Lichen Responses to Disturbance: Clues for Biomonitoring Land-use Effects on Riparian Andean Ecosystems


Abstract
:1. Introduction
2. Materials and Methods
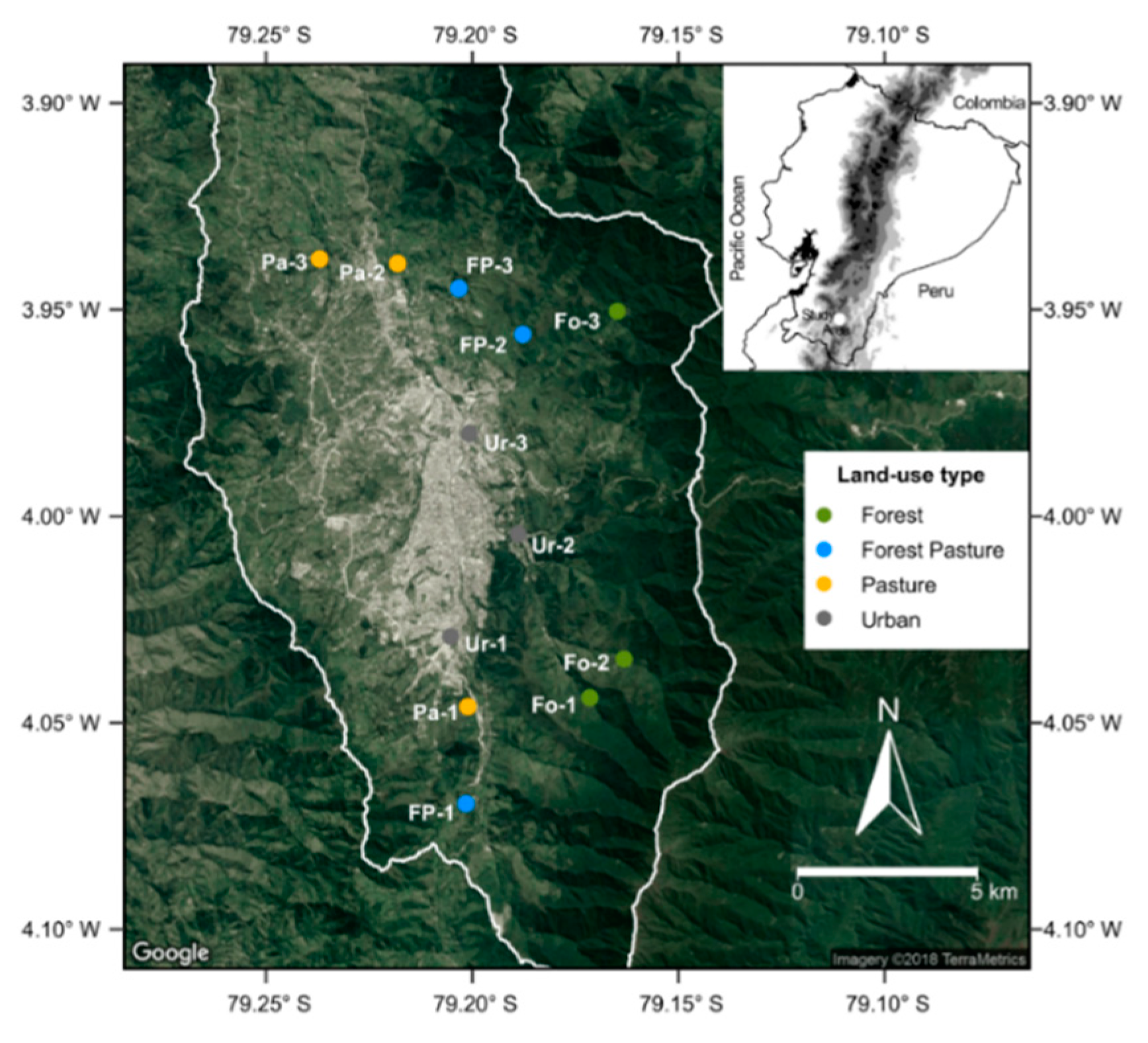
2.1. Study Area
2.2. Data Collection
2.3. Environmental Variables
2.4. Data Analysis
3. Results
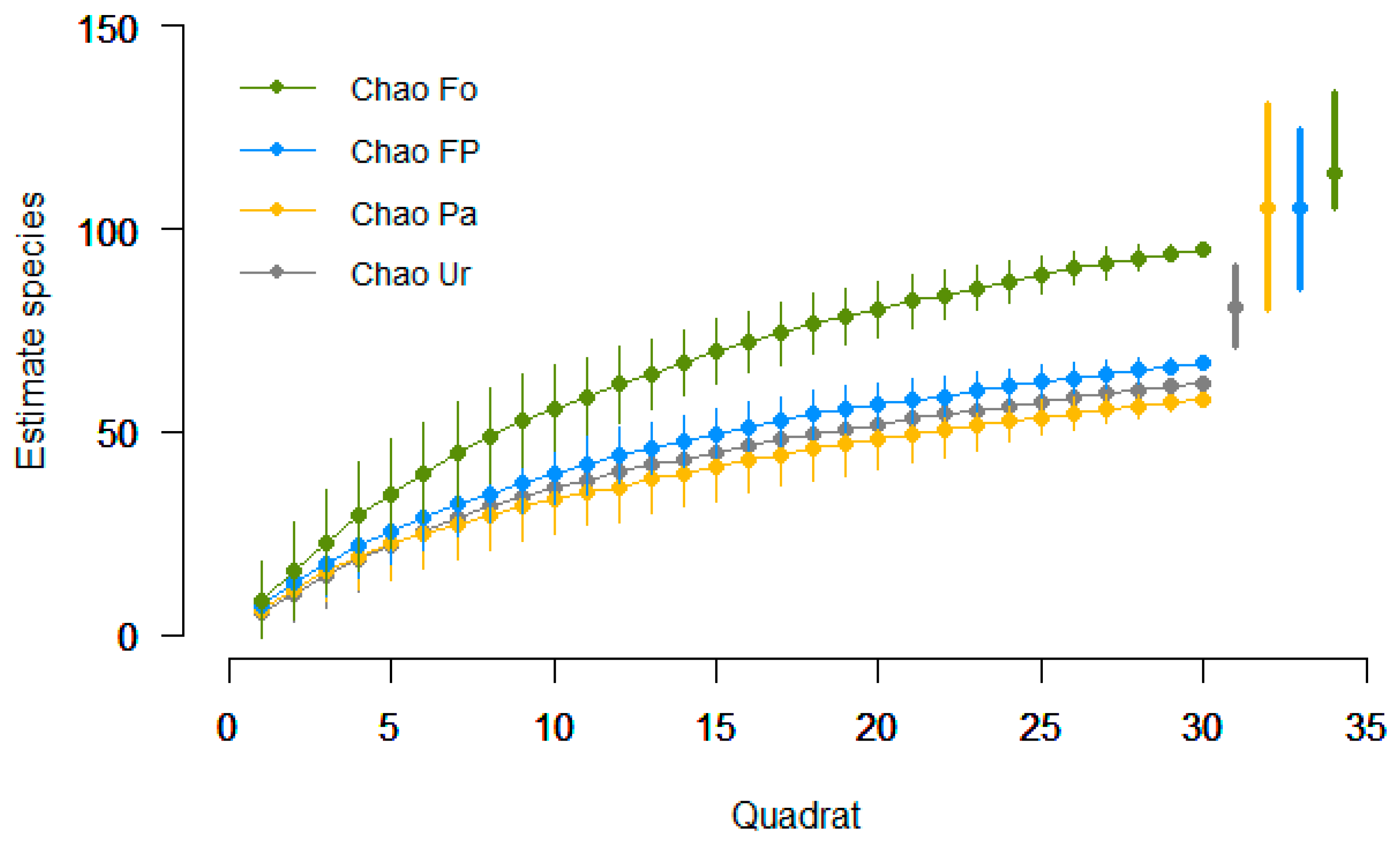
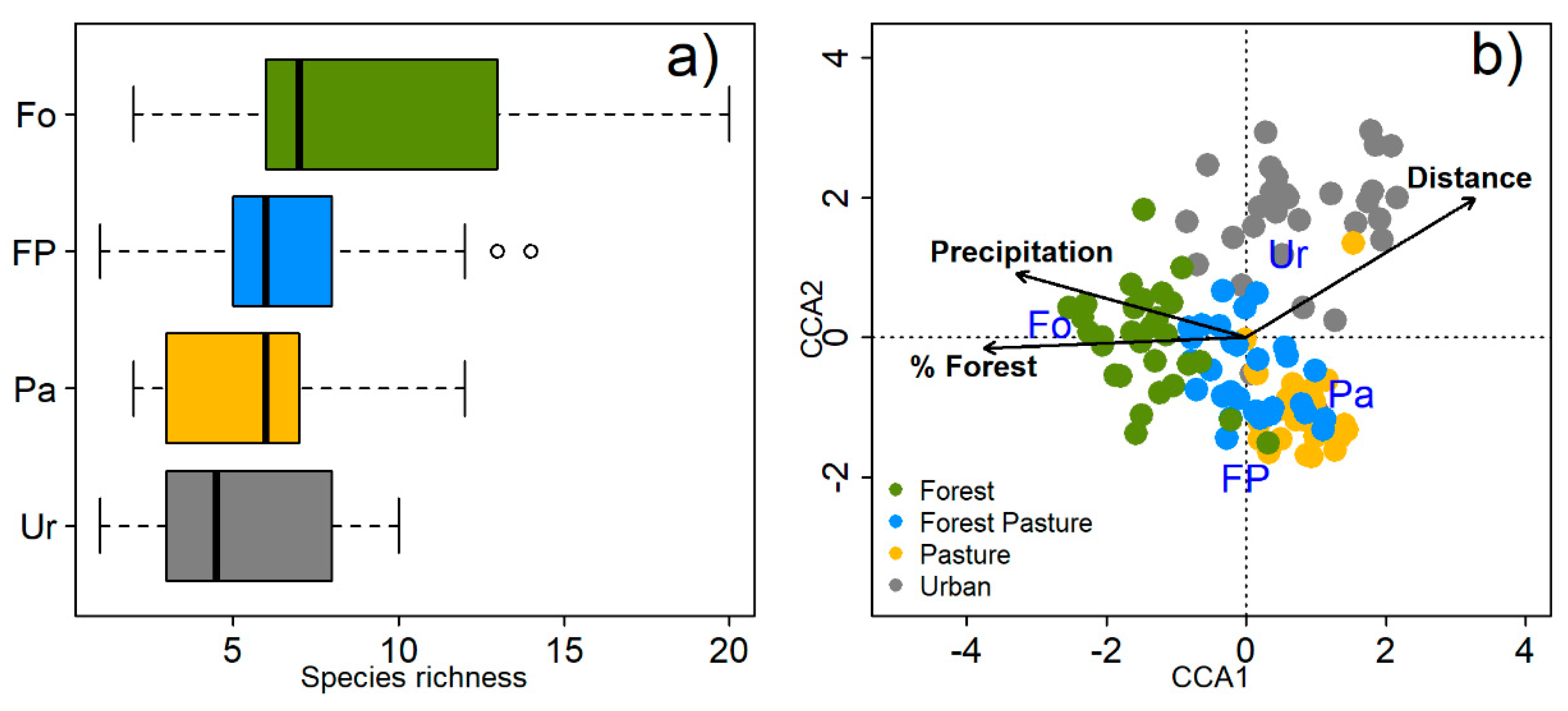
3.1. Species Richness and Community Composition
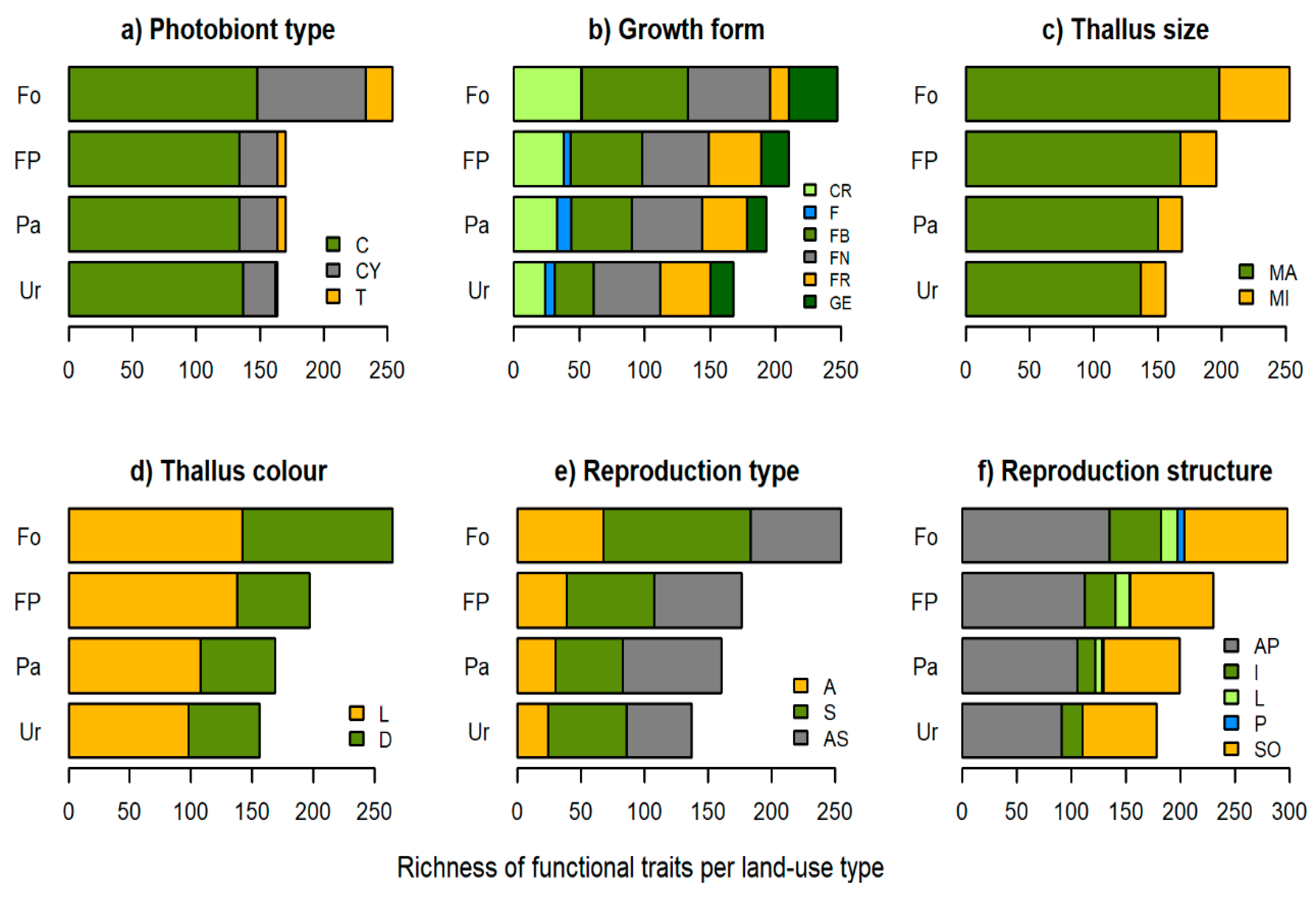
3.2. Functional Traits
4. Discussion
4.1. Species Richness, Functional Traits and Community Composition
4.2. Application in Biomonitoring
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Land-use Intensity | Indicator Value | p-value |
|---|---|---|---|
| Bacidia sp. | Forest | 3.3 | 1.000 |
| Bacidia sp. 1 | Pasture | 1.7 | 1.000 |
| Bacidia sp. 2 | Forest | 3.3 | 1.000 |
| Bacidia sp. 3 | Forest-pasture | 3.3 | 1.000 |
| Byssoloma subdiscordans (Nyl.) P. James | Pasture | 4.8 | 0,3211 |
| Candelaria concolor (Dickson) Stein | Urban | 6.7 | 0.2438 |
| Candelaria sp. | Forest | 3.6 | 0.6059 |
| Canomaculina pilosa (Stizenb.) Elix & Hale | Pasture | 6.2 | 0.2464 |
| Chapsa aff. dilatata (Müll. Arg.) Kalb | Forest | 1.9 | 1.00 |
| Chapsa aff. diploschistoides (Zahlbr.) Frisch | Forest | 3.3 | 1.000 |
| Chapsa sp. | Forest-pasture | 5.3 | 0.4695 |
| Chrysothrix candelaris (L.) JR Laundon | Urban | 10.7 | 0.0494 |
| Cladonia coniocraea (Flörke) Sprengel | Forest | 23.3 | 0.0008 |
| Coccocarpia palmicola (Spreng.) Arv. & D.J. Galloway | Forest | 3.3 | 1.000 |
| Coccocarpia pellita (Ach.) Müll. Arg. | Forest | 6.7 | 0.2368 |
| Coenogonium linkii Ehrenb | Forest | 20 | 0.0016 |
| Coenogoium pineti (Ach.) Lücking & Lumbsch | Forest-pasture | 3.3 | 1.000 |
| Dirinaria picta (Sw.) Clem. & Schear | Pasture | 10 | 0.0444 |
| Flavopunctelia flaventior (Stirt.) Hale | Pasture | 31.7 | 0.0012 |
| Graphis sp. | Forest | 6.7 | 0.2272 |
| Graphis sp. 1 | Forest | 3.3 | 1.000 |
| Graphis sp. 2 | Forest | 3.3 | 1.000 |
| Graphis sp. 3 | Forest | 3.3 | 1.000 |
| Heterodermia albicans (Pers.) Swinscow & Krog | Forest-pasture | 3.3 | 0.7986 |
| Heterodermia andina Moberg | Urban | 5.7 | 0.2745 |
| Heterodermia comosa (Eschw.) Follmann & Redón | Pasture | 3.3 | 1.000 |
| Heterodermia corallophora (Taylor) Skorepa | Pasture | 15.1 | 0.0162 |
| Heterodermia diademata (Taylor) D.D. Awasthi | Forest | 3.3 | 1.000 |
| Heterodermia galactophylla (Tuck.) W.L. Culb. | Pasture | 5.4 | 0.4147 |
| Heterodermia hypoleuca (Mühl.) Trevis. | Urban | 18 | 0.0026 |
| Heterodermia isidiophora (Nyl.) D.D. Awasthi | Forest | 14.1 | 0.0468 |
| Heterodermia japonica (M. Satô) Swinscow & Krog | Pasture | 12.2 | 0.0714 |
| Heterodermia leucomela (L.) Poelt | Urban | 15.5 | 0.0084 |
| Heterodermia pseudospeciosa (Kurok.) Culb. | Forest | 3 | 0.6067 |
| Heterodermia sitchensis Goward & Noble | Pasture | 14.8 | 0,2697 |
| Heterodermia squamulosa (Degel.) Culb | Forest-pasture | 13 | 0.0086 |
| Heterodermia spathulifera Moberg & Purvis | Forest | 21.8 | 0.0006 |
| Heterodermia speciosa (Wulfen) Trevisan | Pasture | 2 | 1.000 |
| Heterodermia sp. | Forest | 3.3 | 1.000 |
| Hypotrachyna costaricensis (Nyl.) Hale | Forest-pasture | 6.7 | 0.2446 |
| Hypotrachyna revoluta (Flörke) Hale | Forest | 4.6 | 0.2931 |
