Reproductive Ecology of Wrinkle-Lipped Free-Tailed Bats Chaerephon plicatus (Buchannan, 1800) in Relation to Guano Production in Cambodia


Abstract
:1. Introduction
2. Materials and Methods
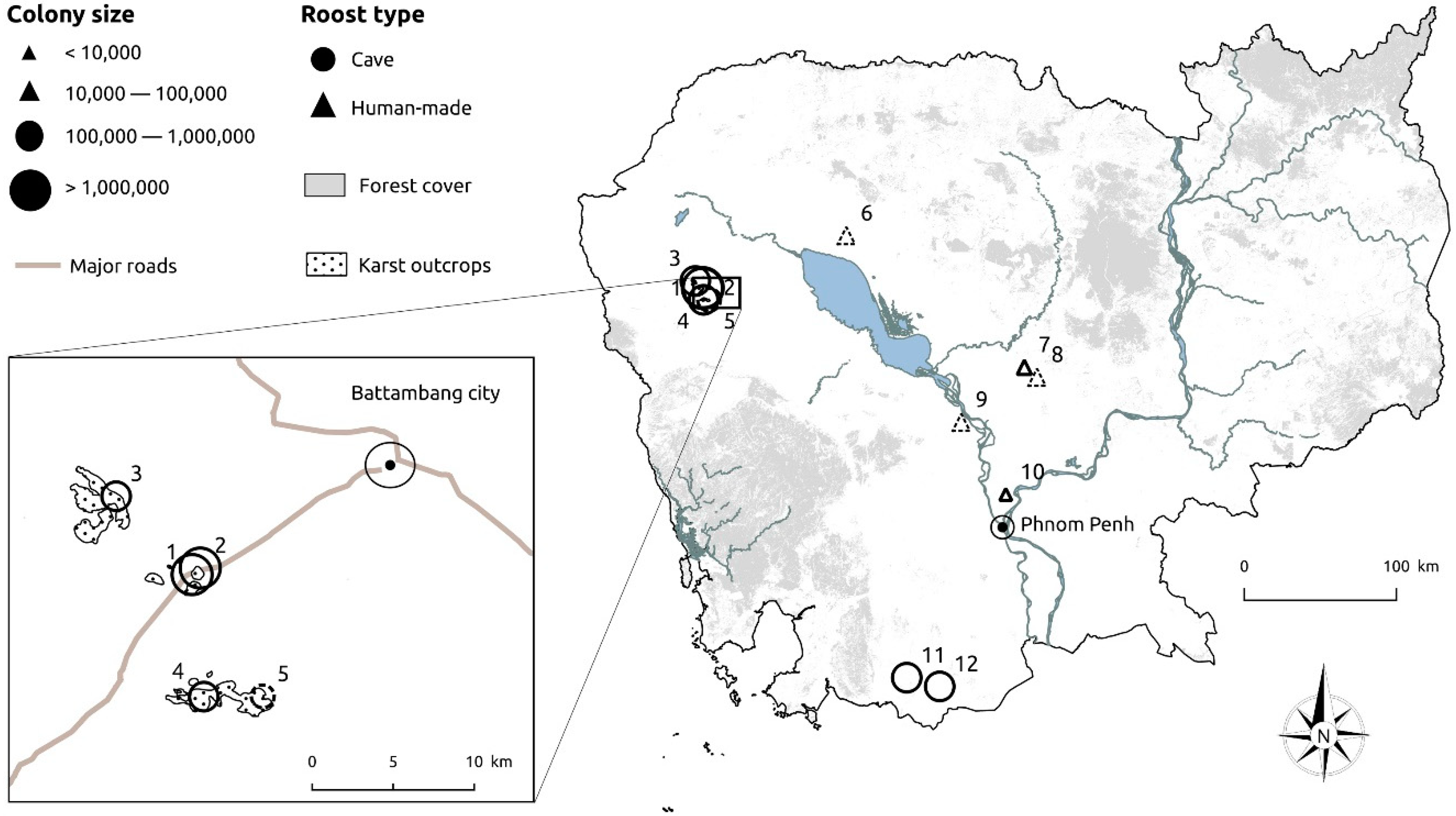
2.1. Study Sites
2.2. Data Collection
2.3. Reproductive Assessment
2.4. Analysis
3. Results
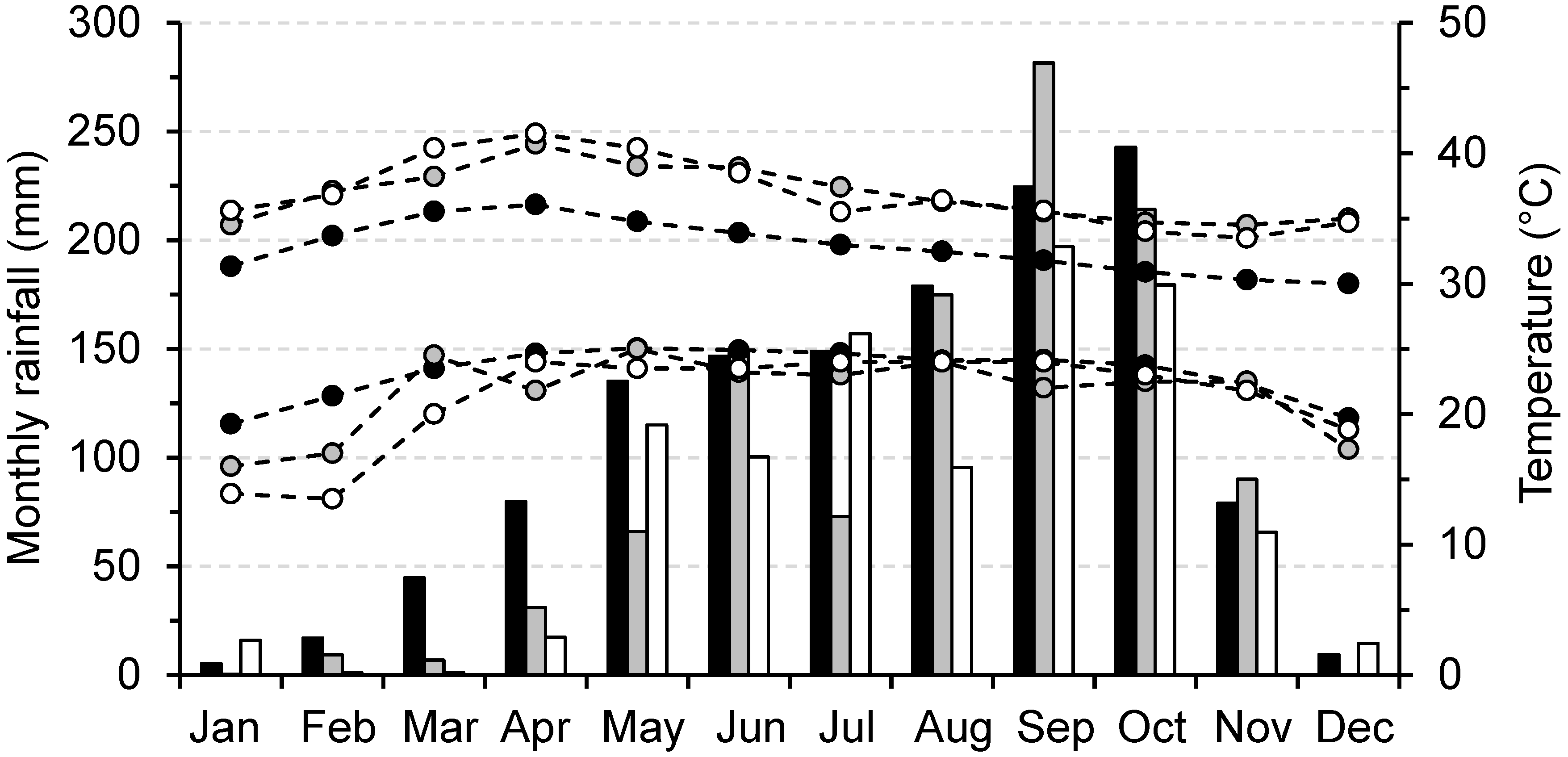
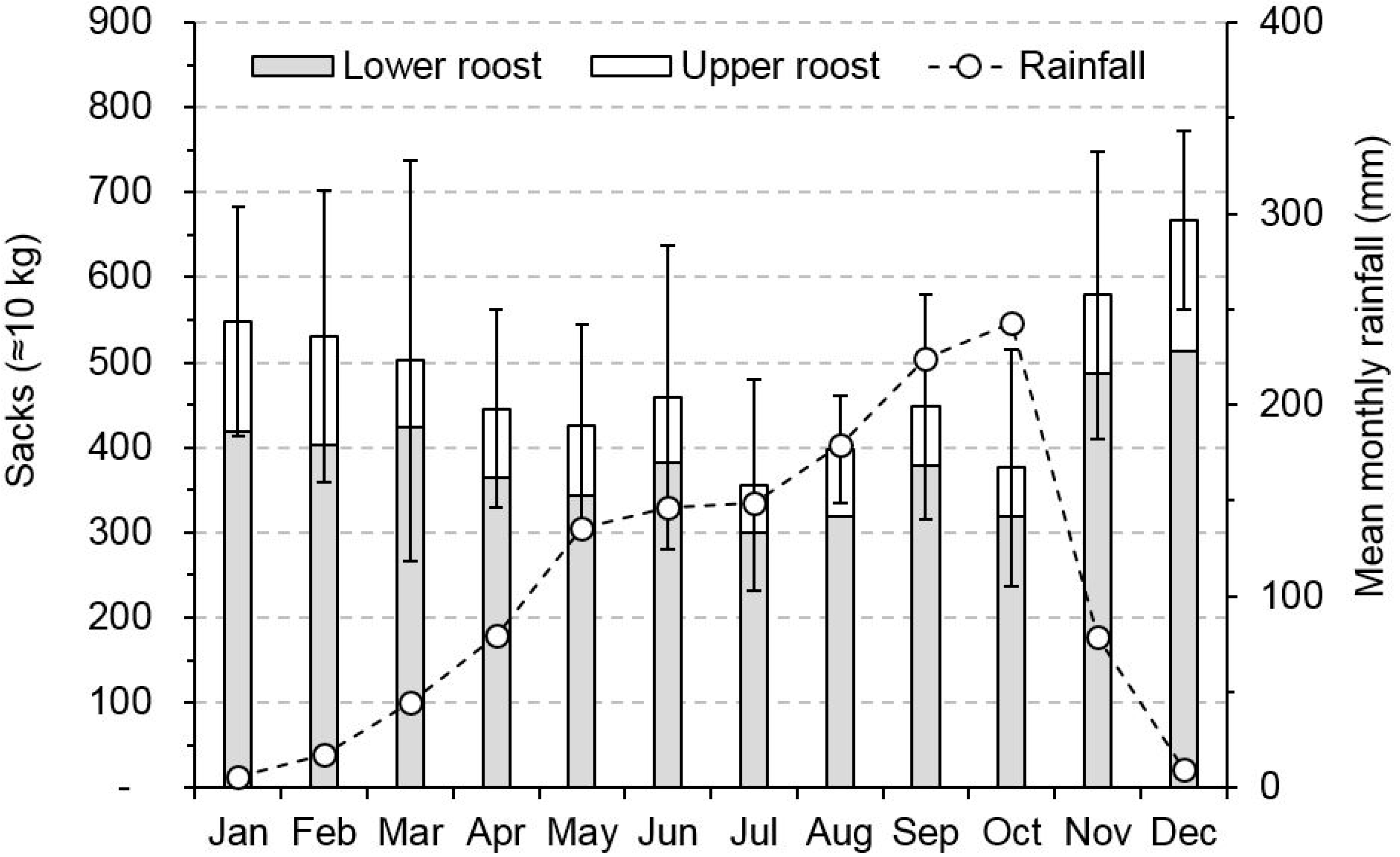
3.1. Climatic Variation
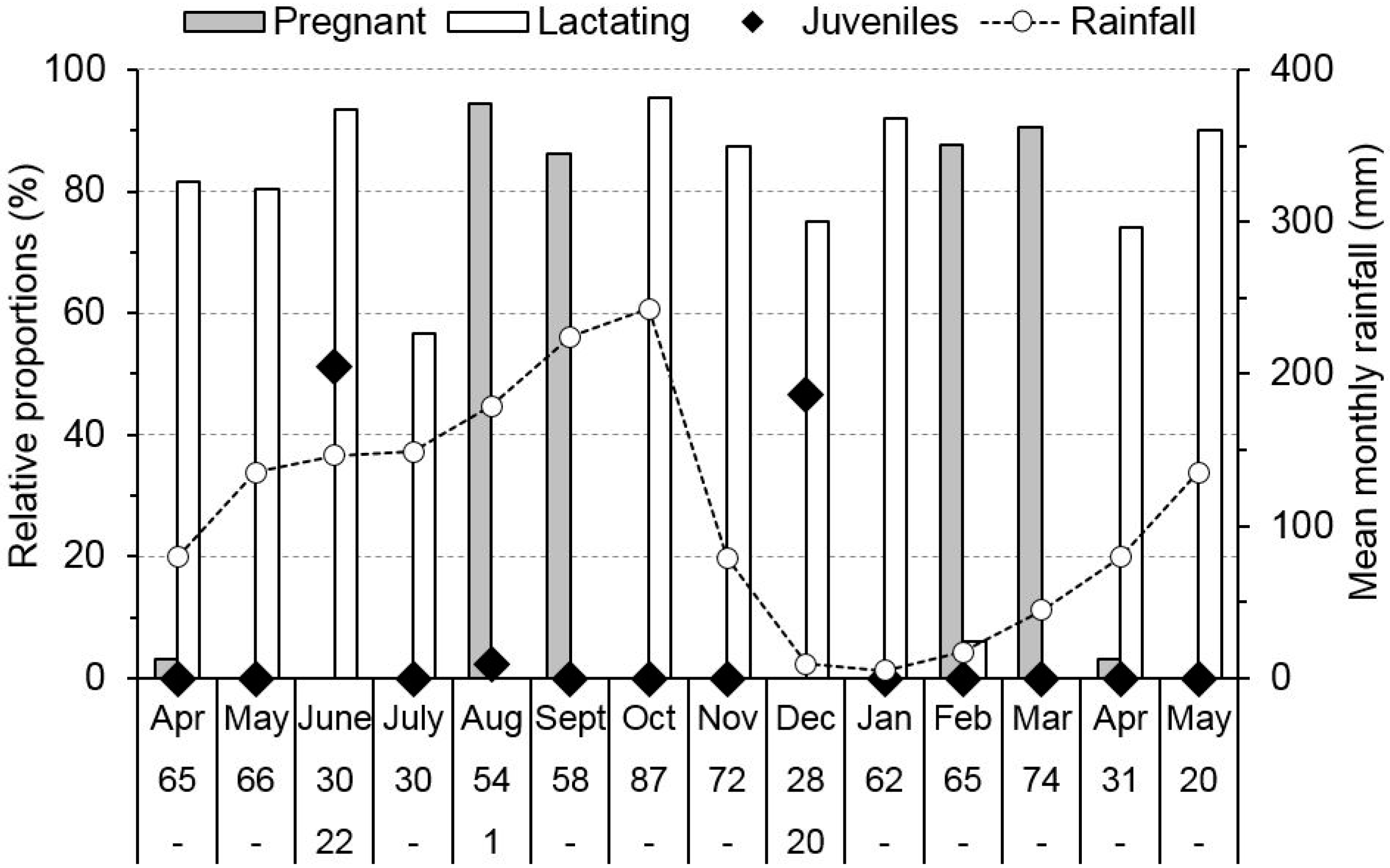
3.2. Reproductive Phenology
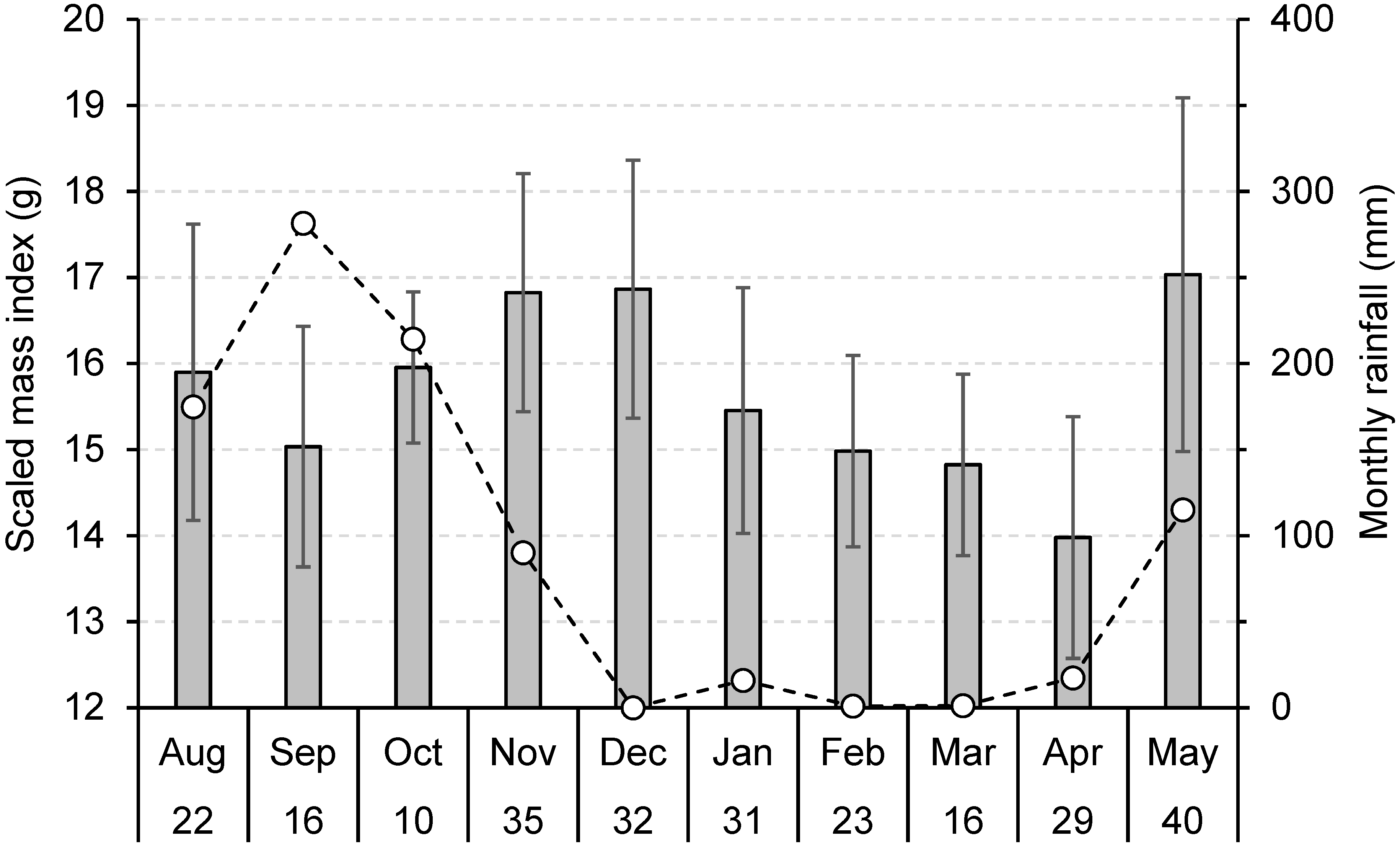
3.3. Seasonal Body Condition
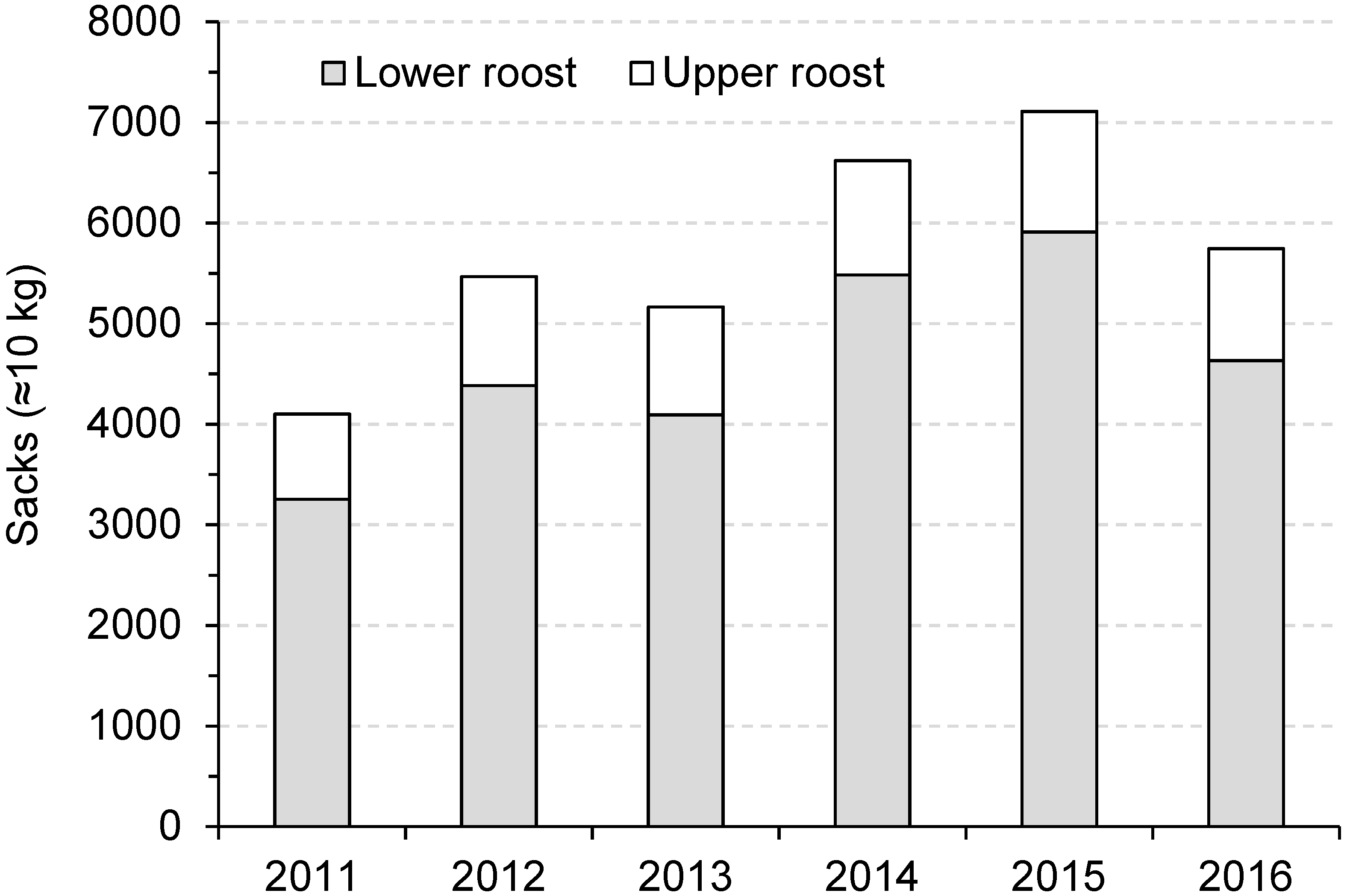
3.4. Guano Harvests
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kingston, T. Response of bat diversity to forest disturbance in Southeast Asia—Insights from long term research in Malaysia. In Bat Evolution, Ecology, and Conservation; Adams, R.A., Pedersen, S.C., Eds.; Springer: New York, NY, USA, 2013; pp. 169–185. ISBN 978-1-4614-7396-1. [Google Scholar]
- Sodhi, N.S.; Koh, L.P.; Clements, R.; Wanger, T.C.; Hill, J.K.; Hamer, K.C.; Clough, Y.; Tscharntke, T.; Posa, M.R.C.; Lee, T.M. Conserving Southeast Asian forest biodiversity in human-modified landscapes. Biol. Conserv. 2010, 143, 2375–2384. [Google Scholar] [CrossRef]
