A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
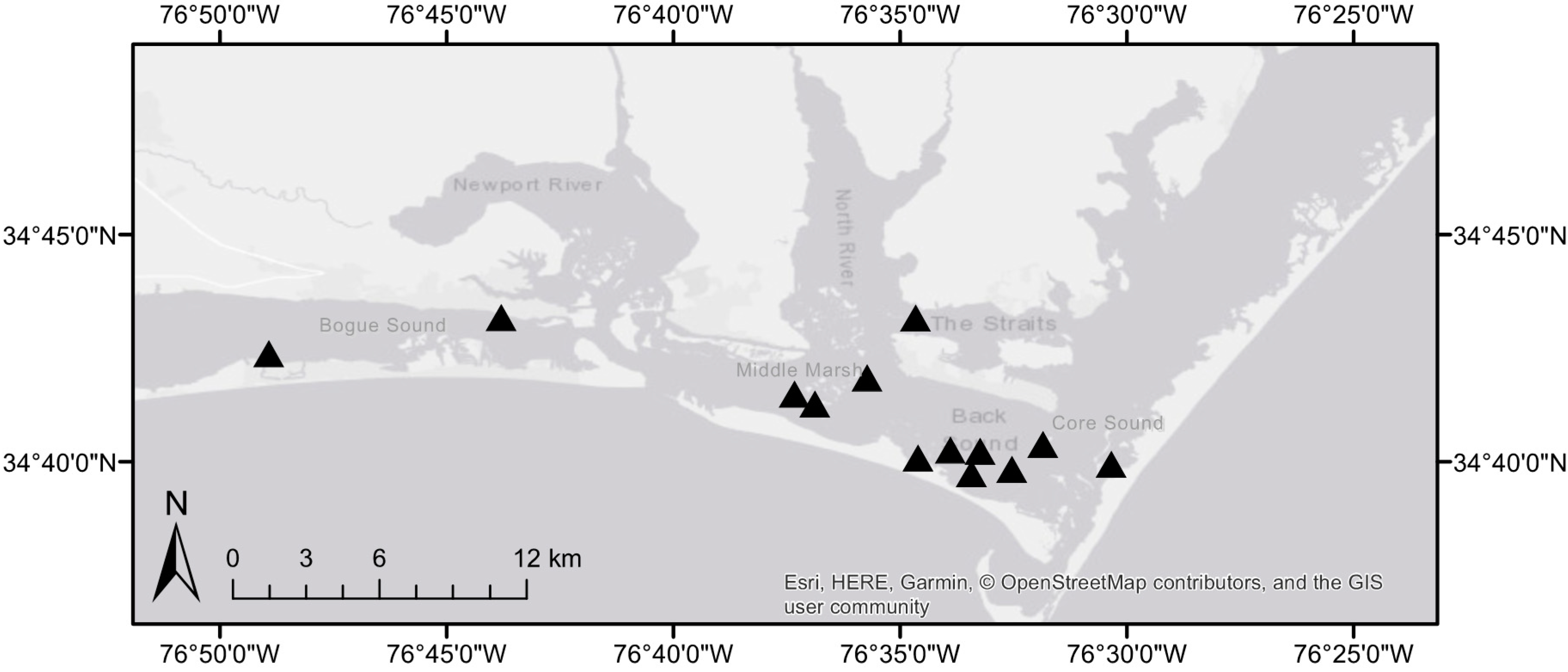
2. Materials and Methods
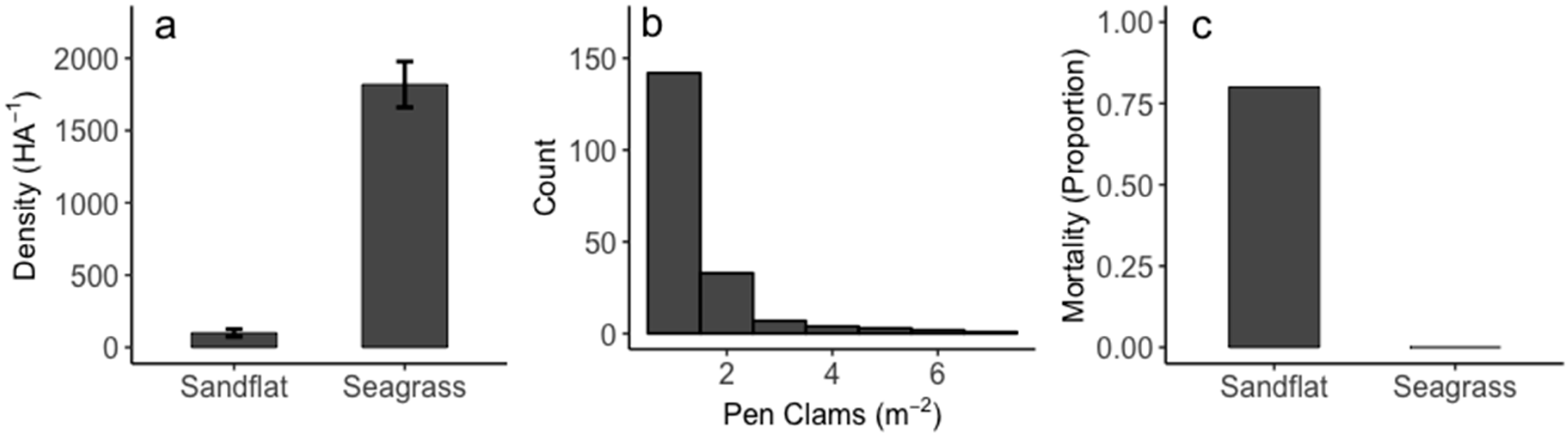
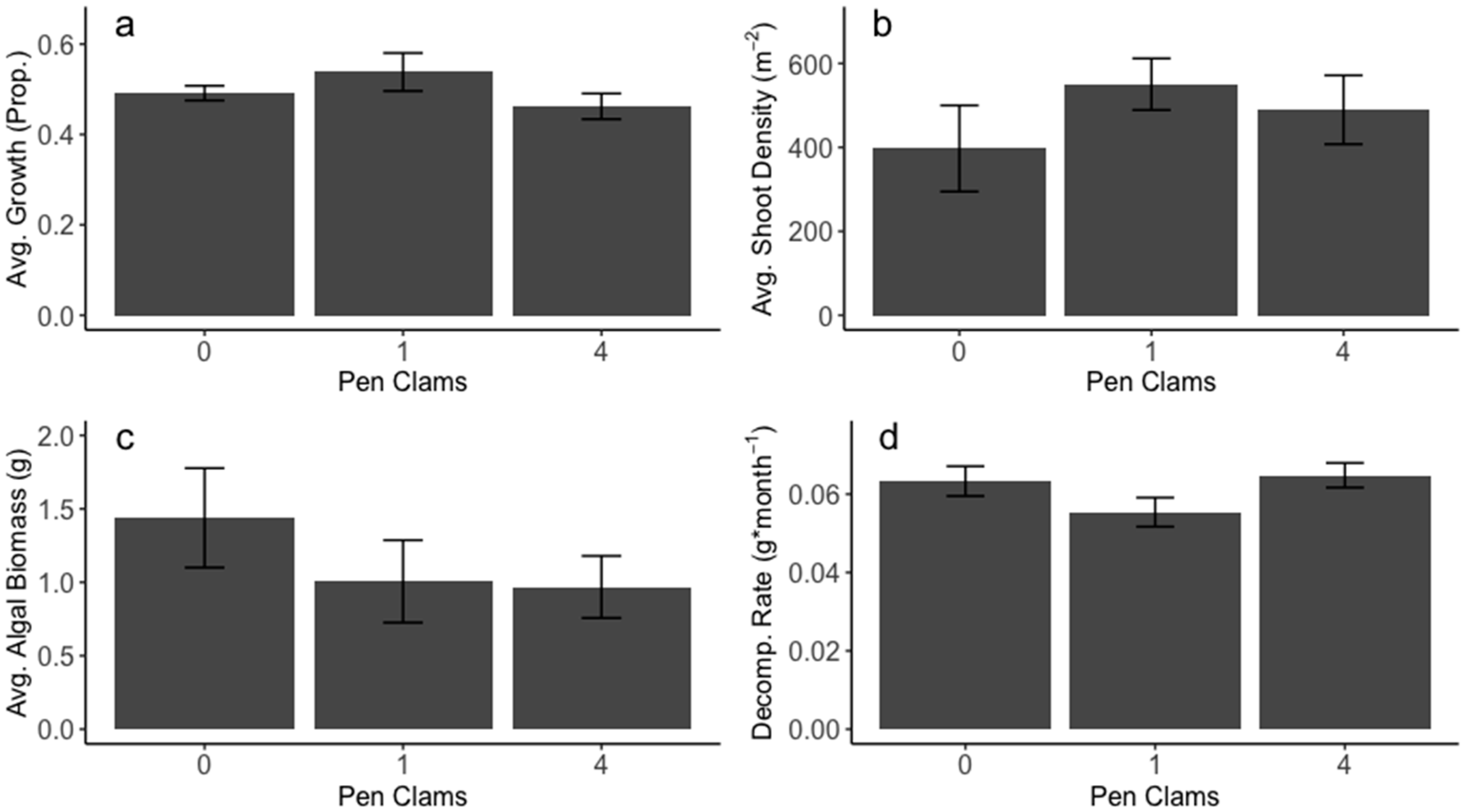
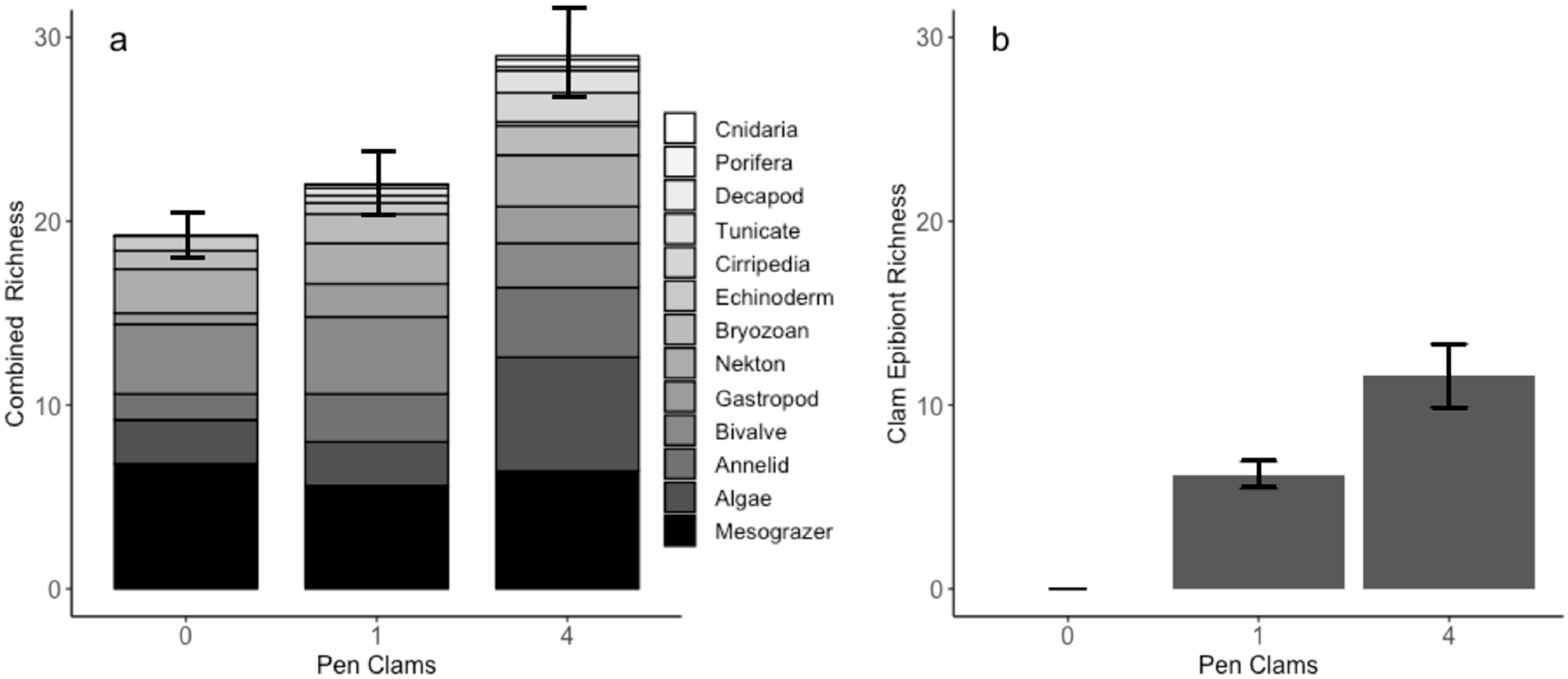
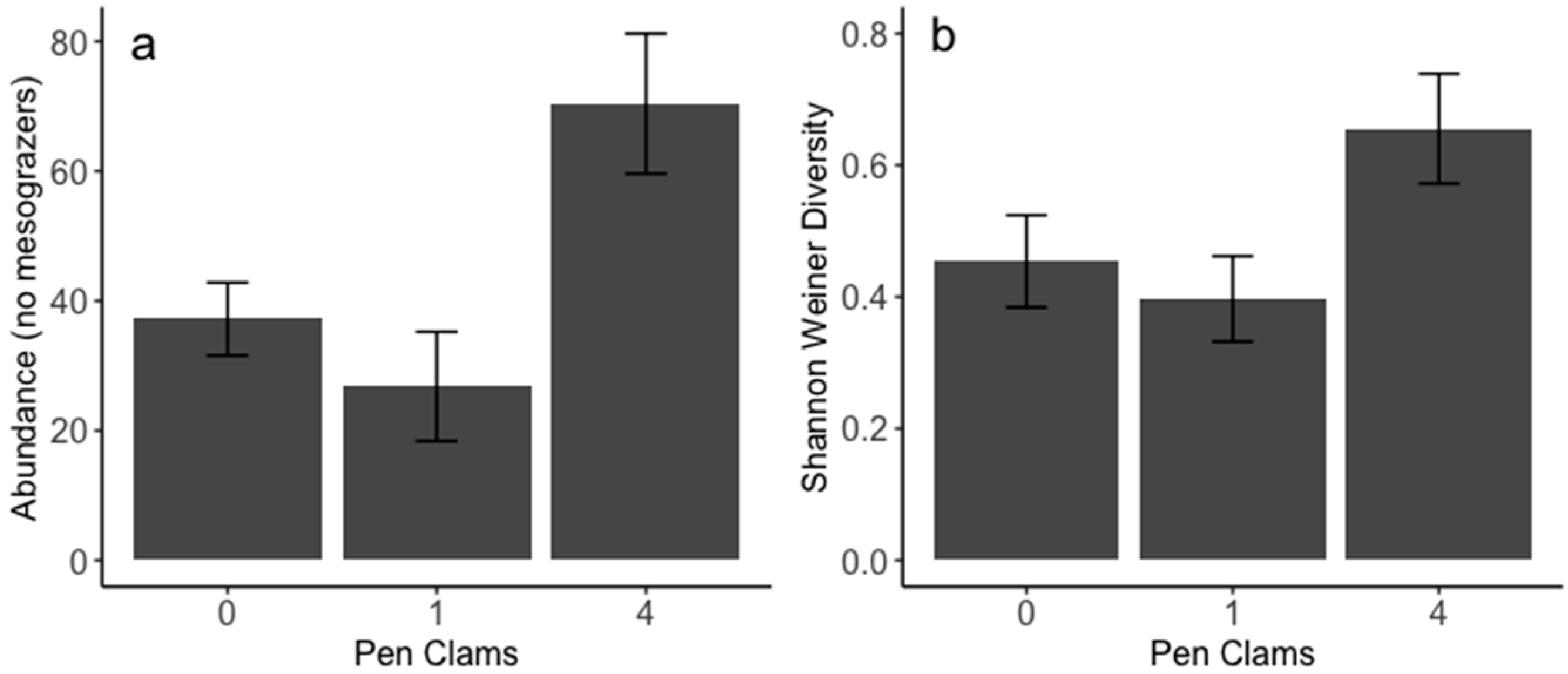
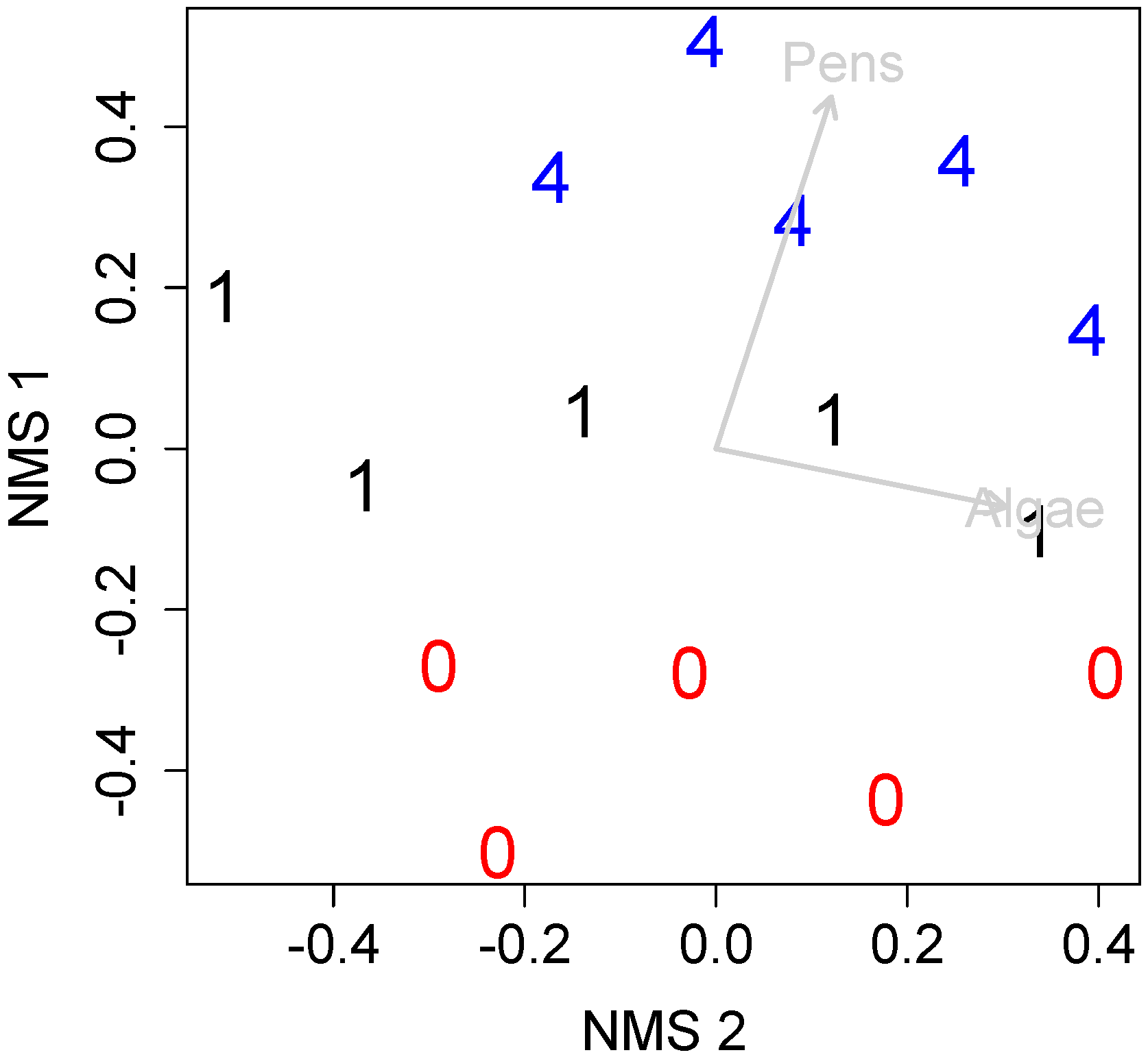
3. Results
4. Discussion
4.1. A Facilitation Cascade Enhances Local Biodiversity
4.2. Effects of Functional Diversity versus Diversity per Se
4.3. Bivalves as Key Drivers of Seagrass Structure and Function
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hairston, N.G.; Smith, F.E.; Slobodkin, L.B. Community structure, population control, and competition. Am. Nat. 1960, 94, 421–425. [Google Scholar] [CrossRef]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Estes, J.A.; Duggins, D.O. Sea otters and kelp forests in Alaska: Generality and variation in a community ecological paradigm. Ecol. Monogr. 1995, 65, 75–100. [Google Scholar] [CrossRef]
- Terborgh, J.; Lopez, L.; Nuñez, P.; Rao, M.; Shahabuddin, G.; Orihuela, G.; Riveros, M.; Ascanio, R.; Adler, G.H.; Lambert, T.D.; et al. Ecological meltdown in predator-free forest fragments. Science 2001, 294, 1923–1926. [Google Scholar] [CrossRef] [PubMed]
- Silliman, B.R.; Zieman, J.C. Top-down control of Spartina alterniflora production by periwinkle grazing in a Virginia salt marsh. Ecology 2001, 82, 2830–2845. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; Altieri, A.; Tuya, F.; Gulbransen, D.; McGlathery, K.J.; Holmer, M.; Silliman, B.R. Habitat cascades: The conceptual context and global relevance of facilitation cascades via habitat formation and modification. Integr. Comp. Biol. 2010, 50, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Hierarchical