| Hypotrachyna rockii (Zahlbr.) Hale | Forest | 34 | 0.0002 |
| Hypotrachyna reducens (Nyl.) Hale | Pasture | 15.7 | 0.1236 |
| Hypotrachyna sinuosa (Sm.) Hale | Forest | 2.2 | 1.000 |
| Lecanora chlarotera Nyl. | Urban | 10 | 0.0566 |
| Lecanora helva Stizenb. | Urban | 3.3 | 1.000 |
| Lecanora sp. | Urban | 3.3 | 1.000 |
| Lepraria sp. | Forest | 3.8 | 0.7365 |
| Leptogium austroamericanum (Malme) CW Dodge | Forest | 6.3 | 0.1882 |
| Leptogium azureum (Sw.) Mont. | Urban | 7.4 | 0.2442 |
| Leptogium burgesii (L.) Mont. | Forest | 20 | 0,0004 |
| Leptogium burnetii Dodge | Forest-pasture | 3.3 | 1.000 |
| Leptogium cochleatum (Dicks.) P.M. Jørg. & P. James | Forest | 24.1 | 0.0002 |
| Leptogium coralloideum (Meyen & Flot.) Vain. | Forest | 11.1 | 0.0388 |
| Leptogium corticola (Taylor) Tuck. | Forest | 15.9 | 0.0036 |
| Leptogium cyanescens (Rabh.) Körb. | Forest-pasture | 5.7 | 0.3835 |
| Leptogium diaphanum (Sw.) Nyl. | Forest-pasture | 20.3 | 0,019 |
| Leptogium laceroides B. de Lesd. | Forest | 10 | 0.0466 |
| Leptogium marginellum (Sw.) Gray | Forest | 10 | 0.0468 |
| Leptogium millegranum Sierk | Urban | 9.8 | 0.0558 |
| Leptogium olivaceum (Hook.) Zahlbr. | Forest | 6.7 | 0,2388 |
| Leptogium phyllocarpum (Pers.) Mont. | Urban | 6.7 | 0.2442 |
| Leptogium sp. | Forest | 13.3 | 0.025 |
| Lobaria erosa (Eschw.) Nyl. | Forest | 2,8 | 1.000 |
| Lobaria subexornata (Yoshim.) Yoshim. | Forest | 16.7 | 0.0024 |
| Megalospora melanodermia (Müll. Arg.) Zahlbr. | Forest | 13.3 | 0.0134 |
| Normandina pulchella (Borrer) Nyl. | Forest | 13.3 | 0.0134 |
| Parmotrema arnoldii (Du Rietz) Hale | Forest-pasture | 4.6 | 0.8822 |
| Parmotrema austrosinense (Zahlbr.) Hale | Pasture | 13.8 | 0.067 |
| Parmotrema chinense (Osbeck) Hale y Ahti | Urban | 5.6 | 0.3569 |
| Parmotrema conferendum Hale | Forest-pasture | 7 | 0.2651 |
| Parmotrema cristiferum (Taylor) Hale | Forest | 8.1 | 0.1466 |
| Parmotrema exquisitum (Kurok.) DePriest & B.W. Hale | Forest-pasture | 4.7 | 0.5609 |
| Parmotrema peralbidum (Hale) Hale | Forest-pasture | 3.1 | 1.000 |
| Parmotrema subtinctorium (Zahlbr.) Hale | Forest | 2.5 | 0.6121 |
| Parmotrema sp. | Forest-pasture | 12.4 | 0.0242 |
| Pannaria conoplea (Ach.) Bory | Forest-pasture | 21.3 | 0.0042 |
| Pertusaria sp. | Forest | 3.3 | 1.000 |
| Pertusaria sp. 1 | Forest | 6.7 | 0.242 |
| Pertusaria sp. 2 | Urban | 6.8 | 0.1912 |
| Pertusaria sp. 3 | Forest | 6.7 | 0.2442 |
| Pertusaria sp. 4 | Forest | 6.7 | 0.232 |
| Pertusaria sp. 5 | Forest-pasture | 3.3 | 1.000 |
| Pertusaria sp. 6 | Forest-pasture | 1,7 | 1.000 |
| Phaeographis brasiliensis (A. Massal.) Kalb & Matthes-Leicht | Forest | 8.8 | 0.0598 |
| Phaeographis decipiens Müll. Arg. | Pasture | 3.3 | 1.000 |
| Phaeographis dendritica (Ach.) Müll.Arg. | Forest-pasture | 2.3 | 1.000 |
| Phaeographis intricans (Nyl.) Vain. | Forest | 6.7 | 0,2476 |
| Phaeographis inusta (Ach.) Müll. Arg. | Pasture | 3.3 | 1.000 |
| Phaeographis quadrifera (Nyl.) Staiger | Forest-pasture | 15,4 | 0,0052 |
| Phaeographis punctiformis (Eschw.) Müll. Arg. | Forest | 3.3 | 1.000 |
| Phaeographis subtigrina (Vain.) Zahlbr. | Forest-pasture | 2.3 | 0.9884 |
| Phaeophyscia aff. limbata (Poelt) Kashiw. | Pasture | 9.2 | 0.1222 |
| Phaeophyscia sp. | Forest | 11.1 | 0.2202 |