- Jones, G.W. The Population of Southeast Asia; Working Paper Series No. 196; Asia Research Institute: Singapore, 2013; pp. 1–39. [Google Scholar]
- Furey, N.M.; Racey, P.A. Conservation ecology of cave bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer: Basel, Switzerland, 2016; pp. 463–500. ISBN 978-3-319-25218-6. [Google Scholar]
- Simmons, N.B. Order Chiroptera. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 312–529. ISBN 0-8018-8221-4. [Google Scholar]
- Bates, P.J.J.; Harrison, D.L. Bats of the Indian Subcontinent; Harrison Zoological Museum: Kent, UK, 1997; pp. 1–258. ISBN 0-9517313-1-9. [Google Scholar]
- Wanger, T.C.; Darras, K.; Bumrungsri, S.; Tscharntke, T.; Klein, A.M. Bat pest control contributes to food security in Thailand. Biol. Conserv. 2014, 171, 220–223. [Google Scholar] [CrossRef]
- Cleveland, C.J.; Betke, M.; Federico, P.; Frank, J.D.; Hallam, T.G.; Horn, J.; López, J.D., Jr.; McCracken, G.F.; Medellín, R.A.; Moreno-Valdez, A.; et al. Economic value of the pest control service provided by Brazilian free-tailed bats in south-central Texas. Front. Ecol. Environ. 2006, 4, 238–243. [Google Scholar] [CrossRef]