organization via a facilitation cascade in intertidal cordgrass bed communities. Am. Nat. 2007, 169, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Angelini, C.; Altieri, A.H.; Silliman, B.R.; Bertness, M.D. Interactions among foundation species and their consequences for community organization, biodiversity, and conservation. BioScience 2011, 61, 782–789. [Google Scholar] [CrossRef]
- Angelini, C.; Silliman, B.R. Secondary foundation species as drivers of trophic and functional diversity: Evidence from a tree–epiphyte system. Ecology 2014, 95, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Halpern, B.S.; Silliman, B.R.; Olden, J.D.; Bruno, J.P.; Bertness, M.D. Incorporating positive interactions in aquatic restoration and conservation. Front. Ecol. Environ. 2007, 5, 153–160. [Google Scholar] [CrossRef]
- Angelini, C.; van der Heide, T.; Griffin, J.N.; Morton, J.P.; Derksen-Hooijberg, M.; Lamers, L.P.; Smolders, A.J.; Silliman, B.R. Foundation species’ overlap enhances biodiversity and multifunctionality from the patch to landscape scale in southeastern United States salt marshes. Proc. R. Soc. B 2015, 282, 20150421. [Google Scholar] [CrossRef] [PubMed]
- Bishop, M.J.; Byers, J.E.; Marcek, B.J.; Gribben, P.E. Density-dependent facilitation cascades determine epifaunal community structure in temperate Australian mangroves. Ecology 2012, 93, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Farnsworth, E.J.; Twilley, R.R. Facultative mutualism between red mangroves and root-fouling sponges in Belizean mangal. Ecology 1996, 77, 175–188. [Google Scholar] [CrossRef]
