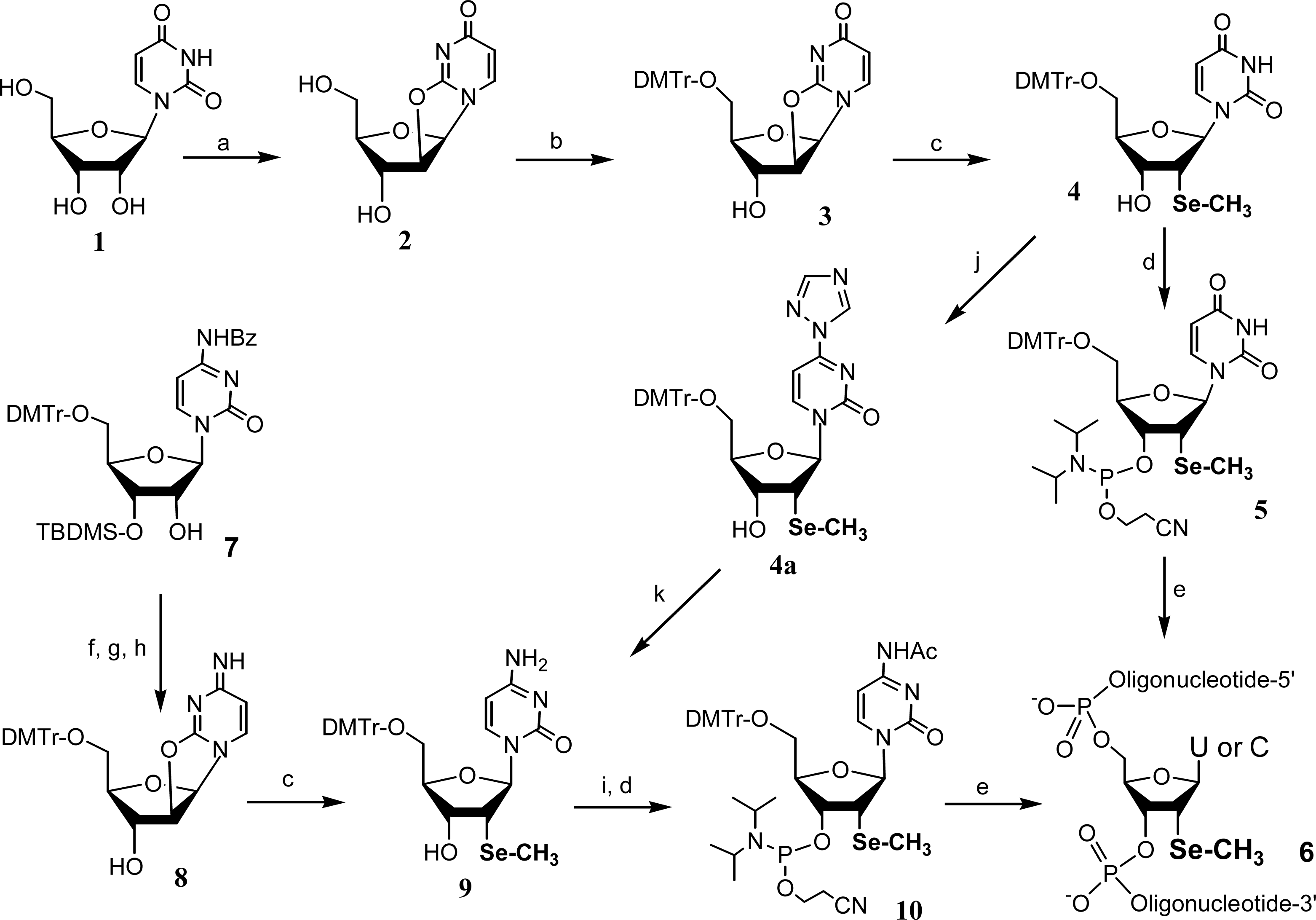
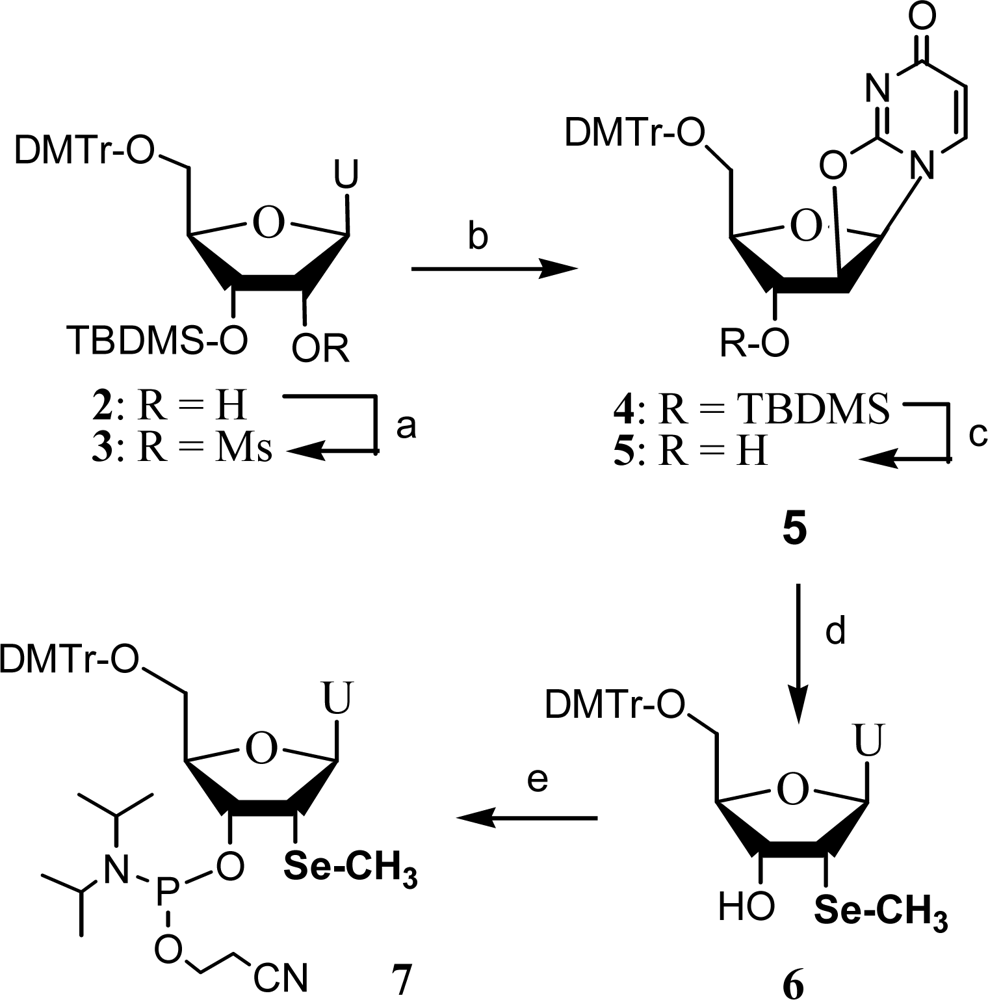
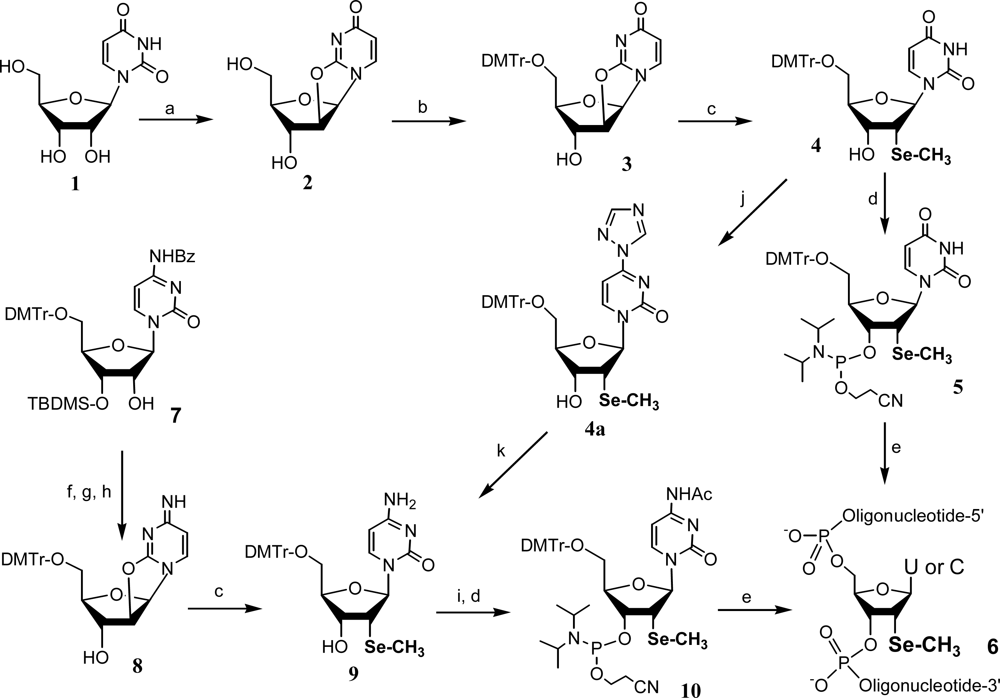
2. Synthesis and application of 2’-Se-pyrimidines
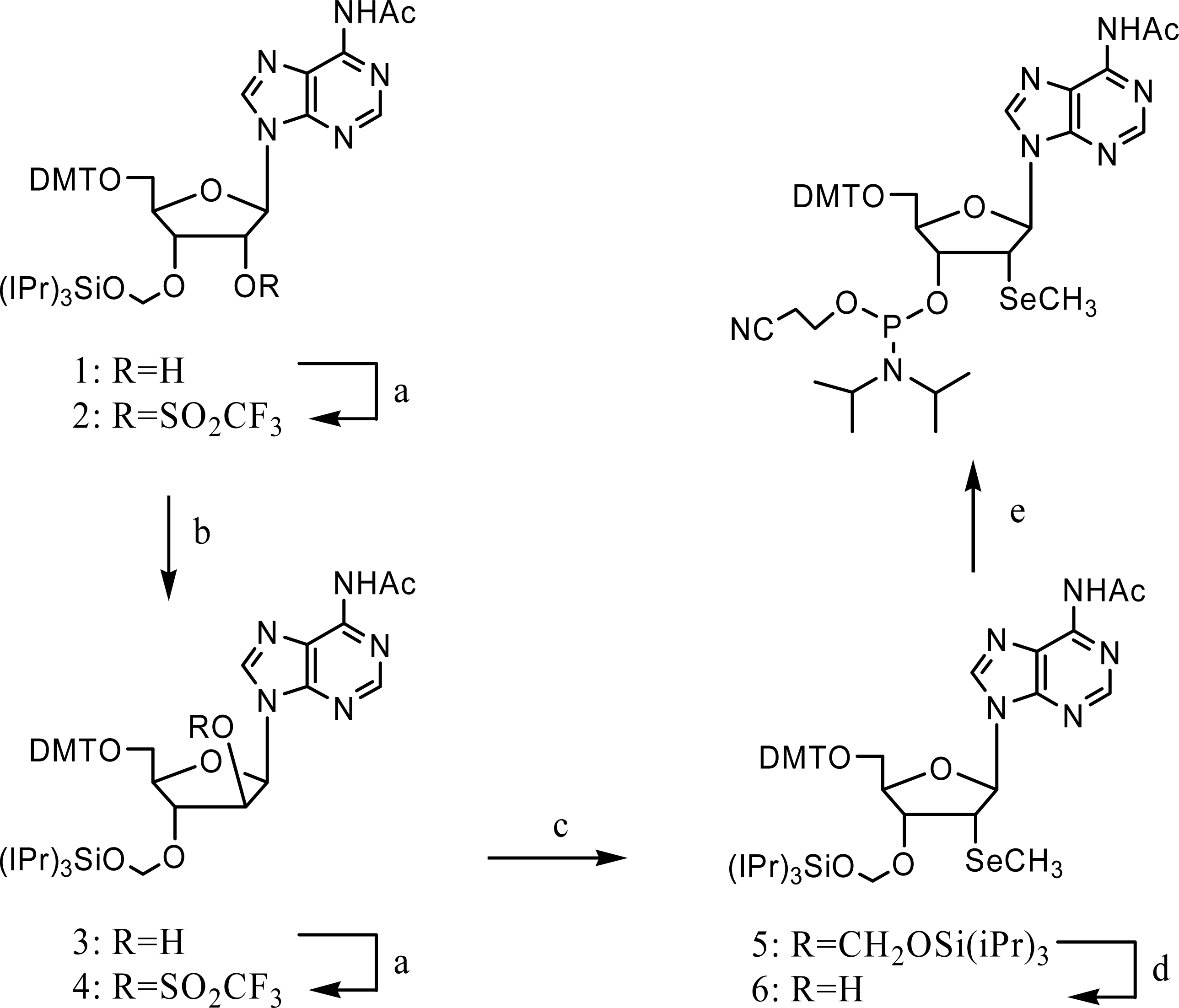
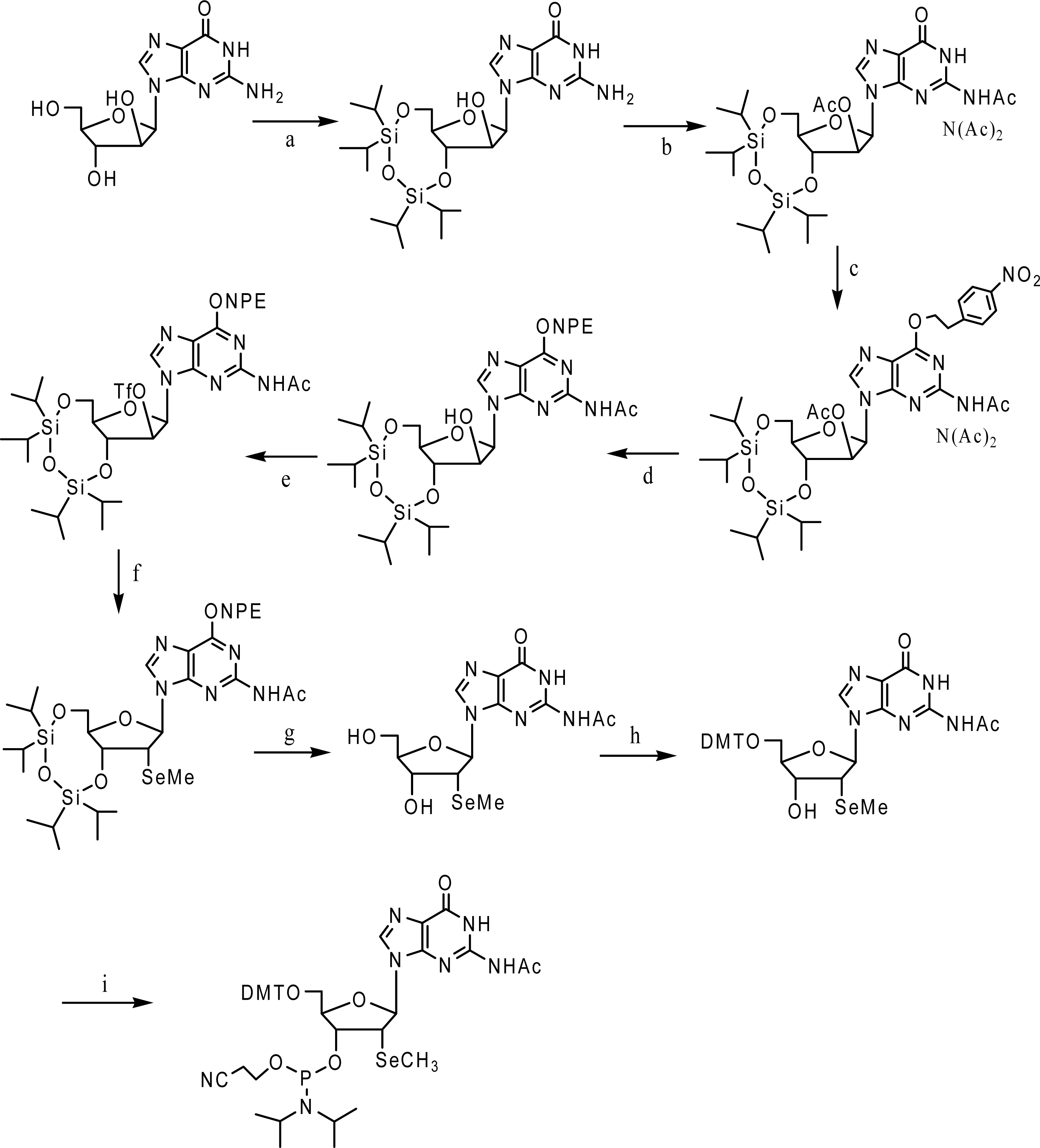
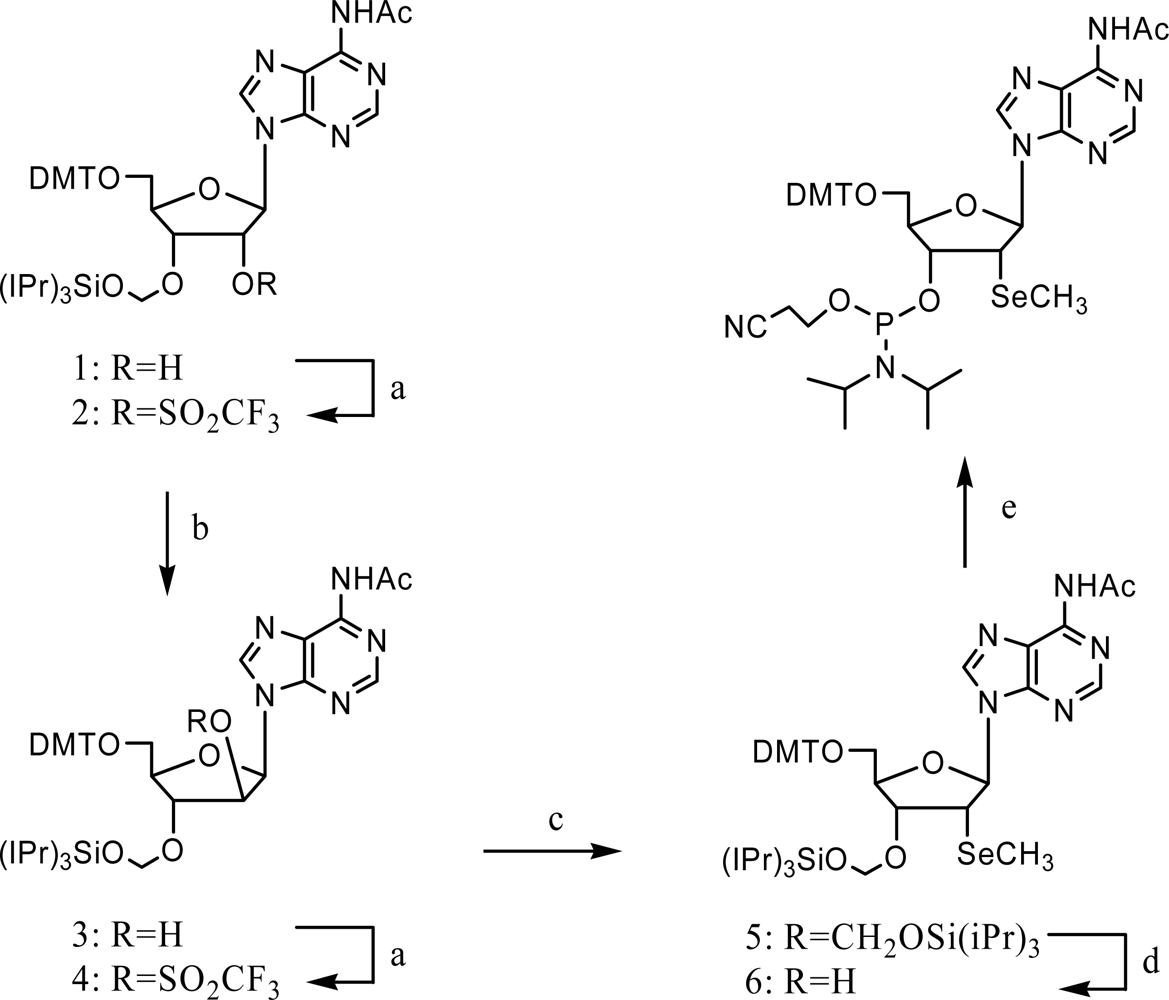
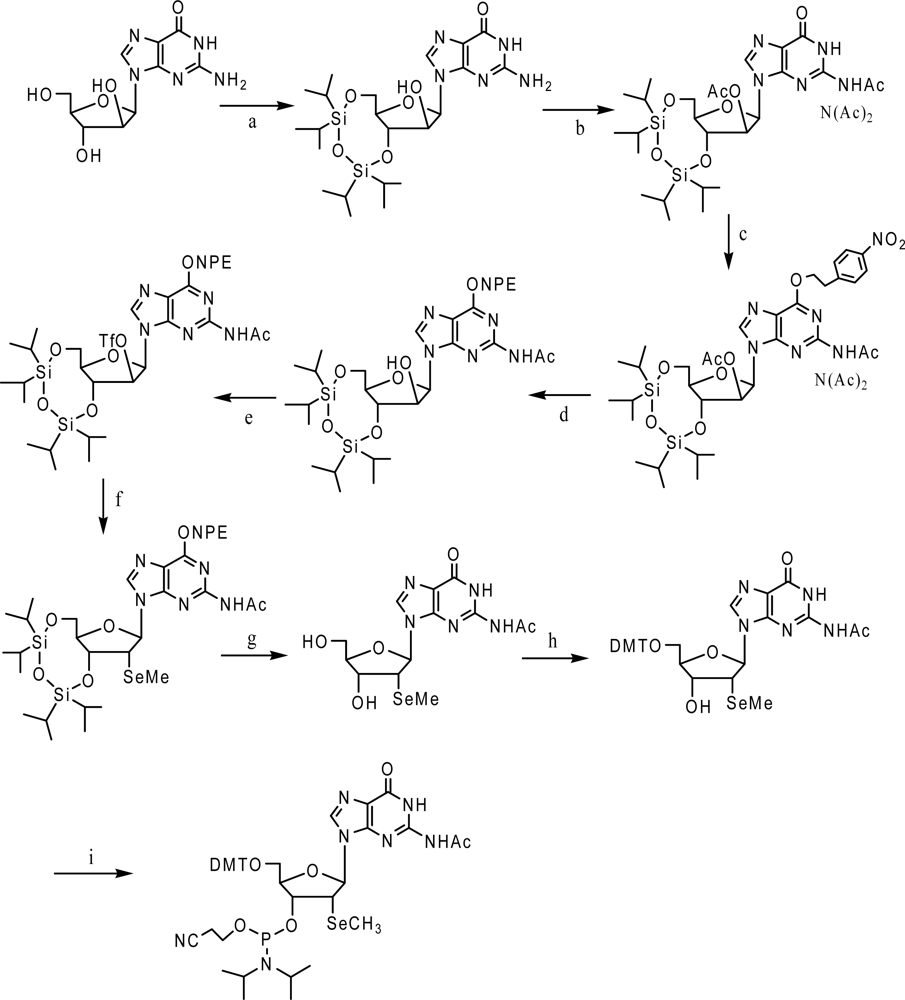
3. Synthesis and application of 2’-Se-purines
4. Facilitation of DNA Crystallization by Selenium Derivatization
5. Prospects of Selenium Derivatization of Nucleic Acids
Acknowledgments
References and Notes
- Noller, HF. RNA structure: reading the ribosome. Science 2005, 309, 1508–1514. [Google Scholar]
- Hendrix, DK; Brenner, SE; Holbrook, SR. RNA structural motifs: building blocks of a modular biomolecule. Q. Rev. Biophys. 2005, 38, 221–243. [Google Scholar]
- Doudna, JA; Lorsch, JR. Ribozyme catalysis: not different, just worse. Nat. Struct. Mol. Biol. 2005, 12, 395–402. [Google Scholar]
- Sali, A; Glaeser, R; Earnest, T; Baumeister, W. From words to literature in structural proteomics. Nature 2003, 422, 216–225. [Google Scholar]
- Hendrickson, WA. Synchrotron crystallography. Trends Biochem. Sci. 2000, 25, 637–643. [Google Scholar]