- Estimating the Economic Value of Insect-Eating Bats: A Case Study from Cambodia. Available online: http://www.batcon.org/resources/media-education/bats-magazine/bat_article/846 (accessed on 10 April 2018).
- Bats Missing from the Museum. The Cambodia Daily, 20 March 2002. Available online: https://www.cambodiadaily.com/news/bats-missing-from-museum-30558/ (accessed on 10 April 2018).
- Yim, S.; Mackie, I. Status and conservation of wrinkle-lipped bat Tadarida plicata in Cambodia. In Proceedings of the International Meeting on “Increasing In-Country Capacity and Regional Co-Operation to Promote Bat Conservation in Cambodia, with Particular Reference to Otomops wroughtoni”, Phnom Penh, Cambodia, 26–27 October 2009. [Google Scholar]
- Furey, N.M.; Whitten, T.; Cappelle, J.; Racey, P.A. The conservation status of Cambodian cave bats. In International Speleological Project to Cambodia 2016 (Provinces of Stoeng Treng, Kampong Speu, Banteay Meanchey and Battambang); Laumanns, M., Ed.; Berliner Höhlenkundliche Berichte: Berlin, Germany, 2016; Volume 64, pp. 82–95, 1617–8572. [Google Scholar]
- Racey, P.A.; Entwistle, A.C. Conservation ecology of bats. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 680–743. ISBN 0-22604620604. [Google Scholar]