- He, Q.; Bertness, M.D.; Altieri, A.H. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 2013, 16, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, M.S.; Altieri, A.H.; Angelini, C.; Bishop, M.J.; Gribben, P.E.; Lear, G.; He, Q.; Schiel, D.R.; Silliman, B.R.; South, P.M.; et al. Secondary foundation species enhance biodiversity. Nat. Ecol. Evol. 2018, 2, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Reynolds, P.L.; Boström, C.; Coyer, J.A.; Cusson, M.; Donadi, S.; Douglass, J.G.; Eklöf, J.S.; Engelen, A.H.; Eriksson, B.K. Biodiversity mediates top–down control in eelgrass ecosystems: A global comparative-experimental approach. Ecol. Lett. 2015, 18, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.E.; Richardson, J.P.; Canuel, E.A. Grazer diversity effects on ecosystem functioning in seagrass beds. Ecol. Lett. 2003, 6, 637–645. [Google Scholar] [CrossRef]
- Peterson, B.J.; Heck, K.L., Jr. An experimental test of the mechanism by which suspension feeding bivalves elevate seagrass productivity. Mar. Ecol. Prog. Ser. 2001, 218, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Van der Heide, T.; Govers, L.L.; de Fouw, J.; Olff, H.; van der Geest, M.; van Katwijk, M.M.; Piersma, T.; van de Koppel, J.; Silliman, B.R.; Smolders, A.J. A three-stage symbiosis forms the foundation of seagrass ecosystems. Science 2012, 336, 1432–1434. [Google Scholar] [CrossRef] [PubMed]
- Castorani, M.C.; Glud, R.N.; Hasler-Sheetal, H.; Holmer, M. Light indirectly mediates bivalve habitat modification and impacts on seagrass. J. Exp. Mar. Biol. Ecol. 2015, 472, 41–53. [Google Scholar] [CrossRef]
- Vinther, H.F.; Laursen, J.S.; Holmer, M. Negative effects of blue mussel (Mytilus edulis) presence in eelgrass (Zostera marina) beds in Flensborg fjord, Denmark. Estuar. Coast. Shelf Sci. 2008, 77, 91–103. [Google Scholar] [CrossRef]
