Fewer Copepods, Fewer Anchovies, and More Jellyfish: How Does Hypoxia Impact the Chesapeake Bay Zooplankton Community?

 , ,
, ,
Abstract
:1. Introduction
1.1. Hypoxia in the Chesapeake Bay
1.2. Zooplankton and Planktivores Diversity in the Bay
1.3. Direct Effects of Seasonal Hypoxia
2. Methods
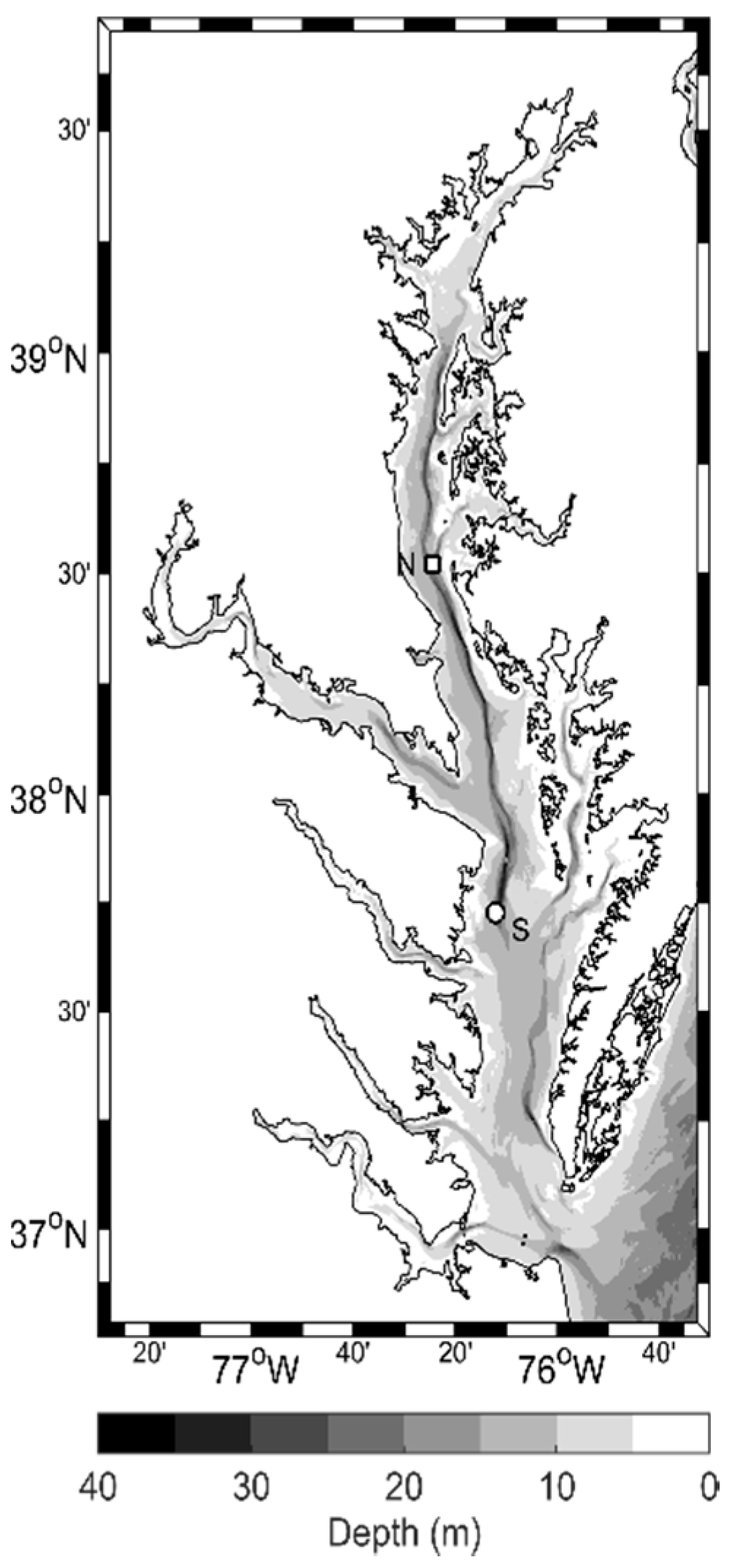
2.1. Cruises and Environmental Data
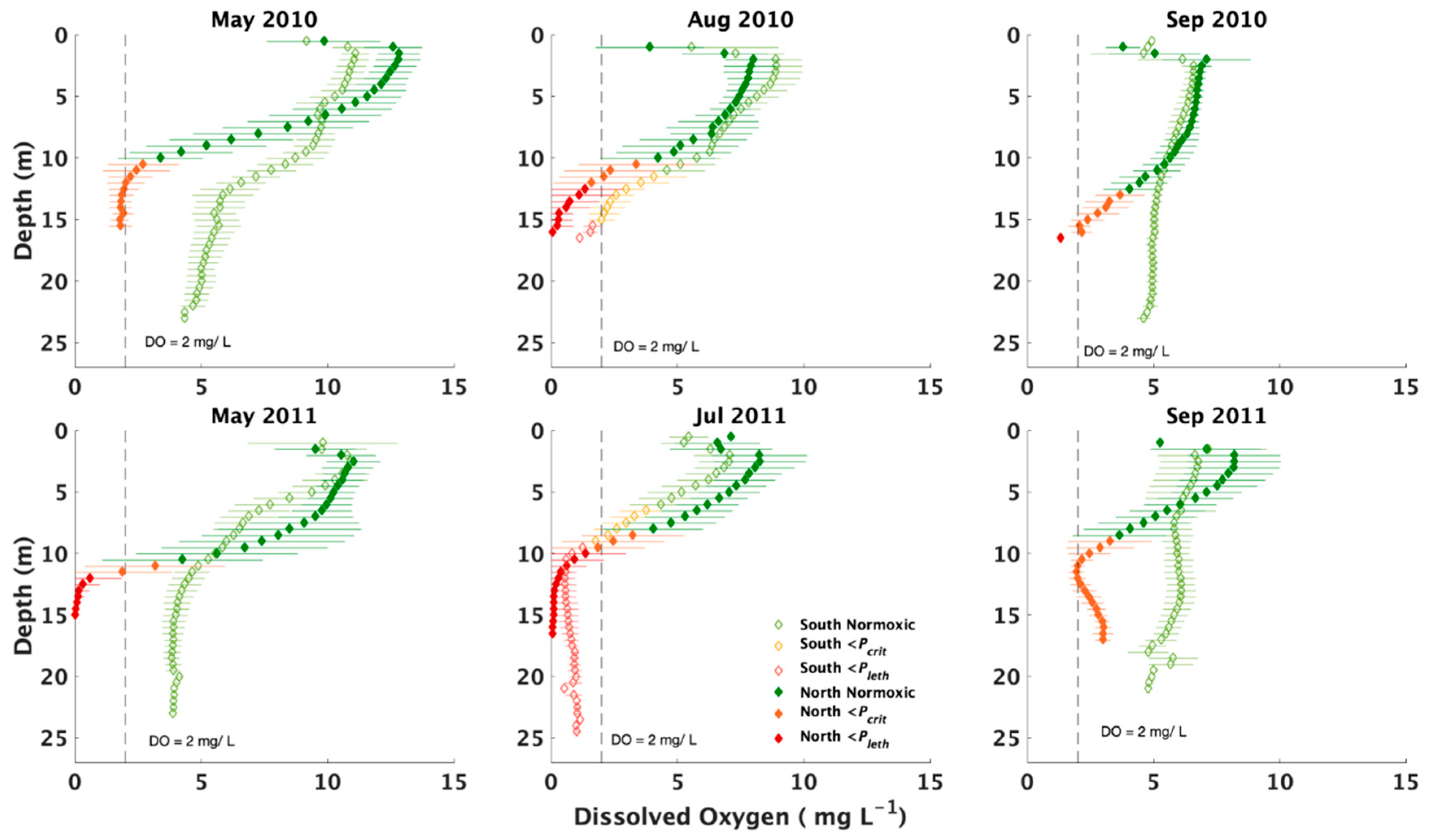
2.2. Evaluation of Environmental Oxygen Supplies and Copepod’s Physiological Oxygen Needs