- Racey, P.A.; Entwistle, A.C. Life history and reproductive strategies in bats. In Reproductive Biology of Bats; Crighton, E.G., Krutzsch, P.H., Eds.; Academic Press: New York, NY, USA, 2000; pp. 363–414. ISBN 0-12-195670-9. [Google Scholar]
- Goh, K.C. The climate of Southeast Asia. In The Physical Geography of Southeast Asia; Gupta, A., Ed.; Oxford University Press: Oxford, UK, 2005; pp. 80–93. ISBN 0-19-924802-8. [Google Scholar]
- Hillman, A. The study on wrinkled-lipped free-tailed bats (Tadarida plicata) at Khao Chong Pran Non-hunting Area, Ratchaburi Province. R. For. Dep. J. 1999, 1, 72–83. [Google Scholar]
- Aye, N.N. Ecology and Economic Importance of Tadarida plicata (Buchannan, 1800) Free-Tailed Bat in Some Areas of Myanmar. Ph.D. Thesis, University of Yangon, Yangon, Myanmar, 2006. [Google Scholar]
- Leelapaibul, W.; Bumrungsri, S.; Pattanawiboon, A. Diet of wrinkle-lipped free-tailed bat (Tadarida plicata Buchannan, 1800) in central Thailand: Insectivorous bats potentially act as biological pest control agents. Acta Chiropterol. 2005, 7, 111–119. [Google Scholar] [CrossRef]
- McWilliam, A.N. Adaptive Responses to Seasonality in Four Species of Microchiroptera in Coastal Kenya. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 1982. [Google Scholar]