- Reusch, T.B.H.; Chapman, A.R.O.; Groger, J.P. Blue mussels Mytilus edulis do not interfere with eelgrass Zostera marina but fertilize shoot growth through biodeposition. Mar. Ecol. Prog. Ser. 1994, 108, 265–282. [Google Scholar] [CrossRef]
- Wall, C.C.; Peterson, B.J.; Gobler, C.J. Facilitation of seagrass Zostera marina productivity by suspension-feeding bivalves. Mar. Ecol. Prog. Ser. 2008, 357, 165–174. [Google Scholar] [CrossRef]
- Gribben, P.E.; Kimbro, D.L.; Vergés, A.; Gouhier, T.C.; Burrell, S.; Garthwin, R.G.; Cagigas, M.L.; Tordoff, Y.; Poore, A.G.B. Positive and negative interactions control a facilitation cascade. Ecosphere 2017, 8, e02065. [Google Scholar] [CrossRef] [Green Version]
- Bologna, P.A.X.; Heck, K.L. Impacts of seagrass habitat architecture on bivalve settlement. Estuaries 2000, 23, 449–457. [Google Scholar] [CrossRef]
- Williams, S.L.; Heck, K.L. Seagrass Community Ecology. In Marine Community Ecology; Bertness, M.D., Gaines, S.D., Hay, M.E., Eds.; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 317–331. [Google Scholar]
- Eckman, J.E. The role of hydrodynamics in recruitment, growth, and survival of Agropecten irradians (L.) and Anomia simplex (D’Orbigny) within eelgrass meadows. J. Exp. Mar. Biol. Ecol. 1987, 106, 165–191. [Google Scholar] [CrossRef]
- Peterson, B.J.; Heck, K.L., Jr. Positive interactions between suspension-feeding bivalves and seagrass-a facultative mutualism. Mar. Ecol. Prog. Ser. 2001, 213, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Goff, J.; Moody, R.M.; Byron, D.; Heck, K.L.; Powers, S.P.; Ferraro, C.; Cebrian, J. Do restored oyster reefs benefit seagrasses? An experimental study in the Northern Gulf of Mexico. Restor. Ecol. 2016, 24, 306–313. [Google Scholar] [CrossRef]