- Egli, M. Nucleic acid crystallography: current progress. Curr. Opin. Chem. Biol. 2004, 8, 580–591. [Google Scholar]
- Holbrook, SR. RNA structure: the long and the short of it. Curr. Opin. Struct. Biol. 2005, 15, 302–308. [Google Scholar]
- Egli, M; Pallan, PS. Insights from crystallographic studies into the structural and pairing properties of nucleic acid analogs and chemically modified DNA and RNA oligonucleotides. Annu. Rev. Biophys. Biomol. Struct. 2007, 36, 281–305. [Google Scholar]
- Yang, W; Hendrickson, WA; Crouch, RJ; Satow, Y. Structure of ribonuclease H phased at 2 A resolution by MAD analysis of the selenomethionyl protein. Science 1990, 249, 1398–1405. [Google Scholar]
- Ferre-D'Amare, AR; Zhou, K; Doudna, JA. Crystal structure of a hepatitis delta virus ribozyme. Nature 1998, 395, 567–574. [Google Scholar]
- Garman, E; Murray, JW. Heavy-atom derivatization. Acta. Crystallogr. D: Biol. Crystallogr. 2003, 59, 1903–1913. [Google Scholar]
- Xiong, Y; Sundaralingam, M. Crystal structure of a DNA.RNA hybrid duplex with a polypurine RNA r(gaagaagag) and a complementary polypyrimidine DNA d(CTCTTCTTC). Nucleic Acids Res. 2000, 28, 2171–2176. [Google Scholar]