2.3. Estimation of Zooplankton and Planktivorous Fish Concentrations
2.4. Nonpredatory Mortality Rates
2.5. Statistical Analysis
3. Results
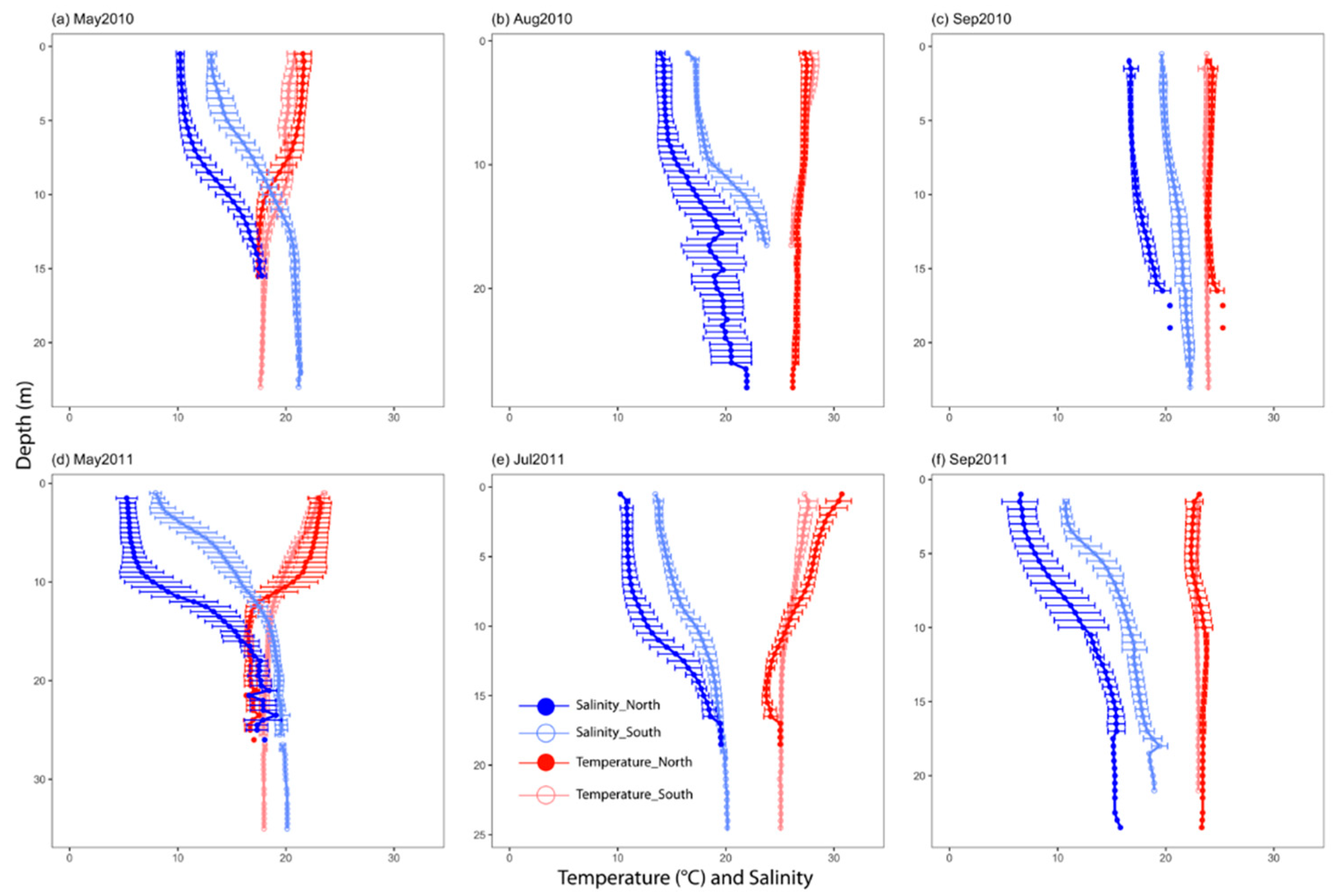
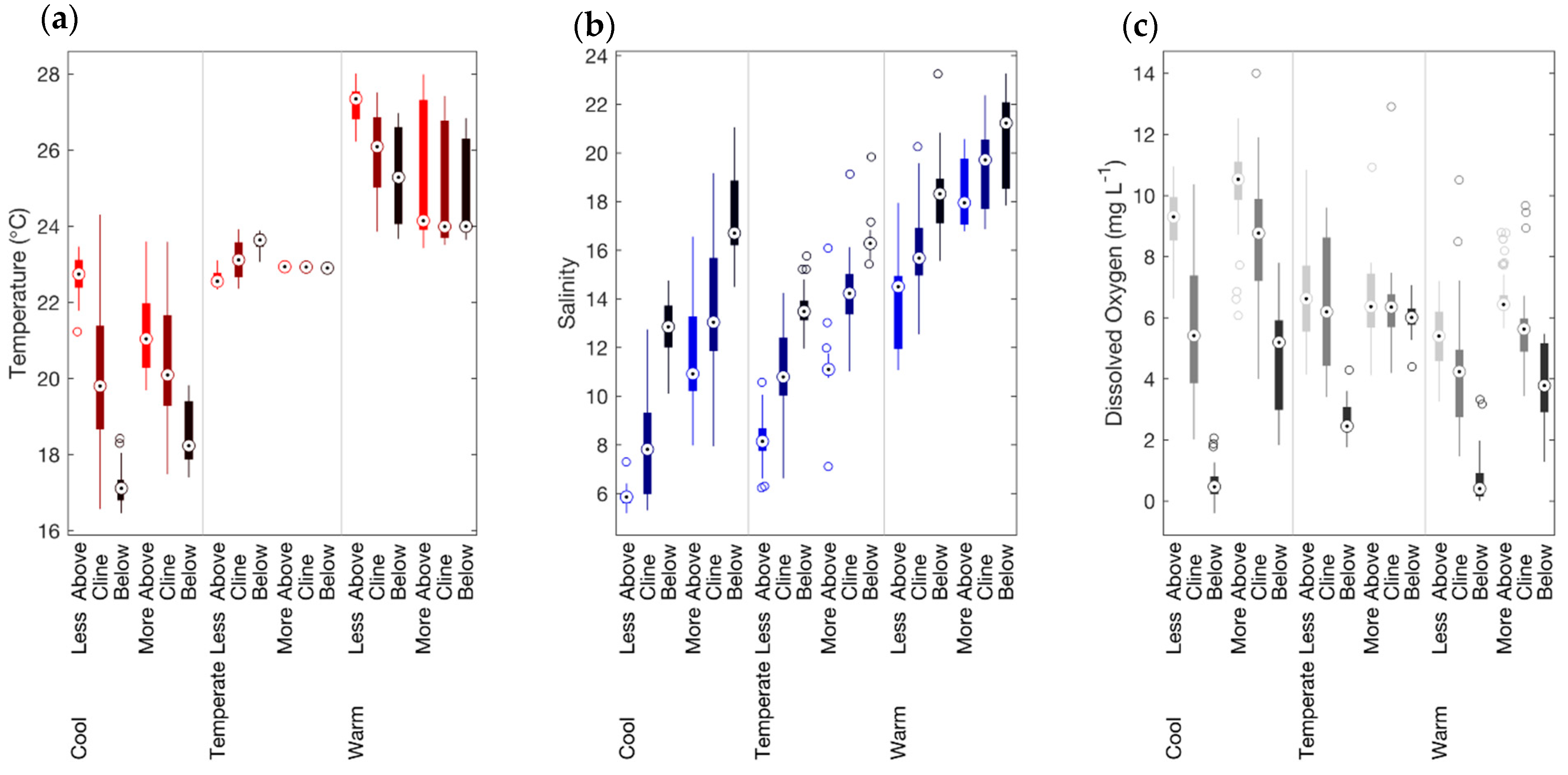
3.1. General Environmental Conditions
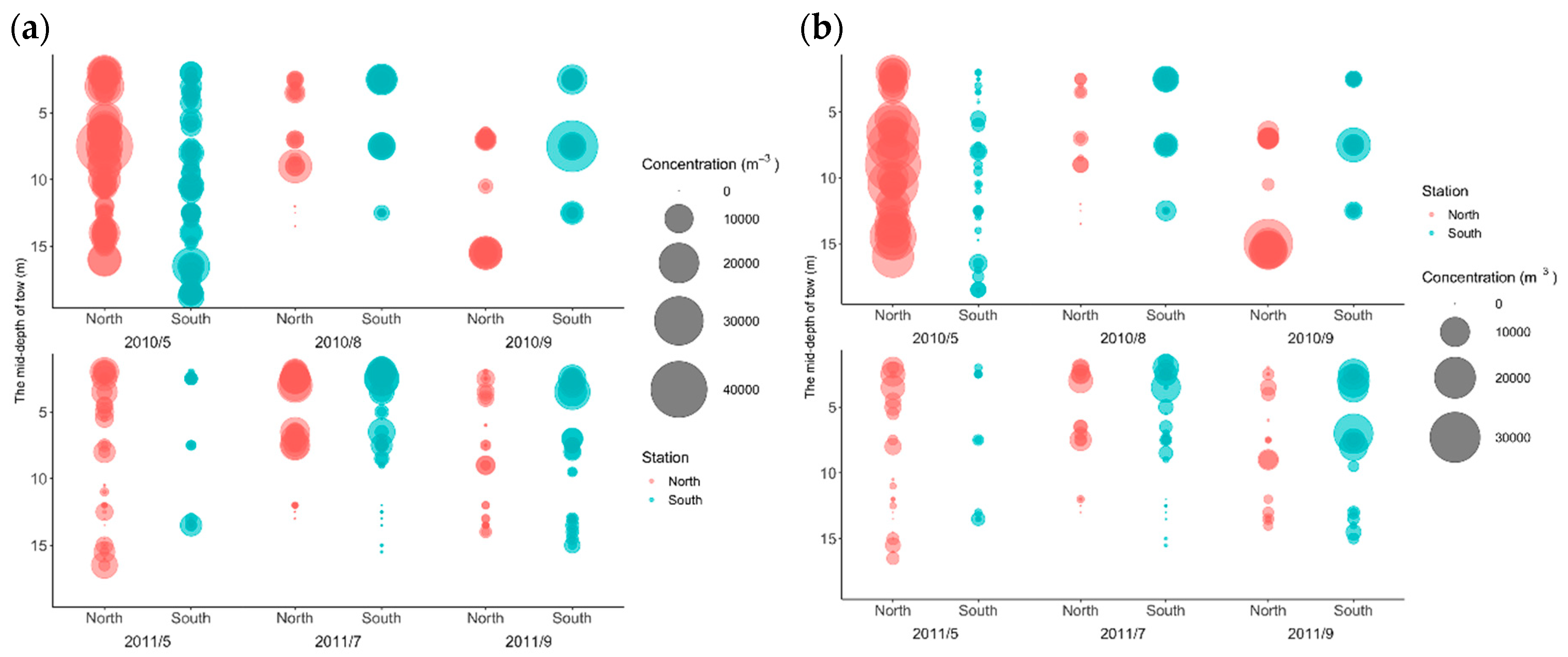
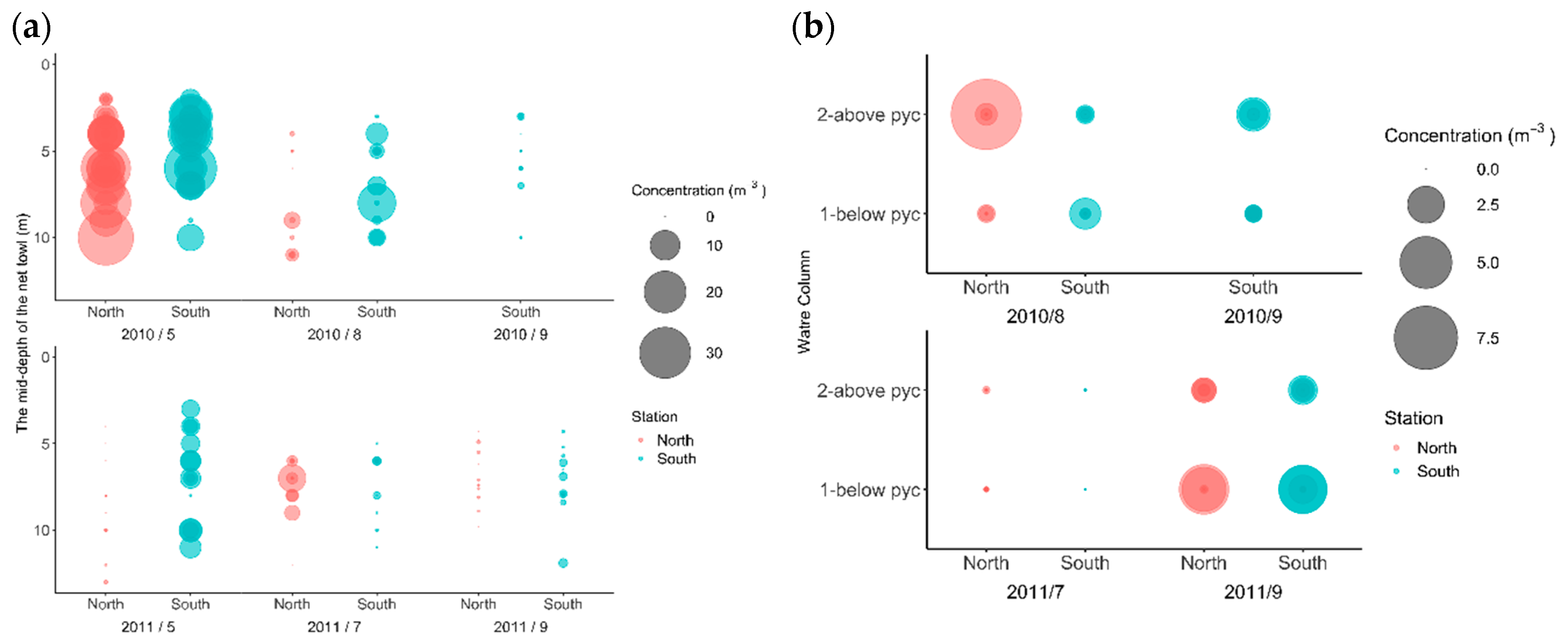
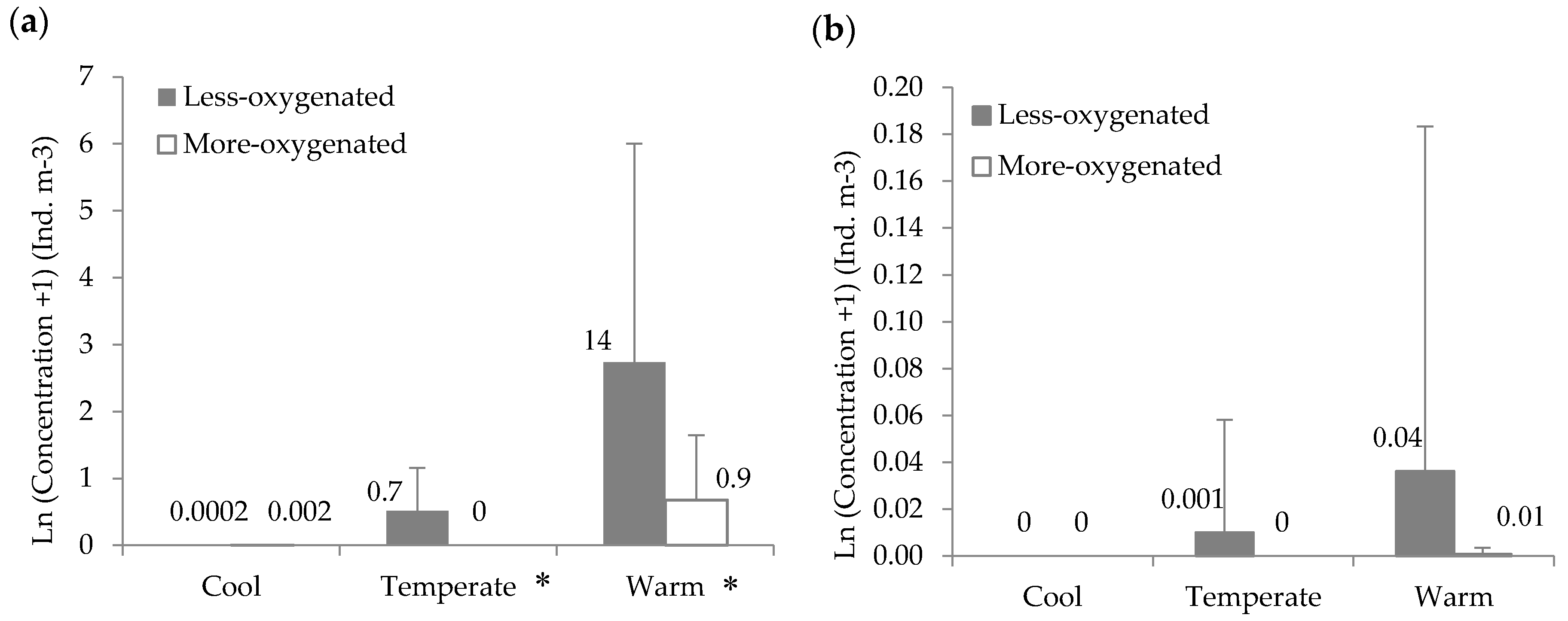
3.2. Zooplankton and Fish Concentrations
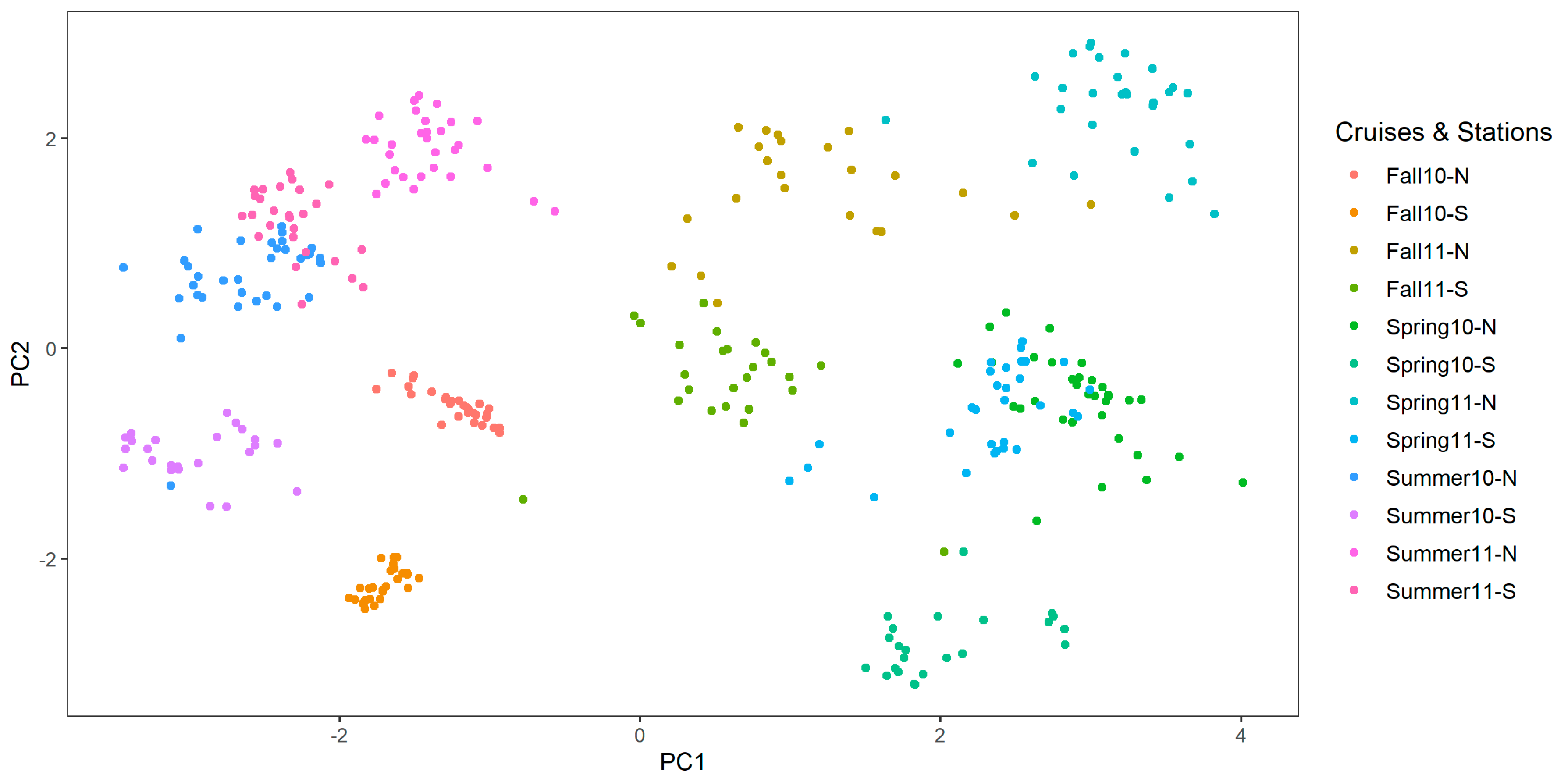
3.3. Grouping with PCA Results
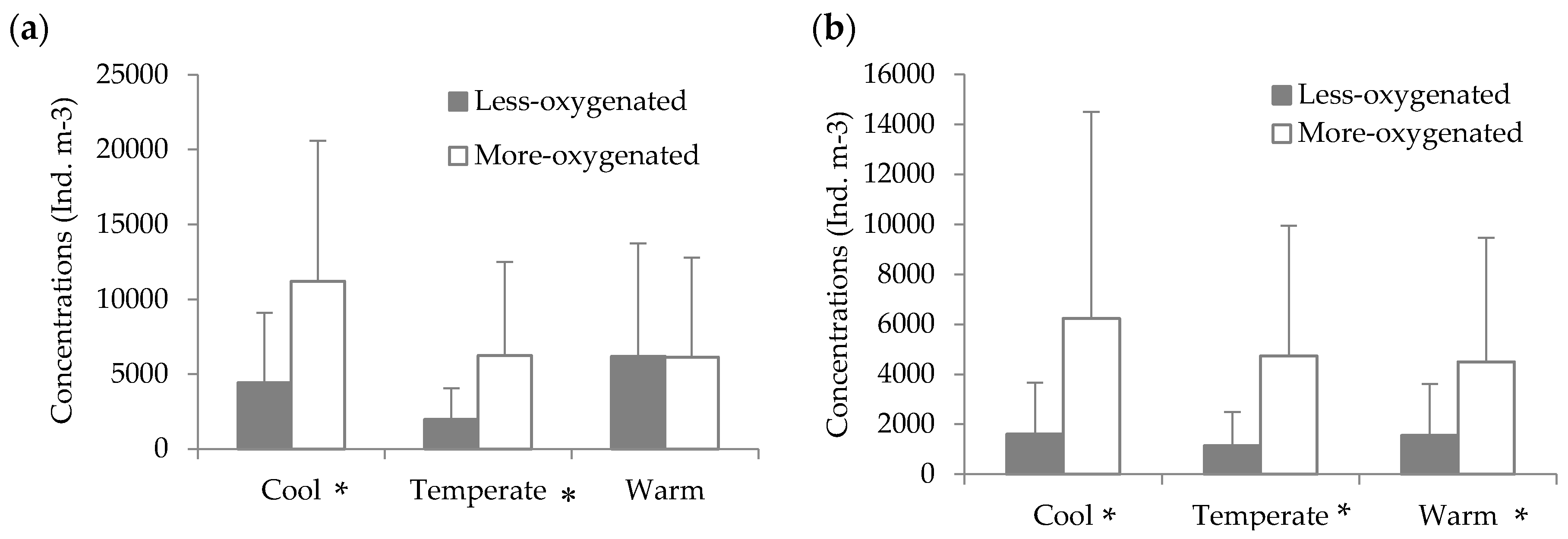