- Nurul-Ain, E.; Rosli, H.; Kingston, T. Resource availability and roosting ecology shape reproductive phenology of rainforest insectivorous bats. Biotropica 2017, 49, 382–394. [Google Scholar] [CrossRef]
- Laumanns, M. International Speleological Project to Cambodia 2008 (Battambang Area); Berliner Höhlenkundliche Berichte: Berlin, Germany, 2009; Volume 34, pp. 1–69, 1617–8572. [Google Scholar]
- Chheng, C.; Ta Rumm #1 Cave Concession Holder, Phnom Sampeu, Battambang Province, Cambodia. Personal communication, 2016.
- Lunn, B.; Phnom Romseisak Community Forest Chief, Bobh Village, Bannon District, Battambang Province, Cambodia. Personal communication, 2016.
- Sikes, R.S.; Gannon, W.L.; Animal Care and Use Committee of the American Society of Mammalogists. Guidelines of the American Society of Mammalogists for the use of wild animals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Francis, C.M. A Guide to the Mammals of Southeast Asia; Princeton University Press: Princeton, NJ, USA, 2008; pp. 1–392. ISBN 978-0-691-13551-9. [Google Scholar]
- Kruskop, S. Bats of Vietnam, Checklist and an Identification Manual, 2nd ed.; KMK Scientific Press: Moscow, Russia, 2013; pp. 1–299. ISBN 978-5-87317-901-5. [Google Scholar]
