Effect of Climatic Conditions and Land Cover on Genetic Structure and Diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
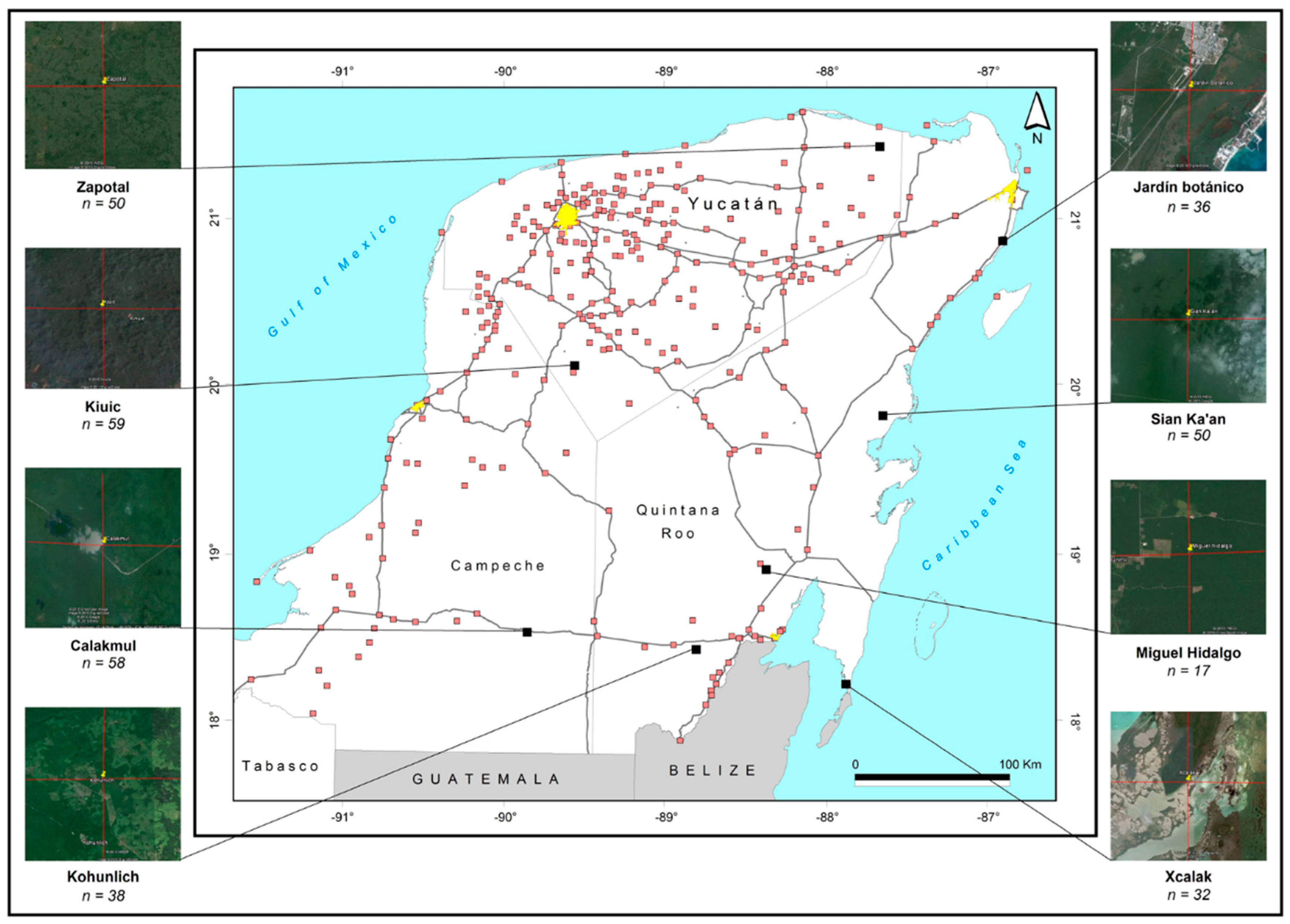
2.1. Study Sites and Butterfly Samples
2.2. DNA Extraction
2.3. ISSR-PCR Amplification
2.4. Molecular Analysis
2.5. Genetic Diversity
2.6. Relation among Genetic Diversity, Environmental Conditions, and Land Cover
2.7. Population Differentiation and Genetic Structure
2.8. Sex-Bias Dispersal Analysis
3. Results
3.1. Genetic Diversity
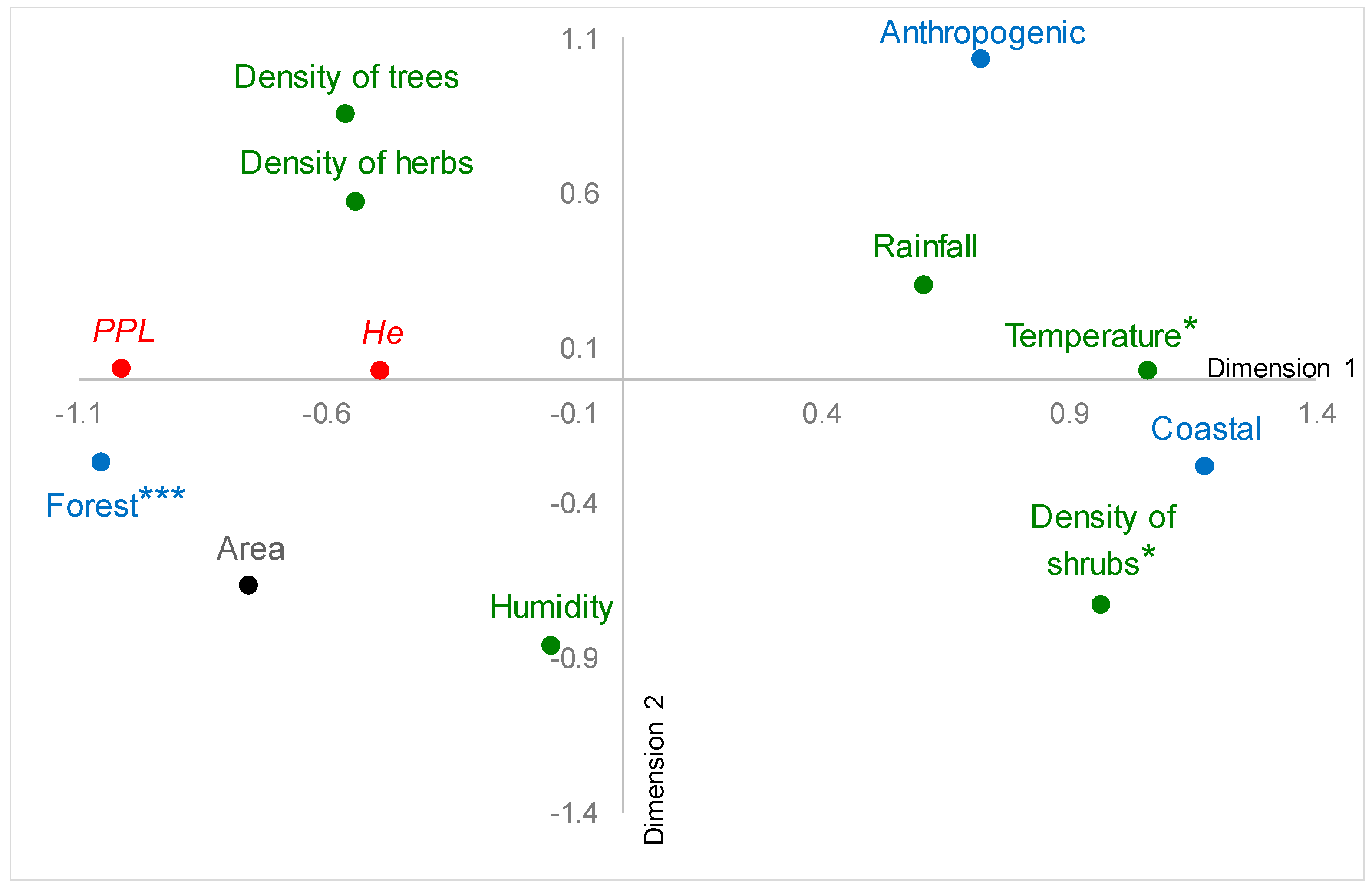
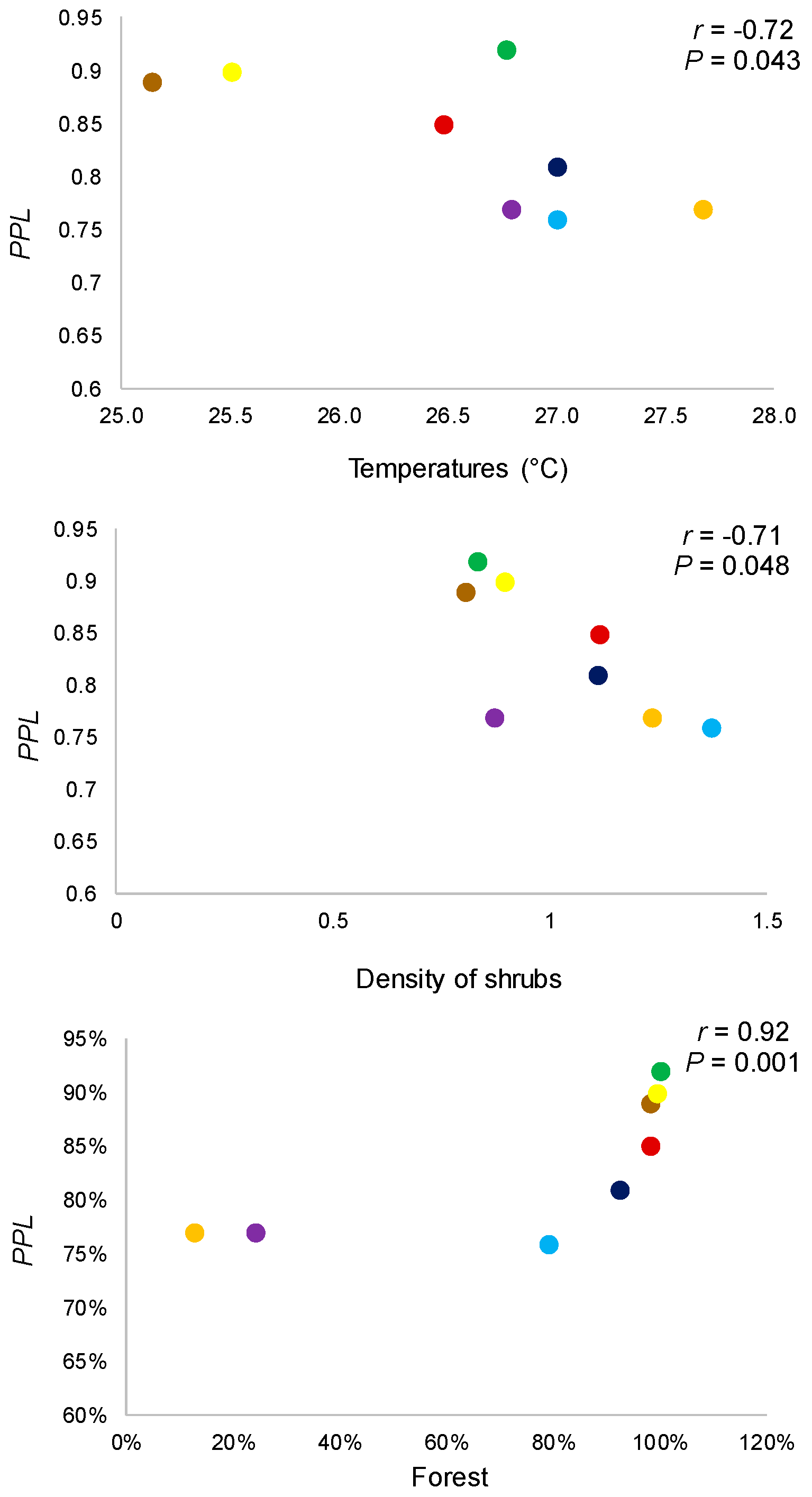
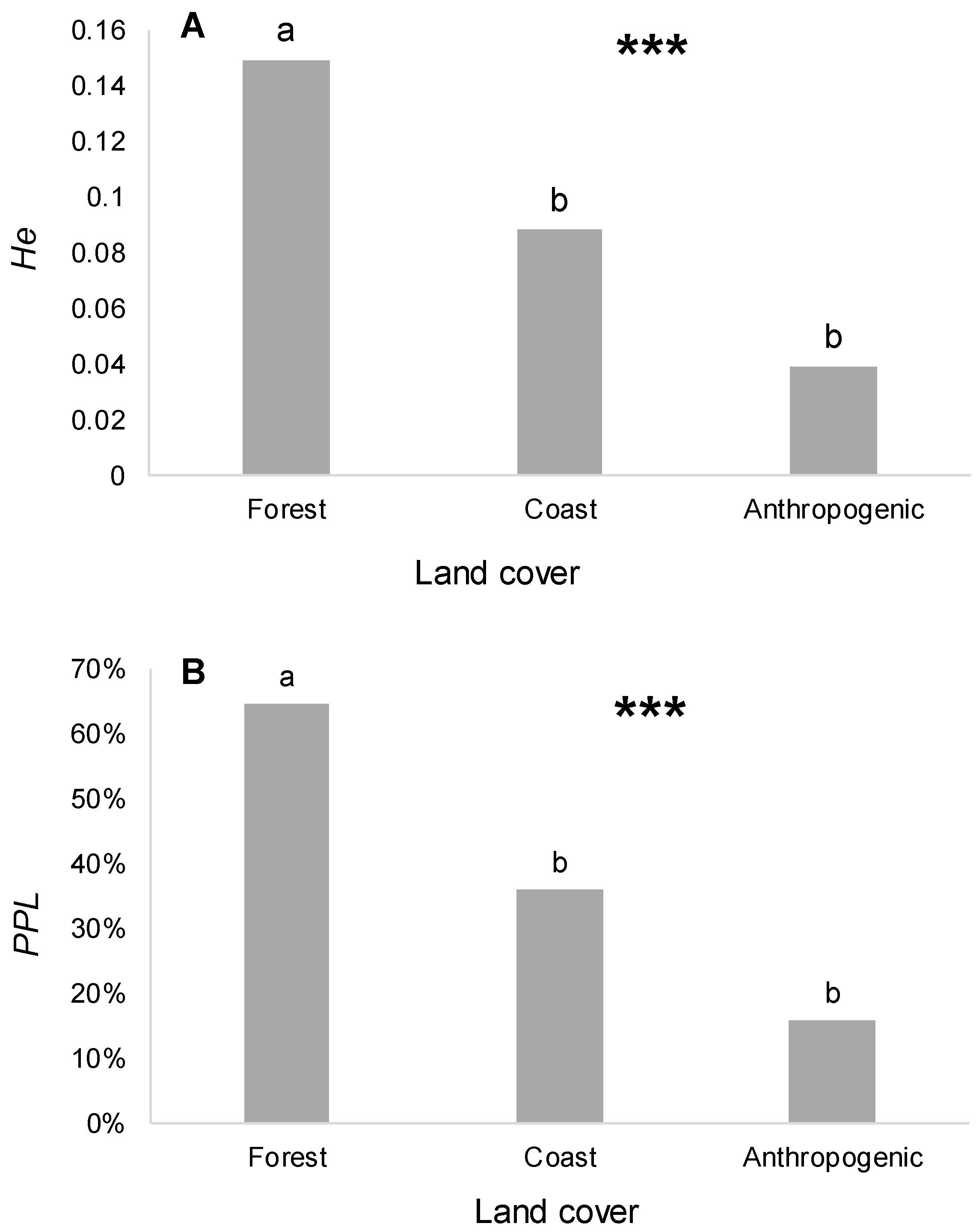
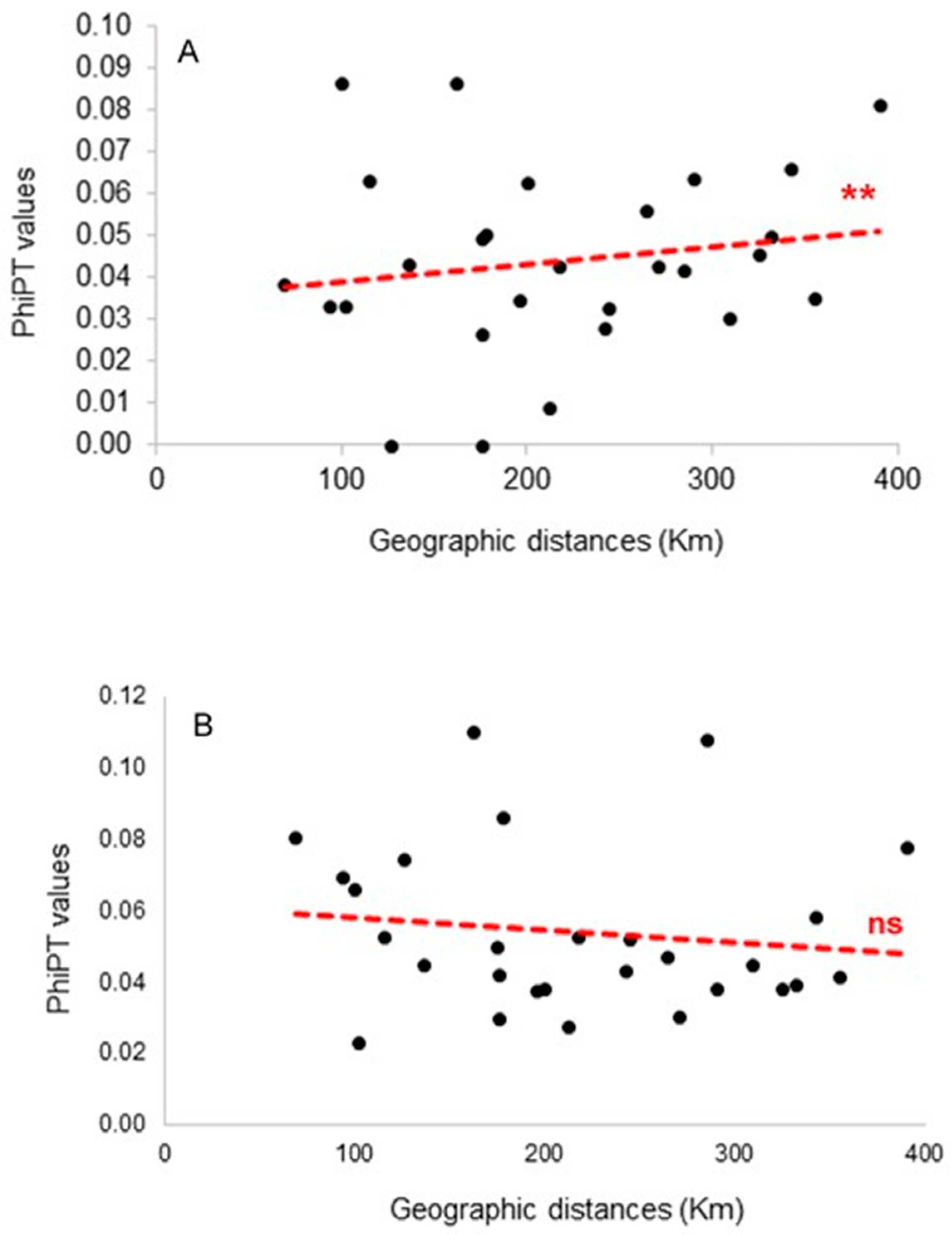
3.2. Relation among Genetic Diversity, Environmental Conditions, and Land Cover
3.3. Population Differentiation and Genetic Structure
3.4. Sex-Bias Dispersal Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franklin, A.B.; Noon, B.R.; George, T.L. What is habitat Fragmentation? Stud. Avian Biol. 2002, 25, 20–29. [Google Scholar]
- Frankham, R.; Ballou, J.D.; Briscoe, D. Introduction to Conservation Genetics, 2nd ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Franklin, J.F.; Lindenmayer, D.B. Importance of matrix habitats in maintaining biological diversity. Proc. Natl. Acad. Sci. USA 2009, 106, 349–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischendorf, L.; Fahrig, L. On the usage and measurement of landscape connectivity. Oikos 2000, 90, 7–19. [Google Scholar] [CrossRef]
- McKenzie, D.H.; Hyatt, D.E.; McDonald, V.J. Ecological Indicators; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Daily, G.C.; Ehrlich, P.R. Preservation of biodiversity in small rainforest patches: Rapid evaluations using butterfly trapping. Biodivers. Conserv. 1995, 4, 35–55. [Google Scholar] [CrossRef]
- Brown, K.S., Jr.; Freitas, A.V.L. Atlantic forest butterflies: Indicators for landscape conservation. Biotropica 2000, 32, 934–956. [Google Scholar] [CrossRef]
- Syaripuddin, K.; Sing, K.W.; Wilson, J.J. Comparison of butterflies, bats and beetles as bioindicators based on four key criteria and DNA barcodes. Trop. Conserv. Sci. 2015, 8, 138–149. [Google Scholar] [CrossRef]
- Whitworth, A.; Huarcaya, R.P.; Mercado, H.G.; Braunholtz, L.D.; MacLeod, R. Food for thought. Rainforest carrion-feeding butterflies are more sensitive indicators of disturbance history than fruit feeders. Biol. Conserv. 2018, 217, 383–390. [Google Scholar] [CrossRef]
- Luque, C.; Legal, L.; Winterton, P.; Mariano, N.A.; Gers, C. Illustration of the Structure of Arthropod Assemblages (Collembola and Lepidoptera) in Different Forest Types: An Example in the French Pyrenees. Diversity 2011, 3, 693–711. [Google Scholar] [CrossRef] [Green Version]
- Van Swaay, C.A.M.; Van Strien, A.J.; Aghababyan, K.; Åström, S.; Botham, M. The European Butterfly Indicator for Grassland Species 1990–2013; Report VS2015.009; De Vlinderstichting: Wageningen, The Netherland, 2015. [Google Scholar]
