Testing the Poleotolerance Lichen Response Trait as an Indicator of Anthropic Disturbance in an Urban Environment



Abstract
:1. Introduction
2. Materials and Methods
2.1. Confirming Species Classification
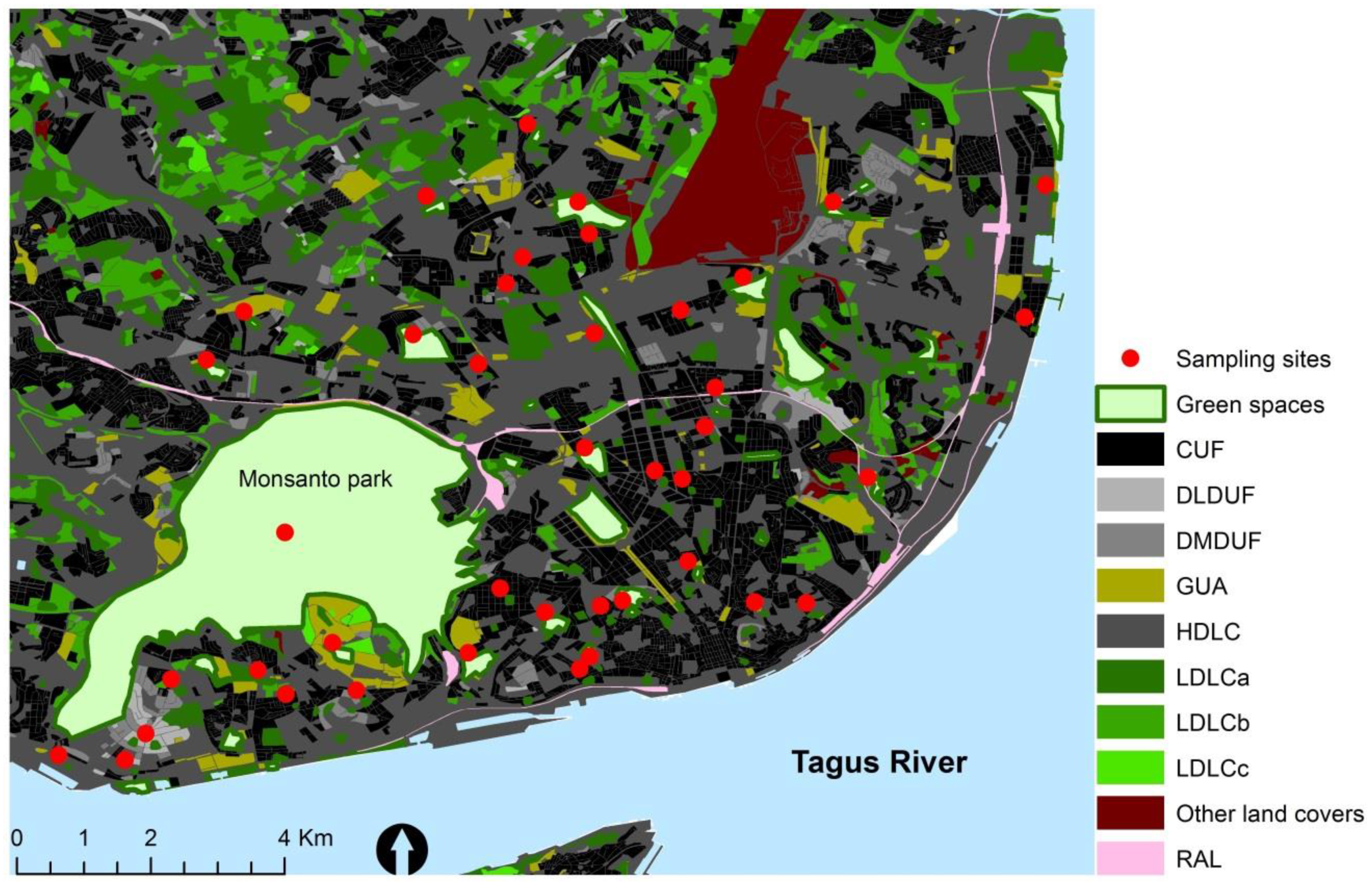
2.2. Lichen Sampling
2.3. Environmental Variables
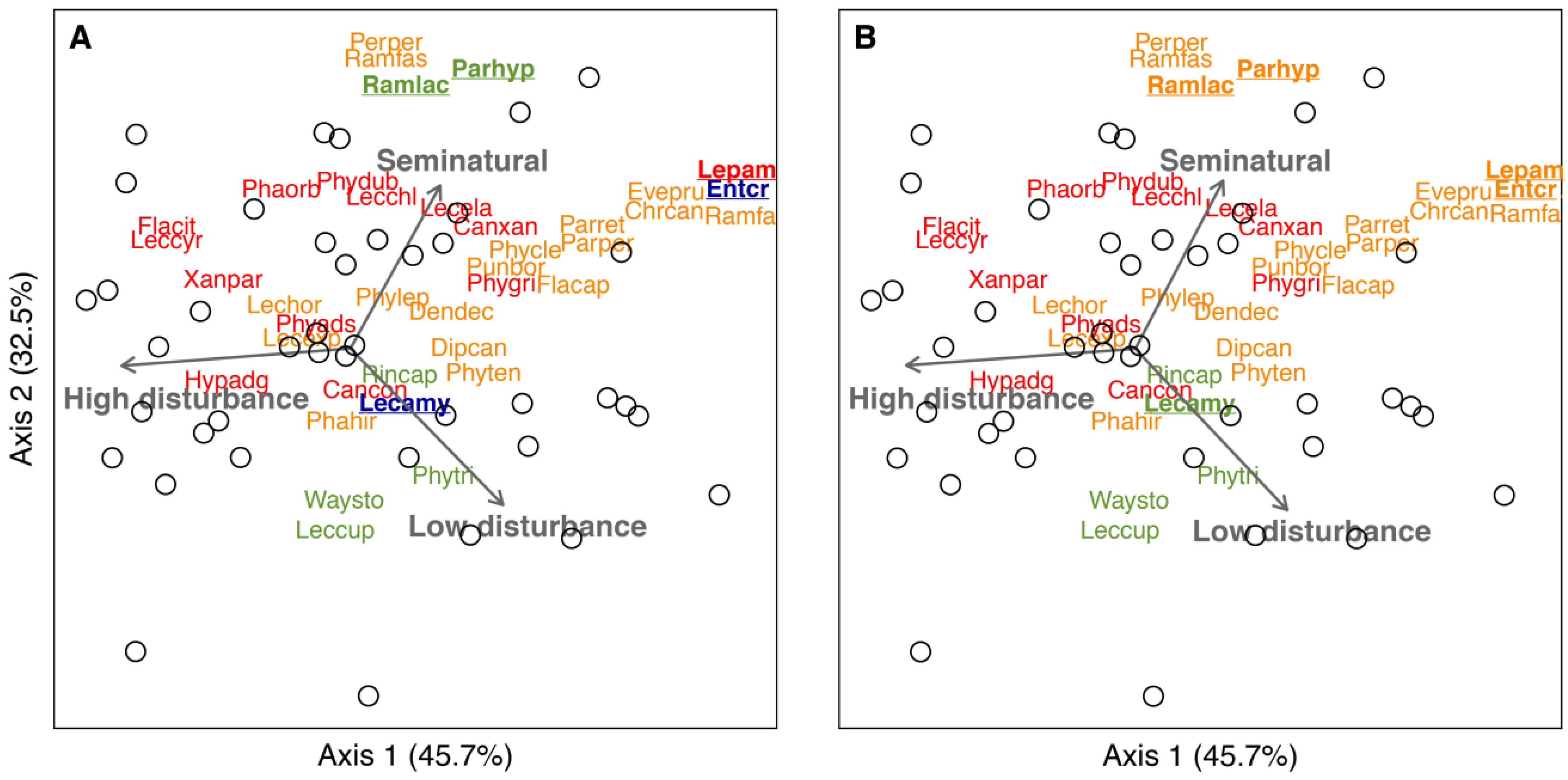
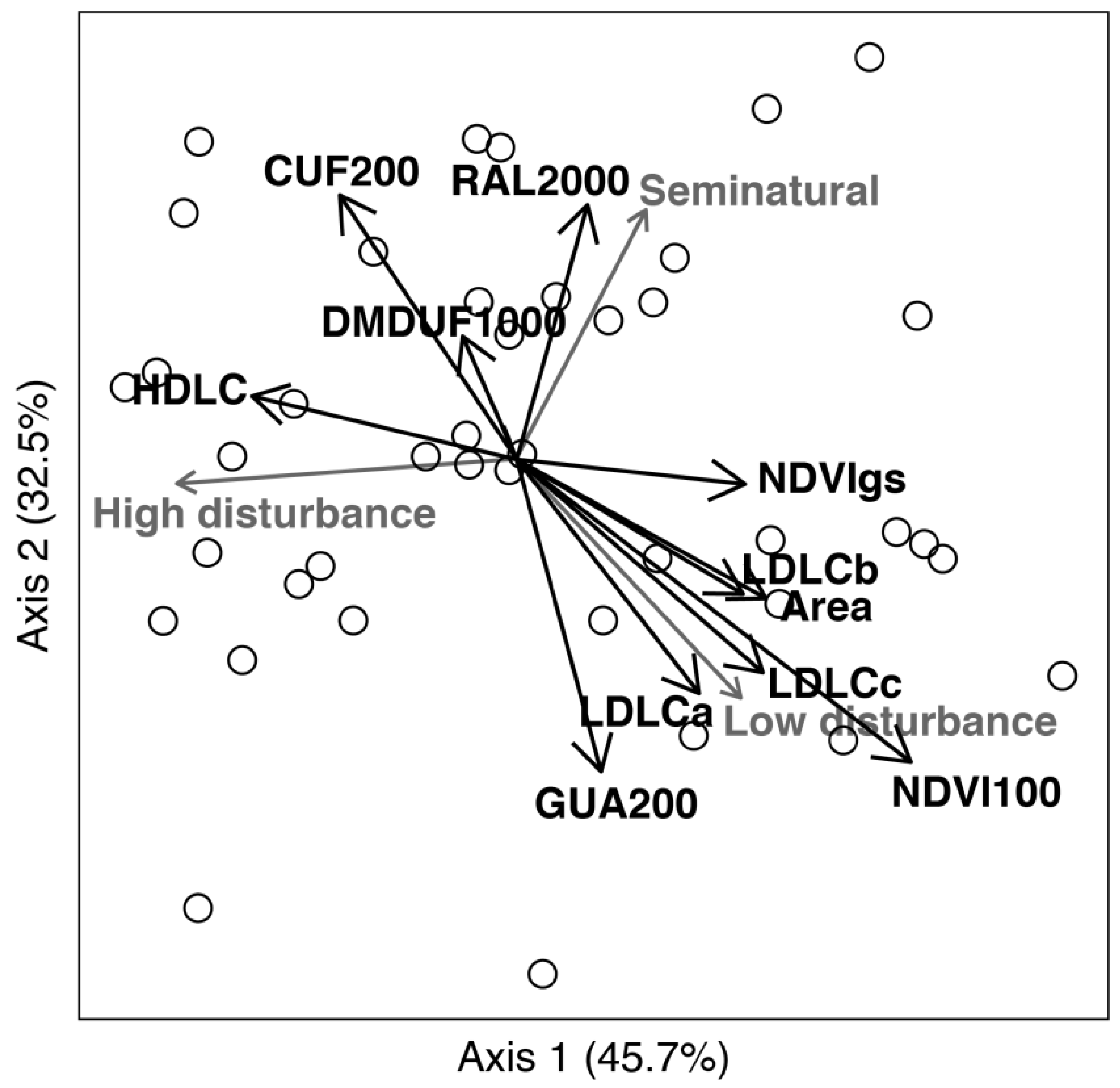
2.4. Data Analysis
3. Results