- Gascoigne, J.C.; Beadman, H.A.; Saurel, C.; Kaiser, M.J. Density dependence, spatial scale and patterning in sessile biota. Oecologia 2005, 145, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Bertness, M.D. Ribbed mussels and Spartina alterniflora production in a new england salt marsh. Ecology 1984, 65, 1794–1807. [Google Scholar] [CrossRef]
- Basurto, X. Commercial diving and the callo de hacha fishery in Seri territory. J. Southwest 2006, 48, 189–209. [Google Scholar]
- Ahumada-Sempoal, M.A.; Serrano-Guzmán, S.J.; Ruiz-García, N. Abundancia, estructura poblacional y crecimiento de Atrina maura (Bivalvia: Pinnidae) en una laguna costera tropical del Pacífico mexicano. Rev. Biol. Trop. 2002, 50, 1091–1100. [Google Scholar] [PubMed]
- García-March, J.R.; García-Carrascosa, A.M.; Peña Cantero, A.L.; Wang, Y.G. Population structure, mortality and growth of Pinna nobilis Linnaeus, 1758 (Mollusca, Bivalvia) at different depths in Moraira bay (Alicante, Western Mediterranean). Mar. Biol. 2006, 150, 861. [Google Scholar] [CrossRef]
- Keough, M.J. Effects of patch size on the abundance of sessile marine invertebrates. Ecology 1984, 65, 423–437. [Google Scholar] [CrossRef]
- Van Hulzen, J.B.; van Soelen, J.; Bouma, T.J. Morphological variation and habitat modification are strongly correlated for the autogenic ecosystem engineer Spartina anglica (common cordgrass). Estuar. Coasts 2007, 30, 3–11. [Google Scholar] [CrossRef]
- Irving, A.D.; Bertness, M.D. Trait-dependent modification of facilitation on cobble beaches. Ecology 2009, 90, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.D.G.; O’Riley, J.L. Non-linear density-dependent effects of an intertidal ecosystem engineer. Oecologia 2011, 166, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.I.; Pergent, G.; Fazi, S. Chapter 16—Seagrass decomposition. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 313–324. [Google Scholar]