- Pozo, C. Los Rhopalocera de la Región de Calakmul, Campeche: Métodos de Estudio, Fenología y su Uso Como Indicadores de Disturbio. Ph.D. Thesis, UNAM, México, Mexico, 2006. [Google Scholar]
- Legrand, D.; Trochet, A.; Moulherat, S.; Calvez, O.; Stevens, V.M.; Ducatez, S.; Clobert, J.; Baguette, M. Ranking the ecological causes of dispersal in a butterfly. Ecography 2015, 38, 822–831. [Google Scholar] [CrossRef]
- Ducatez, S.; Baguette, M.; Trochet, A.; Chaput-Bardy, A.; Legrand, D. Flight endurance and heating rate vary with both latitude and habitat connectivity in a butterfly species. Oikos 2013, 122, 601–611. [Google Scholar] [CrossRef]
- Roux, O.; Gevrey, M.; Arvanitakis, L.; Gers, C.; Bordat, D.; Legal, L. ISSR–PCR: Tool for population identification and genetic structure of Plutella xylostella. Mol. Phylogenet. Evol. 2007, 43, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Machkour-M’Rabet, S.; Leberger, R.; León-Cortés, J.L.; Gers, C.; Legal, L. Population structure and genetic diversity of the only extant Baroninae swallowtail butterfly, Baronia brevicornis, revealed by ISSR markers. J. Insect Conserv. 2014, 18, 385–396. [Google Scholar] [CrossRef]
- Leidner, A.K.; Haddad, N.M. Natural, not urban, barriers define population structure for a coastal endemic butterfly. Conserv. Genet. 2010, 11, 2311–2320. [Google Scholar] [CrossRef]
- Berwaerts, K.; Van Dyck, H.; Van Dongen, S.; Matthysen, E. Morphological and genetic variation in the speckled wood butterfly (Pararge aegeria L.) among differently fragmented landscapes. Neth. J. Zool. 1997, 48, 241–253. [Google Scholar] [CrossRef]
- Williams, B.L.; Brawn, J.D.; Paige, K. Landscape scale genetic effects of habitat fragmentation on a high gene flow species: Speyeria idalia (Nymphalidae). Mol. Ecol. 2003, 12, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.F.; Gomez, L.M.; Rubio, A.; Bock, B.; Uribe, S. Estructura genética de mariposas en un paisaje fragmentado: Una aproximación al manejo ambiental (Antioquia, Colombia). Rev. Lasallista Investig. 2005, 2, 27–32. [Google Scholar]
- Maya-Martínez, M.; Pozo, C.; Schmitter-Soto, J.J. Distribution patterns of Charaxinae (Lepidoptera: Nymphalidae) in Yucatan Peninsula, Mexico. Acta Zool. Mex. 2009, 25, 283–301. [Google Scholar] [CrossRef]
- Gibbs, M.; Wiklund, C.; Van Dyck, H. Temperature, rainfall and butterfly morphology: Does life history theory match the observed pattern? Ecography 2011, 34, 336–344. [Google Scholar] [CrossRef]
- Vandewoestijne, S.; Schtickzelle, N.; Baguette, M. Positive correlation between genetic diversity and fitness in a large, well-connected metapopulation. BMC Biol. 2008, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- González-Iturbe, A.; Granados Castellanos, J.; Tun Dzul, F.; Olmsted, I. Vegetación Atlas de Procesos Territoriales de Yucatán; Universidad Autónoma de Yucatán: México, México, 1999. [Google Scholar]