3.1. Confirming Species Classification
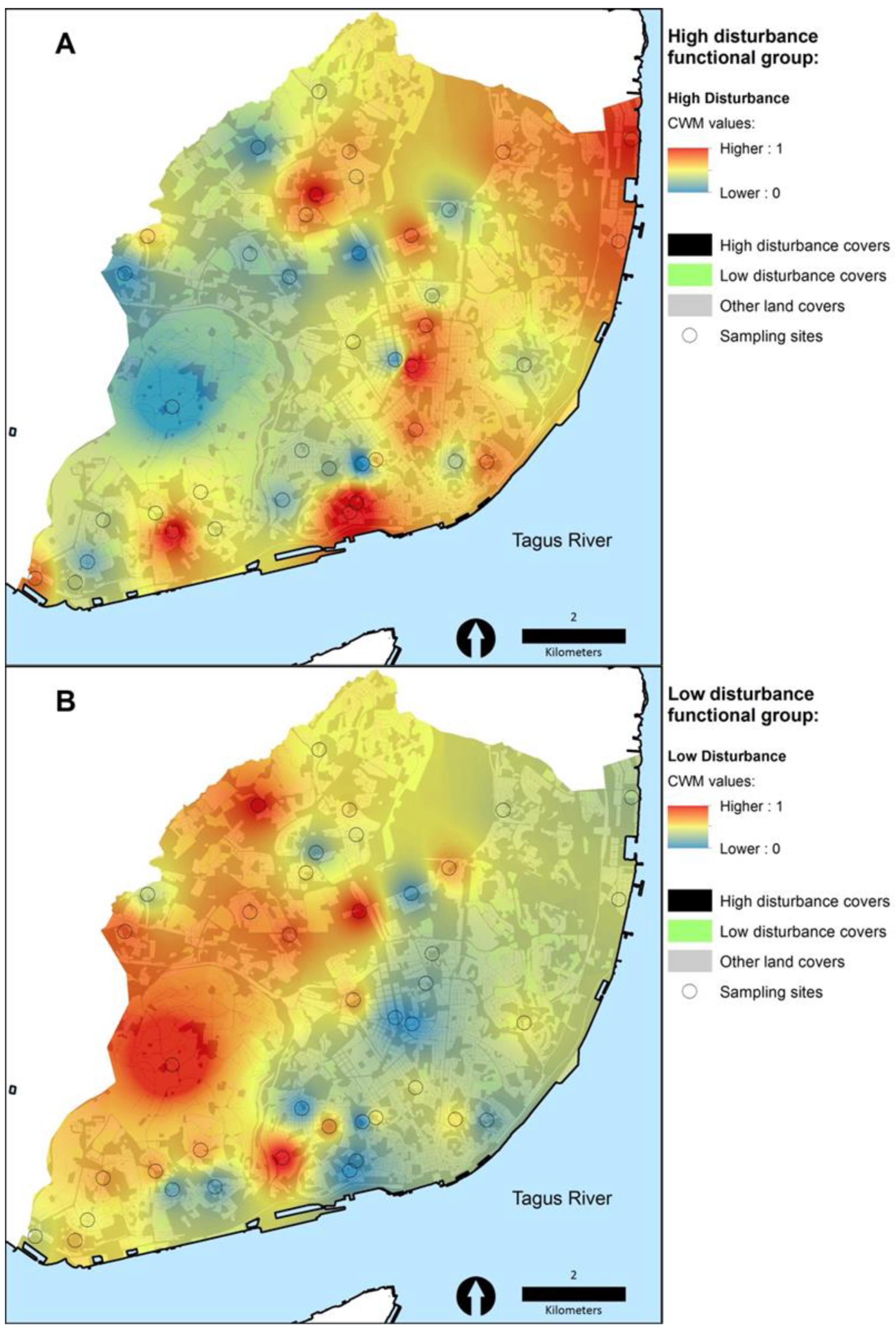
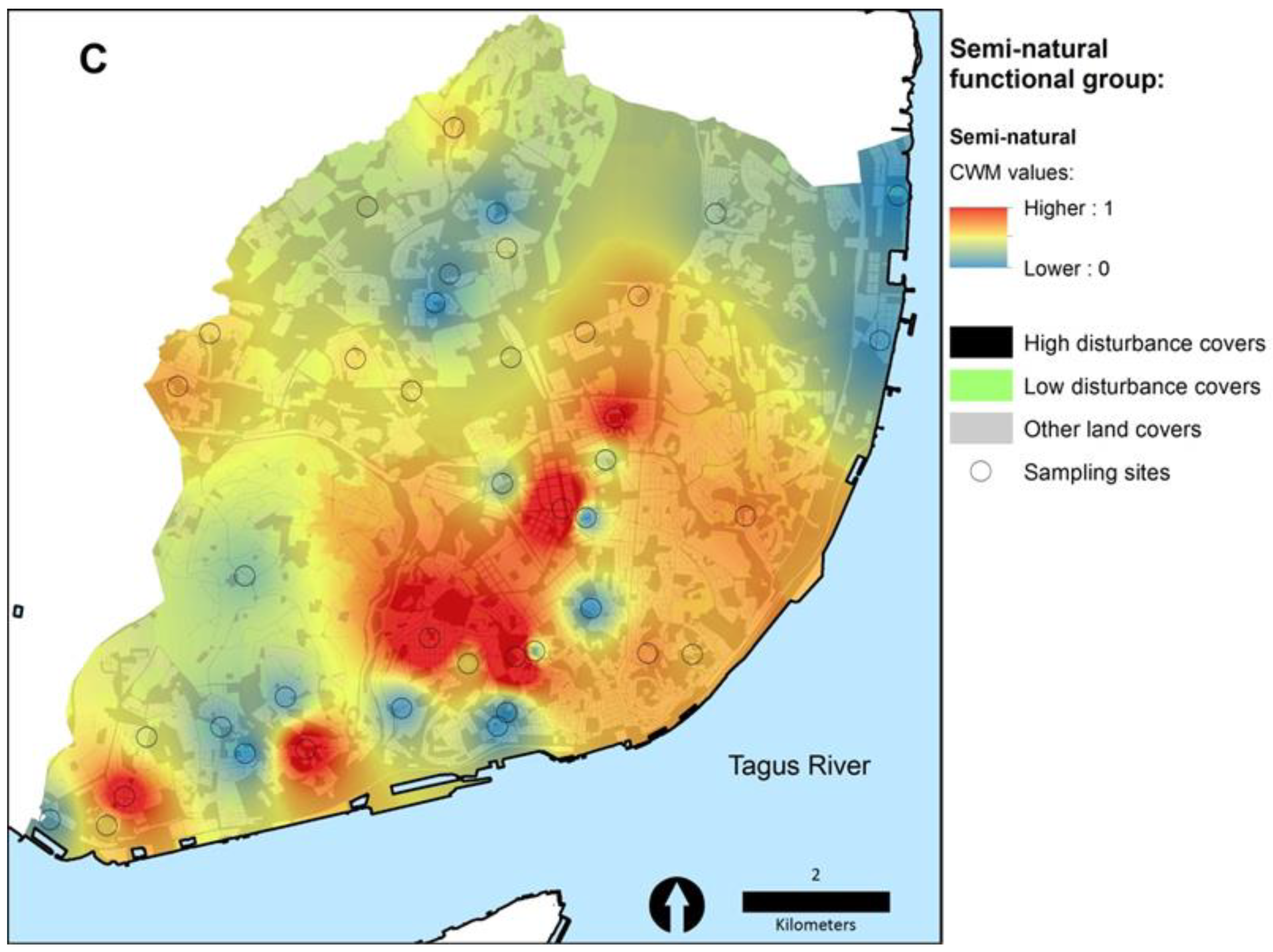
3.2. The Poleotolerance Response Trait as an Indicator of Urban Disturbance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Roser, M.; Ortiz-Ospina, E. World Population Growth. 2019. Available online: https://ourworldindata.org/world-population-growth:OurWorldInData.org (accessed on 19 December 2018).