| Phyllopsora isidiotyla (Vain.) Riddle | Pasture | 30.9 | 0.0002 |
| Physcia aipolia (Ehrh. Ex Humb.) Fürnr. | Urban | 12 | 0.0372 |
| Physcia atrostriata Moberg | Pasture | 3.3 | 1.000 |
| Physcia crispa Nyl. | Urban | 10.9 | 0.0638 |
| Physcia endochrysea (Nyl.) Hampe | Urban | 14.9 | 0.0088 |
| Physcia poncinsii Hue | Urban | 6.7 | 0.2446 |
| Physcia sorediosa (Vain.) | Urban | 6.7 | 0.248 |
| Physcia sp. | Urban | 3.3 | 0.6163 |
| Physcia sp. 1 | Urban | 3.3 | 1.000 |
| Physcia sp. 2 | Pasture | 5.2 | 0.2494 |
| Pseudocyphellaria aurata (Ach.) Vain. | Forest-pasture | 11.9 | 0.0144 |
| Pseudocyphellaria crocata (L.) Vain. | Pasture | 10.3 | 0.0998 |
| Punctelia aff. crispa Marcelli, Jungbluth & Elix | Forest-pasture | 3.3 | 1.000 |
| Punctelia reddenda (Stirt.) Krog | Forest-pasture | 10 | 0.0616 |
| Punctelia rudecta (Ach.) Krog | Forest | 4.2 | 0.6065 |
| Punctelia subrudecta (Nyl.) Krog | Forest | 14.9 | 0.0088 |
| Punctelia sp. | Forest | 2.0 | 1.000 |
| Punctelia sp. 1 | Forest | 3.3 | 1.000 |
| Punctelia sp. 2 | Forest-pasture | 2.1 | 0.912 |
| Pyrenula sp | Forest-pasture | 10.0 | 0.053 |
| Pyrenula sp1. | Forest-pasture | 11.9 | 0.0932 |
| Pyxine cocoës (Sw.) Nyl. | Forest | 10.0 | 0.0616 |
| Ramalina celastri (Spreng.) Krog & Swinscow | Urban | 12.8 | 0,0132 |
| Ramalina cochlearis Zahlbr. | Urban | 2.2 | 1.000 |
| Ramalina peruviana Ach. | Forest-pasture | 4.0 | 0.6189 |
| Rimelia reticulata (Taylor) Hale | Forest-pasture | 2.1 | 1.000 |
| Rimelia subisidiosa (Müll. Arg.) Hale | Pasture | 3.7 | 0,7261 |
| Rinodina sp. | Forest | 11.4 | 0,033 |
| Sticta andensis (Nyl.) Trevis. | Forest | 3.3 | 1.000 |
| Sticta ferax Müll. Arg. | Forest | 16.7 | 0.0028 |
| Sticta fuliginosa (Dicks.) Ach. | Forest | 39.8 | 0.0002 |
| Sticta humboldtii Hook. f. | Forest | 6.7 | 0.2488 |
| Sticta tomentosa (Sw.) Ach. | Forest | 20 | 0.0008 |
| Sticta sp. | Pasture | 13.3 | 0.015 |
| Teloschistes flavicans (Sw.) Norman | Urban | 6.9 | 0.1842 |
| Teloschistes chrysophthalmus (L.) Beltr. | Pasture | 2.5 | 0.8224 |
| Teloschistes exilis (Michaux) Vain. | Forest-pasture | 9.6 | 0.5153 |
| Teloschistes hypoglaucus (Nyl.) Zahlbr. | Urban | 1.7 | 1.000 |
| Usnea sp. | Forest-pasture | 14.9 | 0.0336 |
| Usnea sp. 1 | Urban | 2.4 | 1.000 |
| Usnea sp. 2 | Pasture | 8.2 | 0.3405 |
| Usnea sp. 3 | Urban | 16.4 | 0.0188 |
| Usnea sp. 4 | Urban | 9.0 | 0.1246 |
| Usnea sp. 5 | Urban | 5.3 | 0.4397 |
| Usnea sp. 6 | Forest-pasture | 3.9 | 0.6755 |
| Usnea sp. 7 | Urban | 6.0 | 0.2809 |
References
- Turner, W.; Nakamura, T.; Dinetti, M. Global Urbanization and the Separation of Humans from Nature. BioScience 2004, 54, 585. [Google Scholar] [CrossRef] [Green Version]
- Gibson, L.; Lee, T.M.; Koh, L.P.; Brook, B.W.; Gardner, T.A.; Barlow, J.; Sodhi, N.S. Primary forests are irreplaceable for sustaining tropical biodiversity. Nature 2011, 478, 378. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Shi, J. Urbanization and its environmental effects in Shanghai, China. Urban Clim. 2012, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Day, J. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45. [Google Scholar] [CrossRef]