- Rada, J.M.D.; García, R.D.; García-Contreras, G.; Morín, J.A.; Lugo, E.A. Conservation and Use. In Biodiversity and Conservation of the Yucatán Peninsula; Islebe, G., Calmé, S., León-Cortés, J.L., Schmook, B., Eds.; Springer International Publishing: Basel, Switzerland, 2015; pp. 169–193. [Google Scholar]
- Vázquez-Domínguez, E.; Arita, H. The Yucatán Peninsula: Biogeographical history 65 million years in the making. Ecography 2010, 33, 212–219. [Google Scholar] [CrossRef]
- Carnevali, G.; Ramírez, I.M.; González-Iturbe, J.A. Flora y vegetación de la península de Yucatán. In Naturaleza y Sociedad en el Área Maya, Pasado, Presente y Futuro; Colunga-García Marín, P., Larqué-Saavedra, A., Eds.; Academia Mexicana de Ciencias, CICY: Mérida, Mexico, 2003; pp. 53–67. [Google Scholar]
- Abrams, E.M.; Rue, D.J. The causes and consequences of deforestation among the prehistoric Maya. Hum. Ecol. 1988, 16, 377–395. [Google Scholar] [CrossRef]
- Geoghegan, J.; Cortina, S.V.; Klepeis, P.; Macario, P.M.; Ogneva-Himmelberger, Y.; Chowdhury, R.R. Modeling tropical deforestation in the southern Yucatán peninsular region: Comparing survey and satellite data. Agric. Ecosyst. Environ. 2001, 85, 25–46. [Google Scholar] [CrossRef]
- Pozo, C.; Salas-Suárez, N.; Maya, A. Mariposas Riqueza Biológica de Quintana Roo Un Análisis Para su Conservación; Tomo 2; El Colegio de la Frontera Sur (ECOSUR), Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (Conabio), Gobierno del Estado de Quintana Roo y Programa de Pequeñas Donaciones (PPD): México, México, 2011.
- Pozo, C.; Cabrera, E.; Rangel, J.L.; Viveros, P. Fauna Estudios Ecológicos Preliminares de la Zona sur de Quintana Roo México; Centro de Investigaciones de Quintana Roo: Mexico, Mexico, 1991. [Google Scholar]
- Pozo, C.; Luis-Martínez, A.; Uc, S.; Salas, N.; Maya, A. Butterflies (Papilionoidea and Hesperioidea) of Calakmul, Campeche, México. Southwest. Nat. 2003, 48, 505–525. [Google Scholar] [CrossRef]
- Pozo, C.; Salas-Suárez, N.; Prado, B.R.; May, E. Riqueza de mariposas diurnas (Lepidoptera: Rhopalocera) en el Santuario del Manatí y una propuesta para su uso en el monitoreo de ambientes terrestres del área. In El Sistema Ecológico de la Bahía de Chetumal/Corozal: Costa Occidental del Mar Caribe; Espinoza-Ávalos, J., Islebe, G.A., Hernández-Arana, H.A., Eds.; El Colegio de la Frontera Sur: Mexico, Mexico, 2009; pp. 139–147. [Google Scholar]
- Vargas-Fernández, I.; Llorente-Bousquets, J.; Luis-Martínez, A.; Pozo, C. Nymphalidae de México II (Libytheinae, Ithomiinae, Morphinae y Charaxinae): Distribución Geográfica e Ilustración; UNAM: Mexico, Mexico, 2008. [Google Scholar]
- Prado, B.R.; Pozo, C.; Valdez-Moreno, M.; Hebert, P.D.N. Beyond the Colours: Discovering Hidden Diversity in the Nymphalidae of the Yucatan Peninsula in Mexico through DNA Barcoding. PLoS ONE 2011, 6, e27776. [Google Scholar] [CrossRef] [PubMed]