- Lutz, W.; Sanderson, W.C.; Scherbov, S. Probabilistic World Population Projections Based on ExpertOpinion; IIASA—International Institute for Applied Systems Analysis: Laxenburg, Austria, 1996. [Google Scholar]
- Agency, E.E. Increasing Environmental Pollution (GMT 10); European Union: Brussels, Belgium, 2015; pp. 1–5. [Google Scholar]
- Rosenzweig, C.; Iglesias, A.; Yang, X.; Epstein, P.R.; Chivian, E. Climate change and extreme weather events; implications for food production, plant diseases, and pests. Glob. Chang. Hum. Health 2001, 2, 90–104. [Google Scholar] [CrossRef]
- Achard, F.; Eva, H.D.; Stibig, H.-J.; Mayaux, P.; Gallego, J.; Richards, T.; Malingreau, J.-P. Determination of deforestation rates of the world’s humid tropical forests. Science 2002, 297, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- DeFries, R.S.; Rudel, T.; Uriarte, M.; Hansen, M. Deforestation driven by urban population growth and agricultural trade in the twenty-first century. Nat. Geosci. 2010, 3, 178. [Google Scholar] [CrossRef]
- Boyd, B. Urbanization and the Mass Movement of People to Cities. Available online: https://graylinegroup.com/urbanization-catalyst-overview/ (accessed on 28 December 2018).