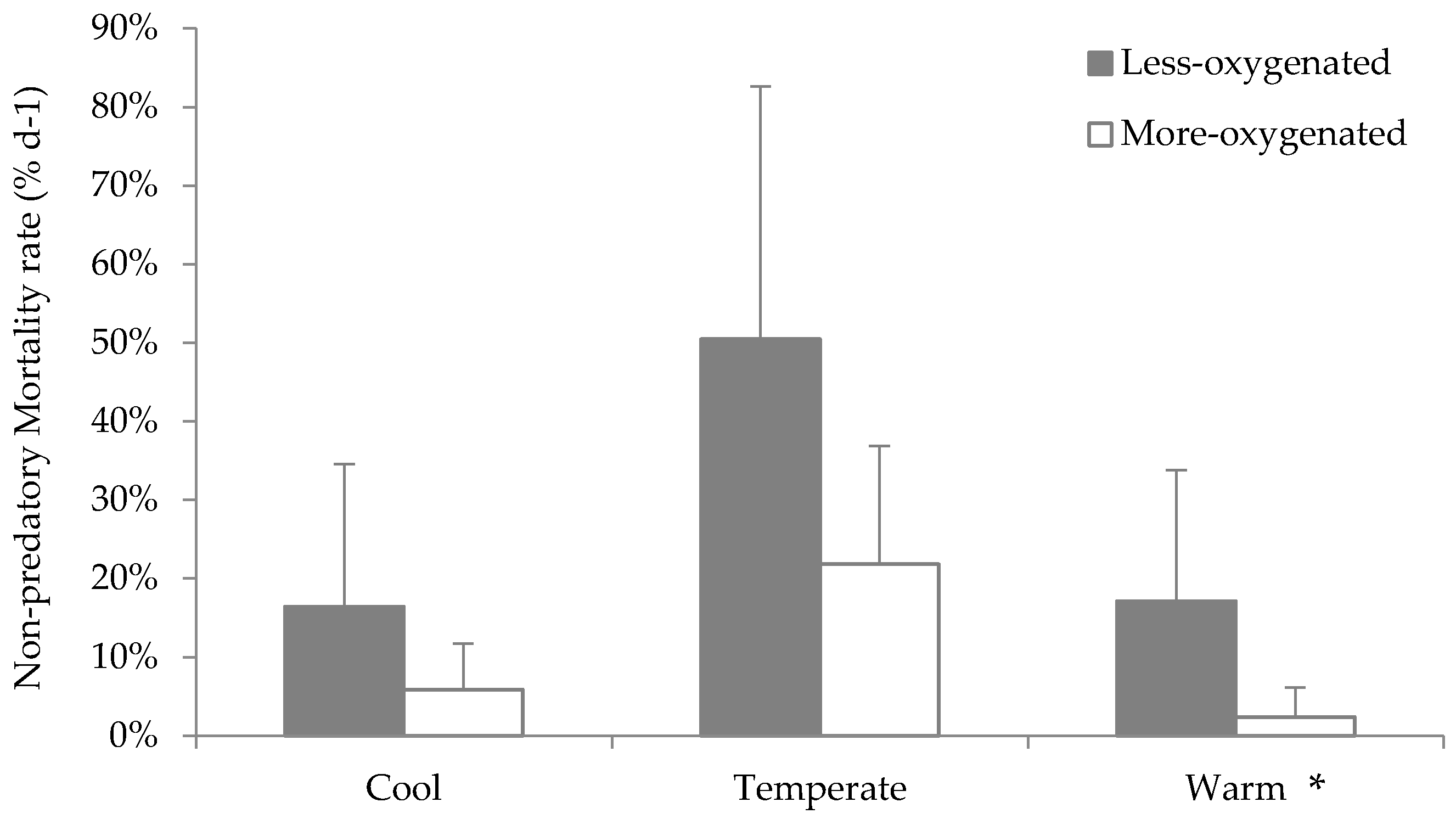
3.4. Effects of Hypoxia
4. Discussion
4.1. Hydrographical and Biological Hypoxia
4.2. Seasonal and Episodic Hypoxia
4.3. Strengths and Limitations of the PCA Grouping Method and Our Sampling Regime
4.4. Copepod’s Predators in Hypoxia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cruise | 2010 | 2011 |
|---|---|---|
| Season | ||
| Spring | 77 | 88 |
| Summer | 88 | 69 |
| Autumn | 64 | 66 |
| Total Casts | 229 | 223 |
| 452 |