- Cavanzón-Medrano, L.; Pozo, C.; Hénaut, Y.; Legal, L.; Salas-Suárez, N.; Machkour-M’Rabet, S. Complex Population Patterns of Eunica tatila Herrich-Schäffer (Lepidoptera: Nymphalidae), with Special Emphasis on Sexual Dimorphism. Neotrop. Entomol. 2016, 45, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Rosin, Z.M.; Myczko, L.; Piotr Skorka, P.; Lenda, M.; Moron, D.; Sparks, T.H.; Tryjanowski, P. Butterfly responses to environmental factors in fragmented calcareous grasslands. J. Insect Conserv. 2012, 16, 321–329. [Google Scholar] [CrossRef]
- Jenkins, D.W. Neotropical Nymphalidae VIII. Revision of Eunica. Bull. Allyn. Mus. 1990, 131, 1–175. [Google Scholar]
- Andrade-C, M.G.; Henao Bañol, E.R.; Triviño, P. Técnicas y procesamiento para la recolección, preservación y montaje de mariposas en estudios de biodiversidad y conservación (Lepidoptera: Hesperoidea–Papilionoidea). Rev. Acad. Colomb. Cienc. Exact 2013, 37, 144. [Google Scholar] [CrossRef]
- Molleman, F.; Van Alphen, M.E.; Brakefield, P.M.; Zwann, B.J. Preferences and food quality of fruit-feeding butterflies in Kibale forest, Uganda. Biotropica 2005, 37, 657–663. [Google Scholar] [CrossRef]
- Torres, C.; Osorio-Beristain, M.; Mariano, N.; Legal, L. Sex-dependant seasonal activity patterns among two subfamilies of Nymphalidae (Lepidoptera) in the Mexican tropical dry forest. Ann. Soc. Entomol. Fr. 2009, 45, 265–274. [Google Scholar] [CrossRef]
- Luque, C.; Legal, L.; Staudter, H.; Gers, C.; Wink, M. ISSR (Inter Simple Sequence Repeats) as genetic markers in Noctuids (Lepidoptera, Heterocera, Noctuidae, Noctuinae). Hereditas 2002, 136, 251–253. [Google Scholar] [CrossRef] [PubMed]
- Machkour-M’Rabet, S.; Hénaut, Y.; Charruau, P.; Gevrey, M.; Legal, L. Extinction risk of the American Crocodile in the Mexican Caribbean revealed by ISSR-PCR method: Implication for Conservation. Mar. Biol. 2009, 156, 1321–1333. [Google Scholar] [CrossRef]
- Machkour-M’Rabet, S.; Hénaut, Y.; Calmé, S.; Legal, L. When landscape modification is advantageous for protected species. The case of a synanthropic tarantula, Brachypelma vagans. J. Insect Conserv. 2012, 16, 479–488. [Google Scholar] [CrossRef]
- Coutinho, J.P.; Carvalho, A.; Lima-Brito, J. Genetic diversity assessment and estimation of phylogenetic relationships among 26 Fagaceae species using ISSRs. Biochem. Syst. Ecol. 2014, 54, 247–256. [Google Scholar] [CrossRef]
- Wink, M. Use of ADN markers to study birds migration. J. Ornithol. 2006, 147, 234–244. [Google Scholar] [CrossRef]
- Turlure, C.; Vandewoestijne, S.; Baguette, M. Conservation genetics of a threatened butterfly: Comparison of allozymes, RAPDs and microsatellites. BMC Genet. 2014, 15, 114. [Google Scholar] [CrossRef] [PubMed]
- Sinama, M.; Dubut, V.; Costedoat, C.; Gilles, A.; Junker, M. Challenges of microsatellite development in Lepidoptera: Euphydryas aurinia (Nymphalidae) as a case study. Eur. J. Entomol. 2011, 108, 261–266. [Google Scholar] [CrossRef]