- United Nations, Department of Economic and Social Affairs, Population Division. World Urbanization Prospects: The 2014 Revision-Highlights; UN (ST/ESA/SER.A/352): New York, NY, USA, 2014. [Google Scholar]
- Niemi, G.J.; McDonald, M.E. Application of ecological indicators. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 89–111. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Tongway, D.J.; Bastin, G.N.; James, C.D. Monitoring ecological indicators of rangeland functional integrity and their relation to biodiversity at local to regional scales. Austral Ecol. 2004, 29, 108–120. [Google Scholar] [CrossRef]
- Marinari, S.; Mancinelli, R.; Campiglia, E.; Grego, S. Chemical and biological indicators of soil quality in organic and conventional farming systems in Central Italy. Ecol. Indic. 2006, 6, 701–711. [Google Scholar] [CrossRef]
- McCune, B. Lichen communities as indicators of forest health. Bryologist 2000, 103, 353–356. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Arup, U.; Baranowski, R.; Ekman, S. Tree-dependent lichens and beetles as indicators in conservation forests. Conserv. Biol. 1995, 9, 1208–1215. [Google Scholar] [CrossRef]
- Matos, P.; Vieira, J.; Rocha, B.; Branquinho, C.; Pinho, P. Modeling the provision of air-quality regulation ecosystem service provided by urban green spaces using lichens as ecological indicators. Sci. Total Environ. 2019, 665, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Matos, P.; Mexia, T.; Silva, P.; Lopes, N.; Freitas, C.; Correia, O.; Santos-Reis, M.; Branquinho, C.; Pinho, P. Green spaces are not all the same for the provision of air purification and climate regulation services: The case of urban parks. Environ. Res. 2018, 160, 306–313. [Google Scholar] [CrossRef]
- Matos, P.; Pinho, P.; Aragon, G.; Martínez, I.; Nunes, A.; Soares, A.M.; Branquinho, C. Lichen traits responding to aridity. J. Ecol. 2015, 103, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Aptroot, A.; Van Herk, C. Further evidence of the effects of global warming on lichens, particularly those with Trentepohlia phycobionts. Environ. Pollut. 2007, 146, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, Y.; Seaward, M.R. Is Glyphis cicatricosa an indicator for ‘global warming’or an ‘urban heat island’effect in Japan? Lichenologist 2017, 49, 291–296. [Google Scholar] [CrossRef]
- Asta, J.; Erhardt, W.; Ferretti, M.; Fornasier, F.; Kirschbaum, U.; Nimis, P.; Purvis, O.; Pirintsos, S.; Scheidegger, C.; Van Haluwyn, C. Mapping lichen diversity as an indicator of environmental quality. In Monitoring with Lichens—Monitoring Lichens; Springer: Berlin/Heidelberg, Germany, 2002; pp. 273–279. [Google Scholar]
- Smith, W.B. Forest inventory and analysis: A national inventory and monitoring program. Environ. Pollut. 2002, 116, S233–S242. [Google Scholar] [CrossRef]
- Llop, E.; Pinho, P.; Matos, P.; Pereira, M.J.; Branquinho, C. The use of lichen functional groups as indicators of air quality in a Mediterranean urban environment. Ecol. Indic. 2012, 13, 215–221. [Google Scholar] [CrossRef]
- Saiki, M.; Fuga, A.; Alves, E.; Vasconcellos, M.; Marcelli, M. Biomonitoring of the atmospheric pollution using lichens in the metropolitan area of São Paulo city, Brazil. J. Radioanal. Nucl. Chem. 2006, 271, 213–219. [Google Scholar] [CrossRef]
- Munzi, S.; Ravera, S.; Caneva, G. Epiphytic lichens as indicators of environmental quality in Rome. Environ. Pollut. 2007, 146, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Munzi, S.; Correia, O.; Silva, P.; Lopes, N.; Freitas, C.; Branquinho, C.; Pinho, P. Lichens as ecological indicators in urban areas: Beyond the effects of pollutants. J. Appl. Ecol. 2014, 51, 1750–1757. [Google Scholar] [CrossRef]
- Branquinho, C.; Serrano, H.C.; Nunes, A.; Pinho, P.; Matos, P. From Assessing to Conserving Biodiversity. In History, Philosophy and Theory of the Life Sciences, 1st ed.; Casetta, E., Silva, J.M.D., Vecchi, D., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; p. 290. [Google Scholar]
- Dıaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Mouillot, D.; Villéger, S.; Parravicini, V.; Kulbicki, M.; Arias-González, J.E.; Bender, M.; Chabanet, P.; Floeter, S.R.; Friedlander, A.; Vigliola, L. Functional over-redundancy and high functional vulnerability in global fish faunas on tropical reefs. Proc. Natl. Acad. Sci. USA 2014, 111, 13757–13762. [Google Scholar] [CrossRef] [Green Version]
- Koch, N.M.; Matos, P.; Branquinho, C.; Pinho, P.; Lucheta, F.; de Azevedo Martins, S.M.; Vargas, V.M.F. Selecting lichen functional traits as ecological indicators of the effects of urban environment. Sci. Total Environ. 2019, 654, 705–713. [Google Scholar] [CrossRef]
- Varela, Z.; López-Sánchez, G.; Yáñez, M.; Pérez, C.; Fernández, J.; Matos, P.; Branquinho, C.; Aboal, J. Changes in epiphytic lichen diversity are associated with air particulate matter levels: The case study of urban areas in Chile. Ecol. Indic. 2018, 91, 307–314. [Google Scholar] [CrossRef]
- Barkman, J.J. Phytosociology and Ecology of Cryptogamic Epiphytes; Van Gorcum, University of Michigan: Ann Arbor, MI, USA, 1958. [Google Scholar]
- DeSLOOVER, J. Mapping on atmospheric pollution on the basis of lichen sensitivity. Proc. Symposium Recent Adv. Trop. Ecol. 1968, 1, 42–56. [Google Scholar]
- Trass, H. Lichen sensitivity to the air pollution and index of poleotolerance (IP). Folia Cryptog Est. 1973, 3, 19–22. [Google Scholar]
- Nimis, P.L. The Epiphytic Lichen Vegetation of the Trieste Province (North Eastern Italy); EUT Edizioni Università di Trieste: Triste, Italy, 1982. [Google Scholar]
- Nimis, P. Urban Lichen Studies in Italy. II: The town of Udine. Gortania 1986, 7, 147–172. [Google Scholar]
- Nimis, P. Urban lichen studies in Italy. III: The city of Rome. Braun-Blanquetia 1989, 2, 223–238. [Google Scholar]
- Nimis, P.L. The Lichens of Italy. A Second Annotated Catalogue; EUT Edizioni Università di Trieste: Triste, Italy, 2016. [Google Scholar]
- Degtjarenko, P.; Matos, P.; Marmor, L.; Branquinho, C.; Randlane, T. Functional traits of epiphytic lichens respond to alkaline dust pollution. Fungal Ecol. 2018, 36, 81–88. [Google Scholar] [CrossRef]
- Nimis, P.L.; Martellos, S.; Spitale, D.; Nascimbene, J. Exploring patterns of commonness and rarity in lichens: A case study from Italy (Southern Europe). Lichenologist 2018, 50, 385–396. [Google Scholar] [CrossRef]
- Gilbert, O. Further studies on the effect of sulphur dioxide on lichens and bryophytes. New Phytol. 1970, 69, 605–627. [Google Scholar] [CrossRef]
- Branquinho, C.; Matos, P.; Pinho, P. Lichens as ecological indicators to track atmospheric changes: Future challenges. In Indicators and Surrogates of Biodiversity and Environmental Change; CSIRO Publishing: Melbourne, Australian; CRC Press: London, UK, 2015; pp. 77–87. [Google Scholar]
- Nowak, D.J.; Jovan, S.; Branquinho, C.; Augusto, S.; Ribeiro, M.C.; Kretsch, C.E. Biodiversity, air quality and human health. In Connecting Global Priorities: Biodiversity and Human Health: A State of Knowledge Review; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- INE. Censos 2011 Resultados Definitivos-Portugal; Instituto Nacional de Estatística, IP: Lisboa, Portugal, 2012. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- PORDATA. Available online: https://www.pordata.pt/Portugal (accessed on 15 January 2019).
- Económico, O.J. Todos os dias entram 370 mil carros em Lisboa. O Jornal Económico. Available online: https://jornaleconomico.sapo.pt/noticias/os-dias-entram-lisboa-370-mil-carros-84046 (accessed on 6 April 2019).
- EEA. Copernicus Land Monitoring Service—Urban Atlas. 2018. Available online: www.eea.europa.eu/data-and-maps/data/copernicus-land-monitoring-service-urban-atlas (accessed on 10 January 2015).