- Stofer, S.; Bergamini, A.; Aragon, G.; Carvalho, P.; Coppins, B.J.; Davey, S.; Lökös, L. Species richness of lichen functional groups in relation to land use intensity. Lichenologist 2006, 38, 331–353. [Google Scholar] [CrossRef]
- Liira, J.; Sepp, T.; Parrest, O. The forest structure and ecosystem quality in conditions of anthropogenic disturbance along productivity gradient. For. Ecol. Manag. 2007, 250, 34–46. [Google Scholar] [CrossRef]
- Gowda, J.H.; Kitzberger, T.; Premoli, A.C. Landscape responses to a century of land use along the northern Patagonian forest-steppe transition. Plant Ecol. 2012, 213, 259–272. [Google Scholar] [CrossRef]
- Mckinney, M. Urbanization, Biodiversity, and Conservation: The impacts of urbanization on native species are poorly studied but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; de la Cruz, M. Deforestation and forest fragmentation in South Ecuador since the 1970s–losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef]
- Benítez, Á.; Prieto, M.; González, Y.; Aragón, G. Effects of tropical montane forest disturbance on epiphytic macrolichens. Sci. Total Environ. 2012, 441, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Alberti, M. Eco-evolutionary dynamics in an urbanizing planet. Trends Ecol. Evol. 2015, 30, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Iñiguez-Armijos, C.; Hampel, H.; Breuer, L. Land-use effects on structural and functional composition of benthic and leaf-associated macroinvertebrates in four Andean streams. Aquat. Ecol. 2018, 52, 77–92. [Google Scholar] [CrossRef]
- Holz, I.; Gradstein, R.S. Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica–species richness, community composition and ecology. Plant Ecol. 2005, 178, 89–109. [Google Scholar] [CrossRef]
- Nöske, N.M.; Hilt, N.; Werner, F.A.; Brehm, G.; Fiedler, K.; Sipman, H.J.; Gradstein, S.R. Disturbance effects on diversity of epiphytes and moths in a montane forest in Ecuador. Basic. Appl. Ecol. 2008, 9, 4–12. [Google Scholar] [CrossRef]
- Hauck, M.; Lkhagvadorj, D. Epiphytic lichens as indicators of grazing pressure in the Mongolian forest-steppe. Ecol. Indic. 2013, 32, 82–88. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Solhaug, K.A.; Longinotti, S. Functional traits prolonging photosynthetically active periods in epiphytic cephalolichens during desiccation. Environ. Exp. Bot. 2017, 141, 83–91. [Google Scholar] [CrossRef]
- Giordani, P. Variables influencing the distribution of epiphytic lichens in heterogeneous areas: A case study for Liguria, NW Italy. J. Veg. Sci. 2006, 17, 195–206. [Google Scholar] [CrossRef]
- Benítez, Á.; Prieto, M.; Aragón, G. Large trees and dense canopies: Key factors for maintaining high epiphytic diversity on trunk bases (bryophytes and lichens) in tropical montane forests. For. J. For. Res 2015, 88, 521–527. [Google Scholar] [CrossRef]
- Giordani, P.; Brunialti, G.; Bacaro, G.; Nascimbene, J. Functional traits of epiphytic lichens as potential indicators of environmental conditions in forest ecosystems. Ecol. Indic. 2012, 18, 413–420. [Google Scholar] [CrossRef]
- Pinho, P.; Bergamini, A.; Carvalho, P.; Branquinho, C.; Stofer, S.; Scheidegger, C.; Maguas, C. Lichen functional groups as ecological indicators of the effects of land-use in Mediterranean ecosystems. Ecol. Indic. 2012, 15, 36–42. [Google Scholar] [CrossRef]