- Luque, C.; Legal, L.; Machkour-M’Rabet, S.; Winterton, P.; Gers, C.; Wink, M. Apparent influences of host-plant distribution on the structure and the genetic variability of local populations of the Purple Clay (Diarsia brunnea). Biochem. Syst. Ecol. 2009, 37, 6–15. [Google Scholar] [CrossRef]
- Hundsdoerfer, A.K.; Wink, M. New Source of Genetic Polymorphisms in Lepidoptera? Z. Naturforsch. C 2005, 60, 618–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayan, K.; Anuradha, H.J.; Nair, C.V.; Pradeep, A.R.; Awasthi, A.; Saratchandra, B.; Rahman, S.A.; Singh, K.C.; Chakraborti, R.; Urs, S.R. Genetic diversity and differentiation among populations of the Indian eri silkworm, Samia cynthia ricini, revealed by ISSR markers. J. Insect Sci. 2006, 6, 30. [Google Scholar] [CrossRef]
- Khurad, A.M.; Kanginakudru, S.; Qureshi, S.O.; Rathod, M.K.; Rai, M.M.; Nagaraju, J. A new Bombyx mori larval ovarian cell line highly susceptible to nucleopolyhedrovirus. J. Invertebr. Pathol. 2006, 92, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Legal, L.; Bermudez-Torres, K.; Leyva Sanchez, E.; Machkour-M’Rabet, S. ISSR, una nueva herramienta para el estudio de especies y poblaciones en el marco de la genética de la conservación. Mesoamericana 2008, 12, 74. [Google Scholar]
- Srivastava, P.P.; Vijayan, K.; Kar, P.K.; Saratchandra, B. Diversity and marker association in tropical silkworm breeds of Bombyx mori (Lepidoptera: Bombycidae). Int. J. Trop. Insect Sci. 2011, 31, 182–191. [Google Scholar] [CrossRef]
- Radjabi, R.; Sarafrazi, A.; Tarang, A.; Kamali, K.; Tirgari, S. Intraspecific Biodiversity of Iranian Local Races of Silkworm Bombyx mori by ISSR (Inter-Simple Sequence Repeat) Molecular Marker. World J. Zool. 2012, 7, 17–22. [Google Scholar]
- Gradish, A.E.; Keyghobadi, N.; Otis, G.W. Population genetic structure and genetic diversity of the threatened White Mountain arctic butterfly (Oeneis melissa semidea). Conserv. Genet. 2015, 16, 1253–1264. [Google Scholar] [CrossRef]
- Mariette, S.; Le Corre, V.; Austerlitz, F.; Kremer, A. Sampling within the genome for measuring within-population diversity: Trade-offs between markers. Mol. Ecol. 2002, 11, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.F.; Anderson, N.O. How many marker loci are necessary? Analysis of dominant marker data sets using two popular population genetic algorithms. Ecol. Evol. 2013, 3, 3455–3470. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Milligan, B.G. Analysis of population genetic structure with RAPD markers. Mol. Ecol. 1994, 3, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GENALEX V6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.; Boyle, T.J.B. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg. J. Bot. 1999, 6, 1234–1236. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction sites. Genetics 1992, 131, 479–491. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P.J. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for STRUCTURE Software; Version 2.3.; University of Chicago: Chicago, IL, USA, 2010. [Google Scholar]