- Boggon, TJ; Shapiro, L. Screening for phasing atoms in protein crystallography. Structure 2000, 8, R143–R149. [Google Scholar]
- Shah, K; Wu, H; Rana, TM. Synthesis of uridine phosphoramidite analogs: reagents for site-specific incorporation of photoreactive sites into RNA sequences. Bioconjug. Chem. 1994, 5, 508–512. [Google Scholar]
- Jiang, J; Sheng, J; Carrasco, N; Huang, Z. Selenium derivatization of nucleic acids for crystallography. Nucleic Acids Res. 2007, 35, 477–485. [Google Scholar]
- Ennifar, E; Carpentier, P; Ferrer, JL; Walter, P; Dumas, P. X-ray-induced debromination of nucleic acids at the Br K absorption edge and implications for MAD phasing. Acta. Crystallogr. D: Biol. Crystallogr. 2002, 58, 1262–1268. [Google Scholar]
- Hendrickson, WA; Horton, JR; LeMaster, DM. Selenomethionyl proteins produced for analysis by multiwavelength anomalous diffraction (MAD): a vehicle for direct determination of three-dimensional structure. EMBO J. 1990, 9, 1665–1672. [Google Scholar]
- Hendrickson, WA. Determination of macromolecular structures from anomalous diffraction of synchrotron radiation. Science 1991, 254, 51–58. [Google Scholar]
- Deacon, AM; Ealick, SE. Selenium-based MAD phasing: setting the sites on larger structures. Structure 1999, 7, R161–R166. [Google Scholar]
- Jiang, J; Sweet, RM. Protein Data Bank depositions from synchrotron sources. J. Synchrotron Radiat. 2004, 11, 319–327. [Google Scholar]
- Rupert, PB; Ferre-D'Amare, AR. Crystal structure of a hairpin ribozyme-inhibitor complex with implications for catalysis. Nature 2001, 410, 780–786. [Google Scholar]
- Carrasco, N; Ginsburg, D; Du, Q; Huang, Z. Synthesis of selenium-derivatized nucleosides and oligonucleotides for X-ray crystallography. Nucleos. Nucleot. Nucleic Acids 2001, 20, 1723–1734. [Google Scholar]
- Du, Q; Carrasco, N; Teplova, M; Wilds, CJ; Egli, M; Huang, Z. Internal derivatization of oligonucleotides with selenium for X-ray crystallography using MAD. J. Am. Chem. Soc. 2002, 124, 24–25. [Google Scholar]
- Buzin, Y; Carrasco, N; Huang, Z. Synthesis of selenium-derivatized cytidine and oligonucleotides for X-ray crystallography using MAD. Org. Lett. 2004, 6, 1099–1102. [Google Scholar]