- Benítez, A.; Aragón, G.; González, Y.; Prieto, M. Functional traits of epiphytic lichens in response to forest disturbance and as predictors of total richness and diversity. Ecol. Indic. 2018, 86, 18–26. [Google Scholar] [CrossRef]
- Koch, N.M.; Matos, P.; Branquinho, C.; Pinho, P.; Lucheta, F.; de Azevedo Martins, S.M.; Vargas, V.M.F. Selecting lichen functional traits as ecological indicators of the effects of urban environment. Sci. Total Environ. 2019, 654, 705–713. [Google Scholar] [CrossRef]
- Merinero, S.; Martínez, I.; Rubio-Salcedo, M.; Gauslaa, Y. Epiphytic lichen growth in Mediterranean forests: Effects of proximity to the ground and reproductive stage. Basic. Appl. Ecol. 2015, 16, 220–230. [Google Scholar] [CrossRef]
- Giordani, P.; Rizzi, G.; Caselli, A.; Modenesi, P.; Malaspina, P.; Mariotti, M.G. Fire affects the functional diversity of epilithic lichen communities. Fungal Ecol. 2016, 20, 49–55. [Google Scholar] [CrossRef]
- Koch, N.M.; de Azevedo Martins, S.M.; Lucheta, F.; Müller, S.C. Functional diversity and traits assembly patterns of lichens as indicators of successional stages in a tropical rainforest. Ecol. Indic. 2013, 34, 22–30. [Google Scholar] [CrossRef]
- Hauck, M.; de Bruyn, U.; Leuschner, C. Dramatic diversity losses in epiphytic lichens in temperate broad-leaved forests during the last 150 years. Biol. Conserv. 2013, 157, 136–145. [Google Scholar] [CrossRef]
- Zarabska-Bożejewicz, D.; Kujawa, K. The effect of land use on taxonomical and functional diversity of lichens in an agricultural landscape. Fungal Ecol. 2018, 33, 72–79. [Google Scholar] [CrossRef]
- Aragón, G.; Martínez, I.; Hurtado, P.; Benítez, Á.; Rodríguez, C.; Prieto, M. Using Growth Forms to Predict Epiphytic Lichen Abundance in a Wide Variety of Forest Types. Diversity 2019, 11, 51. [Google Scholar] [CrossRef]
- Gauslaa, Y. Rain, dew, and humid air as drivers of morphology, function and spatial distribution in epiphytic lichens. Lichenologist 2014, 46, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Llop, E.; Pinho, P.; Matos, P.; Pereira, M.J.; Branquinho, C. The use of lichen functional groups as indicators of air quality in a Mediterranean urban environment. Ecol. Indic. 2012, 13, 215–221. [Google Scholar] [CrossRef]
- Ochoa, D.; Cueva, A.; Prieto, M.; Aragón, G.; Benitez, A. Cambios en la composición de líquenes epifitos relacionados con la calidad del aire en la cuidad de Loja (ECUADOR). Caldasia 2015, 37, 333–343. [Google Scholar] [CrossRef]
- Benítez, Á.; Medina, J.; Vásquez, C.; Loaiza, T.; Luzuriaga, Y.; Calva, J. Lichens and Bromeliads as Bioindicators of Heavy Metal Deposition in Ecuador. Diversity 2019, 11, 28. [Google Scholar] [CrossRef]
- Käffer, M.I.; de Azevedo, S.M.; Alves, C.; Pereira, V.C.; Fachel, J.; Vargas, V.M.F. Corticolous lichens as environmental indicators in urban areas in southern Brazil. Ecol. Indic. 2011, 11, 1319–1332. [Google Scholar] [CrossRef]
- Koch, N.M.; Branquinho, C.; Matos, P.; Pinho, P.; Lucheta, F.; Martins, S.M.; Vargas, V.M. The application of lichens as ecological surrogates of air pollution in the subtropics: A case study in South Brazil. Environ. Sci. Pollut. Res. 2016, 23, 20819–20834. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Armijos, M.F.; Homeier, J.; Draper-Munt, D. Spatio-temporal analysis of the human footprint in South Ecuador: Influence of human pressure on ecosystems and effectiveness of protected areas. Appl. Geogra. 2017, 78, 22–32. [Google Scholar] [CrossRef]