- Edgar, G.J.; Mukai, H.; Orth, R.J. Fish, crabs, shrimps and other large mobile epibenthos: Measurement methods for their biomass and abundance in seagrass. In Glogal Seagrass Research Methods; Elsevier: Amesterdam, The Netherlands, 2001; Volume 33, pp. 255–270. [Google Scholar]
- Kendrick, G.A.; Lavery, P.S. Assessing biomass, assemblage structure and productivity of algal epiphytes on seagrasses. In Glogal Seagrass Research Methods; Elsevier: Amesterdam, The Netherlands, 2001; pp. 199–222. [Google Scholar]
- Sidik, B.J.; Bandeira, S.O.; Milchakova, N.A. Chapter 11—Methods to measure macroalgal biomass and abundance in seagrass meadows. In Global Seagrass Research Methods; Short, F.T., Coles, R.G., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 223–235. [Google Scholar]
- Gacia, E.; Duarte, C.M. Sediment retention by a Mediterranean Posidonia oceanica meadow: The balance between deposition and resuspension. Estuar. Coast. Shelf Sci. 2001, 52, 505–514. [Google Scholar] [CrossRef]
- Ward, L.G.; Kemp, W.M.; Boynton, W.R. The influence of waves and seagrass communities on suspended particulates in an estuarine embayment. Mar. Geol. 1984, 59, 85–103. [Google Scholar] [CrossRef]
- Fonseca, M.S.; Fisher, J.S.; Zieman, J.C.; Thayer, G.W. Influence of the seagrass, Zostera marina L. on current flow. Estuar. Coast. Shelf Sci. 1982, 15, 351–364. [Google Scholar] [CrossRef]
- Peterson, C.H.; Summerson, H.C.; Duncan, P.B. The influence of seagrass cover on population structure and individual growth of a suspension-feeding bivalve, Mercenaria mercenaria. J. Mar. Res. 1984, 42, 123–138. [Google Scholar] [CrossRef]