- Anthony, E.L.P. Age determination in bats. In Ecological and Behavioral Methods for the Study of Bats; Kunz, T.H., Ed.; Smithsonian Press: Washington, DC, USA, 1988; pp. 47–58. ISBN 0874745969. [Google Scholar]
- Racey, P.A. Reproductive assessment. In Behavioural and Ecological Methods for the Study of Bats, 2nd ed.; Kunz, T.H., Parsons, S., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2009; pp. 249–264. ISBN 978-0-8018-9147-2. [Google Scholar]
- Furey, N.M.; Mackie, I.J.; Racey, P.A. Reproductive phenology of bat assemblages in Vietnamese karst and its conservation implications. Acta Chiropterol. 2011, 13, 341–354. [Google Scholar] [CrossRef]
- Peig, J.; Green, A.J. New perspectives for estimating body condition from mass/length data: The scaled mass index as an alternative method. Oikos 2009, 118, 1883–1891. [Google Scholar] [CrossRef]
- Thomas, N.M.; Duckworth, J.W.; Doaungboubpha, B.; Williams, M.; Francis, C.M. A checklist of bats (Mammalia: Chiroptera) from Lao PDR. Acta Chiropterol. 2013, 15, 193–260. [Google Scholar] [CrossRef]
- Csorba, G.; Bumrungsri, S.; Francis, C.; Bates, P.; Ong, P.; Gumal, M.; Kingston, T.; Heaney, L.; Balete, D.; Molur, S.; et al. Chaerephon plicatus. The IUCN Red List of Threatened Species. 2014. Available online: http://www.iucnredlist.org/details/4316/0 (accessed on 20 April 2018). [CrossRef]