| Cruise | MOCNESS | Tucker Trawls | Mid-Water Trawls |
|---|---|---|---|
| 2010 | |||
| Spring | 11 | 18 | 0 |
| Summer | 11 | 11 | 11 |
| Autumn | 6 | 6 | 6 |
| 2011 | |||
| Spring | 8 | 12 | 0 |
| Summer | 12 | 12 | 12 |
| Autumn | 12 | 12 | 12 |
| Total | 60 | 71 | 41 |
| Species/Stage | A. tonsa (Adult Female and Male) | A. tonsa (Copepodite) | A. mitchilli (Larval) | A. mitchilli (Juvenile) | M. leidyi (Adult) | C. chesapeakei (Medusa) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nets | MOCNESS | MOCNESS | MOCNESS | Mid-Water Trawl | Tucker Trawl | Tucker Trawl | |||||||
| Station | |||||||||||||
| North | May | 16,232.39 | ±10,908.58 | 12,656.34 | ±8514.45 | 11.46 | ±12.64 | - c | - | 0.0027 | ±0.0046 | 0 | 0 |
| Aug | 4077.83 | ±4882.37 | 981.92 | ±1011.33 | 0.59 | ±0.94 | 1.124 | ±2.874 | 1.0951 | ±1.3414 | 0.0019 | ±0.0035 | |
| Sep | 5592.79 a | ±5802.39 | 6244.62 a | ±6126.24 | - a | - | - a | - | - a | - | - a | - | |
| South | May | 7887.59 b | ±4587.74 | 839.89 b | ±1004.78 | 5.48 b | 7.6 | - c | - | 0.0007 | ±0.0035 | 0 | 0 |