- Iñiguez-Armijos, C.L.; Frede, H.G.; Hampel, H.; Breuer, L. Deforestation and benthic indicators: How much vegetation cover is needed to sustain healthy Andean streams? PLoS ONE 2014, 9, e105869. [Google Scholar]
- Iñiguez-Armijos, C.; Rausche, S.; Cueva, A.; Sánchez-Rodriguez, A.; Espinosa, C.; Breuer, L. Shifts in leaf litter breakdown along a forest–pasture–urban gradient in Andean streams. Ecol. Evol. 2016, 6, 4849–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollenbeck, R.; Bendix, J. Rainfall distribution in the Andes of southern Ecuador derived from blending weather radar data and meteorological field observations. Atmos. Res. 2011, 99, 277–289. [Google Scholar] [CrossRef]
- May, P.F.; Brodo, I.M.; Esslinger, T.L. Identifying North American lichens: A Guide to the Literature; Farlow Herbarium, Harvard University: Cambridge, MA, USA, 2002. [Google Scholar]
- Sipman, H.J.M. Identification Key and Literature Guide to the Genera of Lichenized Fungi (Lichens) in the Neotropics, Provisional Version. Botanical Garden & Botanical Museum Berlin-Dahlen, Free University of Berlin, 2003. Available online: http://www.bgbm.fu-berlin.de/sipman/keys/neokeyA.htm (accessed on 10 January 2018).
- Cáceres, M.E.S.; Lücking, R.; Rambold, G. Phorophyte specificity and environmental parameters as determinants for species composition, richness and area cover in corticolous crustose lichen communities in the Atlantic rainforest of northeastern Brazil. Mycol. Progr. 2007, 6, 117–136. [Google Scholar] [CrossRef]
- Moncada, B.; Lücking, R.; Suárez, A. Molecular phylogeny of the genus Sticta (lichenized Ascomycota: Lobariaceae) in Colombia. Fungal Divers. 2014, 64, 205–231. [Google Scholar] [CrossRef]
- Lange, O.L.; Kilian, E.; Ziegler, H. Water vapor uptake and photosynthesis of lichens: Performance differences in species with green and blue-green algae as phycobionts. Oecologia 1986, 71, 104–110. [Google Scholar] [CrossRef]
- Lakatos, M.; Rascher, U.; Büdel, B. Functional characteristics of corticolous lichens in the understory of a tropical lowland rain forest. New Phytol. 2006, 172, 679–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef] [PubMed]
- Frazer, G.W.; Canham, C.D.; Lertzman, K.P. Gap Light Analyzer (GLA), Version 2.0: Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, Users Manual and Program Documentation; Simon Fraser University: Burnaby, British Columbia; The Institute of Ecosystem Studies: Millbrook, NY, USA, 1999; Volume 36. [Google Scholar]
- McCullagh, P.; Nelder, J.P. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989; p. 511. [Google Scholar]
- Warton, D.I.; Hui, F.K. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and Nonlinear Mixed Effects Models. R Package Vers 1.3-5. 2017. Available online: http://CRAN.R-project.org/package=nlme (accessed on 10 January 2018).
- R Core Team. R: A Language Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.r-project.org/ (accessed on 10 January 2018).