- Lane, D.J.; Kingston, T.; Lee, B.P.H. Dramatic decline in bat species richness in Singapore, with implications for Southeast Asia. Biol. Conserv. 2006, 131, 584–593. [Google Scholar] [CrossRef]
- Lim, T.; Cappelle, J.; Hoem, T.; Furey, N.M. Insectivorous bat reproduction and human cave visitation in Cambodia: A perfect conservation storm? PLoS ONE 2018, 13, e0196554. [Google Scholar] [CrossRef] [PubMed]
- IUCN-SSC. IUCN-SSC Guidelines for Minimizing the Negative Impact to Bats and Other Cave Organisms from Guano Harvesting, Ver. 1.0; IUCN Species Survival Commission: Gland, Switzerland, 2014; pp. 1–12. [Google Scholar]
- Ades, G.W.J.; Dudgeon, D. Insect seasonality in Hong Kong: A monsoonal environment in the northern tropics. Mem. Hong Kong Natl. Hist. Soc. 1999, 22, 81–97. [Google Scholar]
- Kai, K.H.; Corlett, R.T. Seasonality of forest invertebrates in Hong Kong, South China. J. Trop. Ecol. 2002, 18, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Heat Wave Blamed as Hundreds of Bats Die in Siem Reap. The Phnom Penh Post, 22 April 2016. Available online: https://www.phnompenhpost.com/national/heat-wave-blamed-hundreds-bats-die-siem-reap (accessed on 10 April 2018).