- Heck, K.L.; Orth, R.J. Predation in seagrass beds. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R.J., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 537–550. [Google Scholar]
- Orth, R.J.; Heck, K.L.; van Montfrans, J. Faunal communities in seagrass beds: A review of the influence of plant structure and prey characteristics on predator-prey relationships. Estuaries 1984, 7, 339–350. [Google Scholar] [CrossRef]
- Keough, M.J. Dynamics of the epifauna of the bivalve Pinna bicolor: Interactions among recruitment, predation, and competition. Ecology 1984, 65, 677–688. [Google Scholar] [CrossRef]
- Hauxwell, J.; Cebrián, J.; Furlong, C.; Valiela, I. Macroalgal canopies contribute to eelgrass (Zostera marina) decline in temperate estuarine ecosystems. Ecology 2001, 82, 1007–1022. [Google Scholar] [CrossRef]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. USA 2004, 101, 8998–9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, P.L.; Richardson, J.P.; Duffy, J.E. Field experimental evidence that grazers mediate transition between microalgal and seagrass dominance. Limnol. Oceanogr. 2014, 59, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- Cardinale, B.J.; Palmer, M.A.; Collins, S.L. Species diversity enhances ecosystem functioning through interspecific facilitation. Nature 2002, 415, 426. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, C.C.; Hoellein, T.J. Bivalve Impacts in Freshwater and Marine Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 183–208. [Google Scholar] [CrossRef]
- Dame, R.F. Ecology of Marine Bivalves: An Ecosystem Approach; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Bahr, L.M.; Lanier, W.P. The Ecology of Intertidal Oyster Reefs of the South Atlantic Coast: A Community Profile; 81/15; US Fish and Wildlife Service: Washington, DC, USA, 1981.