- Hill, M.O.; Gauch, H.G. Detrended correspondence analysis: An improved ordination technique. In Classification and Ordination; Springer: Berlin/Heidelberg, Germany, 1980; pp. 47–58. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Oksanen, J.; Kindt, R.; Simpson, G.L. Package ‘vegan3d’. 2018. Available online: http://CRAN.R-project.org/package=vegan3d (accessed on 10 January 2018).
- Roberts, D.W. Labdsv: Ordination and Multivariate Analysis for Ecology. Version 1.4-1. 2012. Available online: http://CRAN.R-project.org/package=labdsv (accessed on 10 January 2018).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Ellis, C.J.; Coppins, B.J. 19th century woodland structure controls stand-scale epiphyte diversity in present-day Scotland. Divers. Distrib. 2007, 13, 84–91. [Google Scholar] [CrossRef]
- Baniya, C.B.; Solhøy, T.; Gauslaa, Y.; Palmer, M.W. The elevation gradient of lichen species richness in Nepal. Lichenologist 2010, 42, 83–96. [Google Scholar] [CrossRef]
- Aragón, G.; Martínez, I.; Izquierdo, P.; Belinchón, R.; Escudero, A. Effects of forest management on epiphytic lichen diversity in Mediterranean forests. Appl. Veg. Sci. 2010, 13, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Ellis, C.J.; Coppins, B.J. Partitioning the role of climate, pollution and old-growth woodland in the composition and richness of lichen epiphytes in Scotland. Lichenologist 2010, 42, 601–614. [Google Scholar] [CrossRef]
- Chongbang, T.B.; Keller, C.; Nobis, M.; Scheidegger, C.; Baniya, C.B. From natural forest to cultivated land: Lichen species diversity along land-use gradients in Kanchenjunga, Eastern Nepal. Ecol. Mont 2018, 1, 46–60. [Google Scholar] [CrossRef]
- Moning, C.; Werth, S.; Dziock, F.; Bässler, C.; Bradtka, J.; Hothorn, T.; Müller, J. Lichen diversity in temperate montane forests is influenced by forest structure more than climate. For. Ecol. Manag. 2009, 258, 745–751. [Google Scholar] [CrossRef]
- Aragón, G.; Martínez, I.; García, A. Loss of epiphytic diversity along a latitudinal gradient in southern Europe. Sci. Total Environ. 2012, 426, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Matos, P.; Pinho, P.; Aragon, G.; Martínez, I.; Nunes, A.; Soares, A.M.; Branquinho, C. Lichen traits responding to aridity. J. Ecol. 2015, 103, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Leppik, E.; Jueriado, I.; Liira, J. Changes in stand structure due to the cessation of traditional land use in wooded meadows impoverish epiphytic lichen communities. Lichenologist 2011, 43, 257–274. [Google Scholar] [CrossRef]
- Lange, O.; Büdel, A.; Meyer, A.; Kilian, E. Further evidence that activation of net photosynthesis by dry cyanobacterial lichens requires liquid water. Lichenologist 1993, 25, 175–189. [Google Scholar] [CrossRef]
- Kiebacher, T.; Keller, C.; Scheidegger, C.; Bergamini, A. Epiphytes in wooded pastures: Isolation matters for lichen but not for bryophyte species richness. PLoS ONE 2017, 12, e0182065. [Google Scholar] [CrossRef] [PubMed]




| Functional Trait | Categories |
|---|---|
| Photobiont type | C = Chlorococcoid; CY = Cyanobacteria: T = Trentepohlia |
| Growth form | CR = Crustose; F = Filamentose; FB = Foliose with broad lobes; FN = Foliose with narrow lobes; FR = Fruticose; G = Gelatinose |
| Thallus size | MA = Macrolichens; MI = Microlichens |
| Thallus colour | L = Light; D = Dark |