- Minno, M.C.; Emmel, T.C. Butterflies of the Florida Keys; Scientific Publishers: Gainesville, FL, USA, 1993; p. 168. [Google Scholar]
- Valdez-Hernández, M.; Sánchez, O.; Islebe, G.A.; Snook, L.K.; Negreros-Castillo, P. Recovery and early succession after experimental disturbance in a seasonally dry tropical forest in Mexico. For. Ecol. Manag. 2014, 334, 331–343. [Google Scholar] [CrossRef]
- Ribeiro, D.B.; Batista, R.; Prado, P.I.; Brown, K.S., Jr.; Freitas, A.V.L. The importance of small scales to the fruit-feeding butterfly assemblages in a fragmented landscape. Biodivers. Conserv. 2012, 21, 811–827. [Google Scholar] [CrossRef]
- Epps, C.W.; Palsboll, P.J.; Wehausen, J.D.; Roderick, G.K.; Ramey, R.R., II; McCullough, D.R. Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. Ecol. Lett. 2005, 8, 1029–1038. [Google Scholar] [CrossRef]
- Machkour-M’Rabet, S.; Vilchis-Nestor, C.A.; Barriga-Sosa, I.D.L.A.; Legal, L.; Hénaut, Y. A molecular approach to understand the riddle of the invasive success of the tarantula, Brachypelma vagans, on Cozumel Island, Mexico. Biochem. Syst. Ecol. 2017, 70, 260–267. [Google Scholar] [CrossRef]
- Gasmi, S.; Ferval, M.; Pelissier, C.; D’Amico, F.; Maris, T.; Tackx, M.; Legal, L. Genetic diversity among the Eurytemora affinis species complex in the Scheldt estuary and its tributaries using ISSR-PCR marker assay. Estuar. Coast. Shelf Sci. 2014, S145, 22–30. [Google Scholar] [CrossRef]
- Welling, E. Notes on butterflies migration in the Peninsula of Yucatan. J. Lepid. Soc. 1959, 13, 62–64. [Google Scholar]
- Alayo, P.D.; Hernández, L.R. Atlas de Las Mariposas Diurnas de Cuba (Lepidoptera: Rhopalocera); Editorial Científico Técnica la Habana: Habana, Cuba, 1987. [Google Scholar]
- Norris, T. Day 7–22 June–on to Guanahacabibes. Available online: https://www.butterfliesofcuba.com/blog/day-7-22-june-on-to-guanahacabibes (accessed on 1 June 2018).
- Smith, B.T.; Escalante, P.; Hernandez-Baños, B.E.; Navarro-Sigüenza, A.G.; Rohwer, S.; Klicka, J. The role of historical and contemporary processes on phylogeographic structure and genetic diversity in the Northern Cardinal, Cardinalis cardinalis. BMC Evol. Biol. 2011, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.; Seitz, A. Influence of habitat fragmentation on the genetic structure of Polyommatus coridon (Lepidoptera: Lycaenidae): Implications for conservation. Biol. Conserv. 2002, 107, 291–297. [Google Scholar] [CrossRef]
- Exeler, N.; Kratochwil, A.; Hochkirch, A. Does recent habitat fragmentation affect the population genetics of a heathland specialist, Andrena fuscipes (Hymenoptera: Andrenidae)? Conserv. Genet. 2010, 11, 1679–1687. [Google Scholar] [CrossRef]
- Harrison, S. Local extinction in a metapopulation context: An empirical evaluation. Biol. J. Linn. Soc. 1991, 42, 47–88. [Google Scholar] [CrossRef]