- Lenihan, H.S.; Peterson, C.H.; Byers, J.E.; Grabowski, J.H.; Thayer, G.W.; Colby, D.R. Cascading of habitat degradation: Oyster reefs invaded by refugee fishes escaping stress. Ecol. Appl. 2001, 11, 764–782. [Google Scholar] [CrossRef]
- Wells, H.W. The fauna of oyster beds, with special reference to the salinity factor. Ecol. Monogr. 1961, 31, 239–266. [Google Scholar] [CrossRef]
- Gedan, K.B.; Kellogg, L.; Breitburg, D.L. Accounting for multiple foundation species in oyster reef restoration benefits. Restor. Ecol. 2014, 22, 517–524. [Google Scholar] [CrossRef]
- Grabowski, J.H.; Peterson, C.H. Restoring oyster reefs to recover ecosystem services. Theor. Ecol. Ser. 2007, 4, 281–298. [Google Scholar]
- Gutiérrez, J.L.; Bagur, M.; Palomo, M.G. Algal Epibionts as Co-Engineers in Mussel Beds: Effects on Abiotic Conditions and Mobile Interstitial Invertebrates. Diversity 2019, 11, 17. [Google Scholar] [CrossRef]
- Menge, B.A. Organization of the new england rocky intertidal community: Role of predation, competition, and environmental heterogeneity. Ecol. Monogr. 1976, 46, 355–393. [Google Scholar] [CrossRef]
- Levinton, J. Bioturbators as ecosystem engineers: Control of the sediment fabric, inter-individual interactions, and material fluxes. In Linking Species & Ecosystems; Springer: Boston, MA, USA, 1995; pp. 29–36. [Google Scholar]
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshwat. Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Rosenberg, R. Ecosystem engineering: The impact of bioturbation on biogeochemical processes in marine and freshwater benthic habitats. Aquat. Sci. 2006, 68, 434–442. [Google Scholar] [CrossRef]
- Kristensen, E.; Penha-Lopes, G.; Delefosse, M.; Valdemarsen, T.; Quintana, C.O.; Banta, G.T. What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 2012, 446, 285–302. [Google Scholar] [CrossRef] [Green Version]
- Escamilla-Montes, R.; Diarte-Plata, G.; Luna, A.; Arturo Fierro-Coronado, J.; Esparza-Leal, H.; Granados-Alcantar, S.; Ruiz-Verdugo, C.A. Ecology, Fishery and Aquaculture in Gulf of California, Mexico: Pen Shell Atrina Maura (Sowerby, 1835); IntechOpen: London, UK, 2017. [Google Scholar]
- Beckett, D.C.; Green, B.W.; Thomas, S.A.; Miller, A.C. Epizoic invertebrate communities on upper Mississippi River unionid bivalves. Am. Midl. Nat. 1996, 135, 102–114. [Google Scholar] [CrossRef]
- Gribben, P.E.; Byers, J.E.; Clements, M.; McKenzie, L.A.; Steinberg, P.D.; Wright, J.T. Behavioural interactions between ecosystem engineers control community species richness. Ecol. Lett. 2009, 12, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmann, M.L. Spatial and temporal patterns in the dynamics and use of pen shells (Atrina rigida) as shelters in St. Joseph Bay, Florida. Bull. Mar. Sci. 1998, 62, 157–179. [Google Scholar]
- Gutiérrez, J.; Iribarne, O. Role of Holocene beds of the stout razor clam Tagelus plebeius in structuring present benthic communities. Mar. Ecol. Prog. Ser. 1999, 185, 213–228. [Google Scholar] [CrossRef] [Green Version]
- Finke, D.L.; Denno, R.F. Predator diversity dampens trophic cascades. Nature 2004, 429, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.S.; Cioffi, W.R.; Cope, R.; Daleo, P.; Heywood, E.; Hoyt, C.; Smith, C.S.; Silliman, B.R. A global synthesis reveals gaps in coastal habitat restoration research. Sustainability 2018, 10, 1040. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.S.; Silliman, B.R. A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds. Diversity 2019, 11, 30. https://doi.org/10.3390/d11030030
Zhang YS, Silliman BR. A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds. Diversity. 2019; 11(3):30. https://doi.org/10.3390/d11030030
Chicago/Turabian StyleZhang, Y. Stacy, and Brian R. Silliman. 2019. "A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds" Diversity 11, no. 3: 30. https://doi.org/10.3390/d11030030
APA StyleZhang, Y. S., & Silliman, B. R. (2019). A Facilitation Cascade Enhances Local Biodiversity in Seagrass Beds. Diversity, 11(3), 30. https://doi.org/10.3390/d11030030