- Carrasco, N; Buzin, Y; Tyson, E; Halpert, E; Huang, Z. Selenium derivatization and crystallization of DNA and RNA oligonucleotides for X-ray crystallography using multiple anomalous dispersion. Nucleic Acids Res. 2004, 32, 1638–1646. [Google Scholar]
- Carrasco, N; Huang, Z. Enzymatic synthesis of phosphoroselenoate DNA using thymidine 5'-(alpha-P-seleno)triphosphate and DNA polymerase for X-ray crystallography via MAD. J. Am. Chem. Soc. 2004, 126, 448–449. [Google Scholar]
- Salon, J; Chen, G; Portilla, Y; Germann, MW; Huang, Z. Synthesis of a 2'-Se-uridine phosphoramidite and its incorporation into oligonucleotides for structural study. Org. Lett. 2005, 7, 5645–5648. [Google Scholar]
- Brandt, G; Carrasco, N; Huang, Z. Efficient substrate cleavage catalyzed by hammerhead ribozymes derivatized with selenium for X-ray crystallography. Biochemistry 2006, 45, 8972–8977. [Google Scholar]
- Carrasco, N; Caton-Williams, J; Brandt, G; Wang, S; Huang, Z. Efficient enzymatic synthesis of phosphoroselenoate RNA by using adenosine 5'-(alpha-P-seleno) triphosphate. Angew. Chem. Int. Ed. Engl. 2006, 45, 94–97. [Google Scholar]