- Kitahara, M.; Sei, K.; Fujii, K. Patterns in the structure of grassland butterfly communities along a gradient of human disturbance: Further analysis based on the generalist/specialist concept. Popul. Ecol. 2000, 42, 135–144. [Google Scholar] [CrossRef]
- Brückmann, S.V.; Krauss, J.; Steffan-Dewenter, I. Butterfly and plant specialists suffer from reduced connectivity in fragmented landscapes. J. Appl. Ecol. 2010, 47, 799–809. [Google Scholar] [CrossRef]
- Warren, M.S.; Hill, J.K.; Thomas, J.A.; Asher, J.; Fox, R. Rapid responses of British butterflies to opposing forces of climate and habitat change. Nature 2001, 414, 65–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beccaloni, G.W.; Viloria, A.L.; Hall, S.K.; Robinson, G.S. Catalogue of Hostplants of the Neotropical Butterflies; Sociedad Entomológica Aragonesa: Zaragosa, Spain, 2008. [Google Scholar]
- Exeler, N.; Kratochwil, A.; Hochkirch, A. Strong genetic exchange among populations of a specialist bee, Andrena vaga (Hymenoptera: Andrenidae). Conserv. Genet. 2008, 9, 1233–1241. [Google Scholar] [CrossRef]
- Trochet, A.; Legrand, D.; Larranaga, N.; Ducatez, S.; Calvez, O.; Cote, J.; Clobert, J.; Baguette, M. Population sex ratio and dispersal in experimental, two-patch metapopulations of butterflies. J. Anim. Ecol. 2013, 82, 946–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.K.; Hughes, C.L.; Dytham, C.; Searle, J.B. Genetic diversity in butterflies: Interactive effects of habitat fragmentation and climate-driven range expansion. Biol. Lett. 2006, 2, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Benedick, S.; White, T.A.; Searle, J.B.; Hamer, K.C.; Mustaffa, N. Impacts of habitat fragmentation on genetic diversity in a tropical forest butterfly on Borneo. J. Trop. Ecol. 2007, 23, 623–634. [Google Scholar] [CrossRef]
- Dover, J.; Settele, J. The influences of landscape structure on butterfly distribution and movement: A review. J. Insect Conserv. 2009, 13, 3–27. [Google Scholar] [CrossRef]
- Krauss, J.; Schmitt, T.; Seitz, A.; Steffan-Dewenter, I.; Tscharntke, T. Effects of habitat fragmentation on the genetic structure of the monophagous butterfly Polyommatus coridon along its northern range margin. Mol. Ecol. 2004, 13, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Pozo, C.; (El Colegio de la Frontera Sur, Chetumal, Quintana Roo, México). Personal Communication, 2018.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calakmul (CK) | Kohunlich (KH) | Miguel Hidalgo (MH) | Jardín Botánico (JB) | Xcalak (XC) | Kiuic (KU) | Sian Ka’an (SK) | Zapotal (ZP) | ||
|---|---|---|---|---|---|---|---|---|---|
| Sampling Data | N1 | 68 | 38 | 17 | 36 | 32 | 59 | 50 | 50 |
| N2 | 58 | 38 | 17 | 36 | 31 | 54 | 40 | 49 | |
| Genetic Data | TB | 106 | 98 | 94 | 94 | 95 | 108 | 112 | 104 |
| RB | 17 | 8 | 0 | 9 | 11 | 15 | 5 | 22 | |
| PB | 0 | 1 | 0 | 0 | 0 | 2 | 2 | 0 | |
| PPL | 89 | 81 | 76 | 77 | 77 | 90 | 92 | 85 | |