| Reproduction type | A = Asexual; S = Sexual; AS = Asexual and sexual |
| Reproductive structure | AP = Apothecia; I = Isidia; L = Lirellae; P = Perithecia; SO = Soredia |
| Source of Variation | Coefficient | Standard Error | Z-value | p-value |
|---|---|---|---|---|
| Forest | 2.651 | 0.297 | 8.908 | <0.001 |
| Forest-Pasture | −0.663 | 0.286 | −2.313 | 0.020 |
| Pasture | −0.976 | 0.336 | −2.899 | 0.003 |
| Urban | −1.661 | 0.537 | −3.093 | 0.001 |
| Distance to forest | 0.445 | 0.160 | 2.767 | 0.005 |
| DBH | −0.004 | 0.056 | 0.078 | 0.937 |
| Source of Variation | Degrees of Freedom | Chi Square | F-statistic | p-value |
|---|---|---|---|---|
| Land use | 3 | 1.196 | 2.316 | 0.001 |
| Precipitation | 1 | 0.242 | 1.410 | 0.003 |
| Distance to forest | 1 | 0.283 | 1.6477 | 0.001 |
| % Forest | 1 | 0. 245 | 1.423 | 0.014 |
| Functional Traits | Land-use Intensity | DBH | Dist-F | %Fo | |||
|---|---|---|---|---|---|---|---|
| Fo | FP | Pa | Ur | ||||
| Photobiont type | |||||||
| Chlorococcoid | 1.883 (0.004) | ||||||
| Cyanobacteria | 0.927 (0.001) | −0.871 (0.01) | −0.953 (0.01) | −1.195 (0.002) | |||
| Trentepohlia | −1.969 (0.03) | ||||||
| Growth form | |||||||
| Crustose | 1.033 (0.005) | −0.704 (0.04) | −0.977 (0.02) | −1.847 (0.01) | |||
| Filamentose | −6.699 (0.04) | 2.347 (0.02) | |||||
| Foliose with broad lobes | 1.277 (0.002) | −2.064 (0.01) | 0.789 (0.04) | ||||
| Foliose with narrow lobes | 0.911 (0.003) | ||||||
| Fruticose | −0.844 (0.02) | 1.007 (0.03) | 1.013 (0.04) | ||||
| Gelatinose | −1.028 (0.02) | −2.211 (0.01) | −2.13 (0.02) | ||||
| Thallus size | |||||||
| Macrolichens | 2.112 (0.009) | ||||||
| Microlichens | 1.553 (0.01) | 1.368 (0.006) | |||||
| Thallus colour | |||||||
| Light | 1.519 (0.001) | ||||||
| Dark | 1.818 (0.001) | −1.091 (0.02) | −1.17 (0.03) | ||||
| Reproduction type | |||||||
| Asexual | 1.539 (0.001) | −1.263 (0.004) | −1.929 (0.002) | −2.226 (0.001) | |||
| Sexual | 1.298 (0.001) | −0.818 (0.01) | −0.622 (0.04) | ||||
| Asexual and sexual | 0.775 (0.001) | ||||||
| Reproductive structure | |||||||
| Apothecia | 1.769 (0.001) | ||||||
| Isidia | 0.502 (0.006) | −0.492 (0.03) | −0.98 (0.005) | −1.281 (0.008) | 0.271 (0.02) | ||
| Lirellae | |||||||
| Perithecia | |||||||
| Soredia | 1.099 (0.03) | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuquimarca, L.; Gaona, F.P.; Iñiguez-Armijos, C.; Benítez, Á. Lichen Responses to Disturbance: Clues for Biomonitoring Land-use Effects on Riparian Andean Ecosystems. Diversity 2019, 11, 73. https://doi.org/10.3390/d11050073
Chuquimarca L, Gaona FP, Iñiguez-Armijos C, Benítez Á. Lichen Responses to Disturbance: Clues for Biomonitoring Land-use Effects on Riparian Andean Ecosystems. Diversity. 2019; 11(5):73. https://doi.org/10.3390/d11050073
Chicago/Turabian StyleChuquimarca, Leiddy, Fernando P. Gaona, Carlos Iñiguez-Armijos, and Ángel Benítez. 2019. "Lichen Responses to Disturbance: Clues for Biomonitoring Land-use Effects on Riparian Andean Ecosystems" Diversity 11, no. 5: 73. https://doi.org/10.3390/d11050073
APA StyleChuquimarca, L., Gaona, F. P., Iñiguez-Armijos, C., & Benítez, Á. (2019). Lichen Responses to Disturbance: Clues for Biomonitoring Land-use Effects on Riparian Andean Ecosystems. Diversity, 11(5), 73. https://doi.org/10.3390/d11050073