- Sheng, J; Jiang, J; Salon, J; Huang, Z. Synthesis of a 2'-Se-thymidine Phosphoramidite and Its Incorporation into Oligonucleotides for Crystal Structure Study. Org. Lett. 2007, 9, 749–752. [Google Scholar]
- Salon, J; Sheng, J; Jiang, J; Chen, G; Caton-Williams, J; Huang, Z. Oxygen replacement with selenium at the thymidine 4-position for the Se base pairing and crystal structure studies. J. Am. Chem. Soc. 2007, 129, 4862–4863. [Google Scholar]
- Teplova, M; Wilds, CJ; Wawrzak, Z; Tereshko, V; Du, Q; Carrasco, N; Huang, Z; Egli, M. Covalent incorporation of selenium into oligonucleotides for X-ray crystal structure determination via MAD: proof of principle. Multiwavelength anomalous dispersion. Biochimie 2002, 84, 849–858. [Google Scholar]
- Wilds, CJ; Pattanayek, R; Pan, C; Wawrzak, Z; Egli, M. Selenium-assisted nucleic acid crystallography: use of phosphoroselenoates for MAD phasing of a DNA structure. J. Am. Chem. Soc. 2002, 124, 14910–14916. [Google Scholar]
- Serganov, A; Yuan, YR; Pikovskaya, O; Polonskaia, A; Malinina, L; Phan, AT; Hobartner, C; Micura, R; Breaker, RR; Patel, DJ. Structural basis for discriminative regulation of gene expression by adenine- and guanine-sensing mRNAs. Chem. Biol. 2004, 11, 1729–1741. [Google Scholar]
- Serganov, A; Keiper, S; Malinina, L; Tereshko, V; Skripkin, E; Hobartner, C; Polonskaia, A; Phan, AT; Wombacher, R; Micura, R; Dauter, Z; Jaschke, A; Patel, DJ. Structural basis for Diels-Alder ribozyme-catalyzed carbon-carbon bond formation. Nat. Struct. Mol. Biol. 2005, 12, 218–224. [Google Scholar]
- Moroder, H; Kreutz, C; Lang, K; Serganov, A; Micura, R. Synthesis, oxidation behavior, crystallization and structure of 2'-methylseleno guanosine containing RNAs. J. Am. Chem. Soc. 2006, 128, 9909–9918. [Google Scholar]
- Egli, M; Pallan, PS; Pattanayek, R; Wilds, CJ; Lubini, P; Minasov, G; Dobler, M; Leumann, CJ; Eschenmoser, A. Crystal Structure of Homo-DNA and Nature's Choice of Pentose over Hexose in the Genetic System. J. Am. Chem. Soc. 2006, 128, 10847–10856. [Google Scholar]
- Gerlits, O; Jiang, J; Salon, J; Huang, Z. manuscript in preparation; 2007. [Google Scholar]