- Scorching Hot Season Sets New All-Time Highs. The Phnom Penh Post, 21 April 2016. Available online: https://www.phnompenhpost.com/national/scorching-hot-season-sets-new-all-time-highs (accessed on 10 April 2018).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Site Name (Hill, Province) | Latitude, Longitude | Roost Type/Status | Colony Size 1 |
|---|---|---|---|---|
| 1 | Ta Rumm #1 2 (Sampeu hill, Battambang) | 13.022 N, 103.095 E | Cave/Permanent | 1,800,000 |
| 2 | La Ang Pracheu 2 (Sampeu hill, Battambang) | 13.026 N, 103.099 E | Cave/Permanent | 1,000,000 |
| 3 | La Ang Prakiu #2 2 (Takriem hill, Battambang) | 13.066 N, 103.051 E | Cave/Permanent | 800,000 |
| 4 | La Ang Pracheav 2 (Reichiatra hill, Battambang) | 12.954 N, 103.102 E | Cave/Permanent | 600,000 |
| 5 | La Ang Sankeur 2 (Sankeur hill, Battambang) | 12.955 N, 103.135 E | Cave/Seasonal | 900,000 |
| 6 | Ba Kang 3 (Siem Reap) | 13.341 N, 103.966 E 4 | Pagoda/Seasonal | 30,000 |
| 7 | Kampong Thma 3 (Kampong Thom) | 12.560 N, 105.047 E 4 | Pagoda/Permanent | 80,000 |
| 8 | Tang Krosang 3 (Kampong Thom) | 12.501 N, 105.124 E 4 | Pagoda/Seasonal | 70,000 |
| 9 | Yeay Tep 3 (Kampong Chhnang) | 12.233 N, 104.665 E 4 | Pagoda/Seasonal | 10,000 |
| 10 | Unnamed site 3 (Kandal) | 11.804 N, 104.935 E 4 | Stupa/Permanent | 4000 |
| 11 | Loang 2 (Loang hill, Kampot) | 10.714 N, 104.339 E | Cave/Permanent | 900,000 |
| 12 | Vihear Luong 2 (Koun Sat hill, Kampot) | 10.662 N, 104.538 E | Cave/Permanent | 370,000 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furey, N.M.; Racey, P.A.; Ith, S.; Touch, V.; Cappelle, J. Reproductive Ecology of Wrinkle-Lipped Free-Tailed Bats Chaerephon plicatus (Buchannan, 1800) in Relation to Guano Production in Cambodia. Diversity 2018, 10, 91. https://doi.org/10.3390/d10030091
Furey NM, Racey PA, Ith S, Touch V, Cappelle J. Reproductive Ecology of Wrinkle-Lipped Free-Tailed Bats Chaerephon plicatus (Buchannan, 1800) in Relation to Guano Production in Cambodia. Diversity. 2018; 10(3):91. https://doi.org/10.3390/d10030091
Chicago/Turabian StyleFurey, Neil M., Paul A. Racey, Saveng Ith, Van Touch, and Julien Cappelle. 2018. "Reproductive Ecology of Wrinkle-Lipped Free-Tailed Bats Chaerephon plicatus (Buchannan, 1800) in Relation to Guano Production in Cambodia" Diversity 10, no. 3: 91. https://doi.org/10.3390/d10030091
APA StyleFurey, N. M., Racey, P. A., Ith, S., Touch, V., & Cappelle, J. (2018). Reproductive Ecology of Wrinkle-Lipped Free-Tailed Bats Chaerephon plicatus (Buchannan, 1800) in Relation to Guano Production in Cambodia. Diversity, 10(3), 91. https://doi.org/10.3390/d10030091