- Martellos, S. Multi-authored interactive identification keys: The FRIDA (FRiendly IDentificAtion) package. Taxon 2010, 59, 922–929. [Google Scholar] [CrossRef]
- Smith, C.W.; Aptroot, A.; Coppins, B.J.; Fletcher, A.; Gilbert, O.L.; James, P.W.; Wolseley, P.A. The Lichens of Great Britain and Ireland; British Lichen Society: London, UK, 2009. [Google Scholar]
- Lavorel, S.; Grigulis, K.; McIntyre, S.; Williams, N.S.; Garden, D.; Dorrough, J.; Berman, S.; Quétier, F.; Thébault, A.; Bonis, A. Assessing functional diversity in the field–methodology matters! Funct. Ecol. 2008, 22, 134–147. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology, R package version 1.0-12; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002; Volume 28. [Google Scholar]
- McCune, B.; Mefford, M. PC-ORD: Multivariate Analysis of Ecological Data; Version 4 for Windows; [User’s Guide]; MjM Software Design: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Pinho, P.; Augusto, S.; Maguas, C.; Pereira, M.; Soares, A.; Branquinho, C. Impact of neighbourhood land-cover in epiphytic lichen diversity: Analysis of multiple factors working at different spatial scales. Environ. Pollut. 2008, 151, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Pinho, P.; Dias, T.; Cruz, C.; Sim Tang, Y.; Sutton, M.A.; Martins-Loução, M.A.; Máguas, C.; Branquinho, C. Using lichen functional diversity to assess the effects of atmospheric ammonia in Mediterranean woodlands. J. Appl. Ecol. 2011, 48, 1107–1116. [Google Scholar] [CrossRef] [Green Version]
- Root, H.T.; Geiser, L.H.; Jovan, S.; Neitlich, P. Epiphytic macrolichen indication of air quality and climate in interior forested mountains of the Pacific Northwest, USA. Ecol. Indic. 2015, 53, 95–105. [Google Scholar] [CrossRef]
- Valencia-Islas, N.; Zambrano, A.; Rojas, J. Ozone reactivity and free radical scavenging behavior of phenolic secondary metabolites in lichens exposed to chronic oxidant air pollution from Mexico City. J. Chem. Ecol. 2007, 33, 1619–1634. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Group | Description |
|---|---|
| Natural | Species which exclusively occur on old trees in ancient, undisturbed forests. |
| Seminatural | Species occurring in natural or seminatural habitats |
| Low disturbance | Species occurring in moderately disturbed areas |
| High disturbance | Species occurring in heavily disturbed areas |
| Area | NDVIgs | NDVIb100 | CUF200 | DLDUF100 | DMDUF1000 | GUA200 | RAL2000 | HDLC | LDLCa | LDLCb | LDLCc | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NMS1 | 0.54 *** | 0.34 * | 0.65 *** | −0.33 * | −0.01 | 0.01 | 0.12 | 0.12 | −0.35 * | 0.34 * | 0.49 ** | 0.34 * |
| NMS2 | 0.40 * | 0.01 | 0.39 * | −0.27 | −0.01 | −0.40 * | 0.45 ** | −0.36 * | −0.13 | 0.3 | 0.15 | 0.22 |
| NMS3 | 0.14 | 0.06 | 0 | 0.06 | −0.36 * | −0.02 | −0.02 | 0.06 | 0.05 | −0.13 | −0.05 | −0.01 |
| CWM High disturbance | −0.41 ** | −0.34 * | −0.57 *** | 0.3 | −0.15 | −0.18 | −0.01 | −0.09 | 0.35 * | −0.32 * | −0.43 ** | −0.34 * |
| CWM Low disturbance | 0.60 *** | 0.21 | 0.72 *** | −0.51 *** | 0.1 | 0.01 | 0.3 | −0.17 | −0.15 | 0.48 ** | 0.43 ** | 0.41 ** |
| CWM Seminatural | 0.07 | 0.2 | 0.14 | −0.01 | 0.16 | 0.32 * | −0.17 | 0.19 | −0.19 | 0.04 | 0.34 * | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, B.; Pinho, P.; Vieira, J.; Branquinho, C.; Matos, P. Testing the Poleotolerance Lichen Response Trait as an Indicator of Anthropic Disturbance in an Urban Environment. Diversity 2019, 11, 55. https://doi.org/10.3390/d11040055
Rocha B, Pinho P, Vieira J, Branquinho C, Matos P. Testing the Poleotolerance Lichen Response Trait as an Indicator of Anthropic Disturbance in an Urban Environment. Diversity. 2019; 11(4):55. https://doi.org/10.3390/d11040055
Chicago/Turabian StyleRocha, Bernardo, Pedro Pinho, Joana Vieira, Cristina Branquinho, and Paula Matos. 2019. "Testing the Poleotolerance Lichen Response Trait as an Indicator of Anthropic Disturbance in an Urban Environment" Diversity 11, no. 4: 55. https://doi.org/10.3390/d11040055
APA StyleRocha, B., Pinho, P., Vieira, J., Branquinho, C., & Matos, P. (2019). Testing the Poleotolerance Lichen Response Trait as an Indicator of Anthropic Disturbance in an Urban Environment. Diversity, 11(4), 55. https://doi.org/10.3390/d11040055