- Hobartner, C; Micura, R. Chemical synthesis of selenium-modified oligoribonucleotides and their enzymatic ligation leading to an U6 SnRNA stem-loop segment. J. Am. Chem. Soc. 2004, 126, 1141–1149. [Google Scholar]









Share and Cite
Sheng, J.; Huang, Z. Selenium Derivatization of Nucleic Acids for Phase and Structure Determination in Nucleic Acid X-ray Crystallography. Int. J. Mol. Sci. 2008, 9, 258-271. https://doi.org/10.3390/ijms9030258
Sheng J, Huang Z. Selenium Derivatization of Nucleic Acids for Phase and Structure Determination in Nucleic Acid X-ray Crystallography. International Journal of Molecular Sciences. 2008; 9(3):258-271. https://doi.org/10.3390/ijms9030258
Chicago/Turabian StyleSheng, Jia, and Zhen Huang. 2008. "Selenium Derivatization of Nucleic Acids for Phase and Structure Determination in Nucleic Acid X-ray Crystallography" International Journal of Molecular Sciences 9, no. 3: 258-271. https://doi.org/10.3390/ijms9030258
APA StyleSheng, J., & Huang, Z. (2008). Selenium Derivatization of Nucleic Acids for Phase and Structure Determination in Nucleic Acid X-ray Crystallography. International Journal of Molecular Sciences, 9(3), 258-271. https://doi.org/10.3390/ijms9030258