| He (±SE) | 0.194 (0.015) | 0.195 (0.016) | 0.202 (0.016) | 0.187 (0.016) | 0.189 (0.016) | 0.205 (0.016) | 0.220 (0.014) | 0.182 (0.016) | |
| Ecological Data | dtrees | 0.05 | 0.03 | 0.05 | 0.07 | 0.04 | 0.08 | 0.07 | 0.06 |
| dshrubs | 0.81 | 1.11 | 1.37 | 0.87 | 1.23 | 0.89 | 0.83 | 1.11 | |
| dherbs | 11.38 | 21.25 | 12.77 | 22.33 | 18.85 | 40.63 | 40.12 | 20.17 | |
| Patch size (km2) | 7231 | 0.085 | 23 | 0.65 | 45 | 16.5 | 3750 | 24 | |
| Climatic Data | Temperature (°C) | 25.14 | 27.01 | 27.01 | 26.79 | 27.67 | 25.51 | 26.77 | 26.48 |
| Humidity (%) | 81.04 | 81.53 | 81.53 | 73.42 | 78.82 | 79.14 | 78.94 | 75.75 | |
| Precipitation (mm) | 3.66 | 5.79 | 5.79 | 6.77 | 6.13 | 3.19 | 7.04 | 1.17 | |
| Land cover Data | Forest | 98% | 93% | 79% | 24% | 13% | 100% | 100% | 98% |
| Coast | 0% | 0% | 0% | 6% | 87% | 0% | 0% | 0% | |
| Anthropogenic | 2% | 7% | 21% | 70% | 0% | 0% | 0% | 2% |
| Abbreviation | % GC | Tm | Ta | N Bands | Size (pb) |
|---|---|---|---|---|---|
| (AG)8YC | 52.8 | 59.5 | 58 | 37 | 200–1600 |
| (GAG)5GC | 70.6 | 64.8 | 63 | 36 | 400–1400 |
| (GA)8C | 52.9 | 56 | 54 | 20 | 150–1200 |
| (AC)8C | 52.9 | 62.3 | 61 | 26 | 300–1500 |
| CK | KH | MH | JB | XC | KU | SK | ZP | |
|---|---|---|---|---|---|---|---|---|
| CK | 0.057 | 0.095 | 0.042 | 0.038 | 0.040 | 0.042 | 0.067 | |
| KH | 115 | 0.057 | 0.036 | 0.021 | 0.043 | 0.042 | 0.050 | |
| MH | 162 | 69 | 0.053 | 0.056 | 0.073 | 0.051 | 0.085 | |
| JB | 325 | 332 | 264 | 0.026 | 0.044 | 0.053 | 0.067 | |
| XC | 217 | 102 | 93 | 306 | 0.034 | 0.023 | 0.030 | |
| KU | 176 | 200 | 178 | 290 | 271 | 0.018 | 0.030 | |
| SK | 242 | 196 | 126 | 136 | 176 | 212 | 0.032 | |
| ZP | 390 | 342 | 285 | 100 | 355 | 244 | 175 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavanzón-Medrano, L.E.; Machkour-M’Rabet, S.; Chablé-Iuit, L.R.; Pozo, C.; Hénaut, Y.; Legal, L. Effect of Climatic Conditions and Land Cover on Genetic Structure and Diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico. Diversity 2018, 10, 79. https://doi.org/10.3390/d10030079
Cavanzón-Medrano LE, Machkour-M’Rabet S, Chablé-Iuit LR, Pozo C, Hénaut Y, Legal L. Effect of Climatic Conditions and Land Cover on Genetic Structure and Diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico. Diversity. 2018; 10(3):79. https://doi.org/10.3390/d10030079
Chicago/Turabian StyleCavanzón-Medrano, Laura Elena, Salima Machkour-M’Rabet, Landy Rubí Chablé-Iuit, Carmen Pozo, Yann Hénaut, and Luc Legal. 2018. "Effect of Climatic Conditions and Land Cover on Genetic Structure and Diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico" Diversity 10, no. 3: 79. https://doi.org/10.3390/d10030079
APA StyleCavanzón-Medrano, L. E., Machkour-M’Rabet, S., Chablé-Iuit, L. R., Pozo, C., Hénaut, Y., & Legal, L. (2018). Effect of Climatic Conditions and Land Cover on Genetic Structure and Diversity of Eunica tatila (Lepidoptera) in the Yucatan Peninsula, Mexico. Diversity, 10(3), 79. https://doi.org/10.3390/d10030079