| Aug | 6225.92 | ±7010.20 | 3231.59 | ±2624.41 | 2.95 | ±4.60 | 0.269 | 0.505 | 1.8699 | ±1.9468 | 0.0014 | ±0.0039 | |
| Sep | 7028.05 | ±7896.31 | 2595.67 | ±3111.16 | 0.12 | ±0.17 | 0.501 | 0.624 | 0.0671 | ±0.0670 | 0 | 0 | |
| Species/Stage | A. tonsa (Adult Female and Male) | A. tonsa (Copepodite) | A. mitchilli (Larval) | A. mitchilli (Juvenile) | M. leidyi (Adult) | C. chesapeakei (Medusa) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nets | MOCNESS | MOCNESS | MOCNESS | Mid-Water Trawl | Tucker Trawl | Tucker Trawl | |||||||
| Station | |||||||||||||
| North | May | 4426.36 | ±4682.72 | 1605.72 | ±2056.29 | 0.03 | ±0.04 | - c | - | 0.0002 | ±0.0012 | 0 | 0 |
| Jul | 9818.09 | ±8922.12 | 2000.93 | ±1977.54 | 1.42 | ±2.39 | 0.016 | ± 0.023 | 5.3006 | ±6.4913 | 0.0041 | ±0.0203 | |
| Sep | 1978.11 | ±2090.38 | 1139.02 | ±1356.40 | 0.03 | ±0.04 | 0.973 | ± 1.521 | 0.6708 | ±0.9117 | 0.0101 | ±0.0493 | |
| South | May | 2521.99 | ±2068.83 | 670.82 | ±556.18 | 4.45 | ±0.86 | - c | - | 0.0008 | ±0.0021 | 0 | 0 |
| Jul | 4949.34 | ±7132.56 | 1572.82 | ±2481.06 | 0.19 | ±0.28 | 0.001 | ± 0.002 | 34.6636 | ±33.5756 | 0.0988 | ±0.2581 | |
| Sep | 6258.11 | ±6235.51 | 4734.49 | ±5222.70 | 0.30 | ±0.31 | 1.138 | ± 1.569 | 0 | 0 | 0 | 0 | |
| Group | Sample Size | Layer | Temperature | Salinity | pO2 | Pcrit | Pleth | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LO | MO | LO | MO | LO | MO | LO | MO | LO | MO | LO | MO | ||||||||
| C | 28 | 87 | Surf. | 22.69 | ±0.53 | 21.19 | ±1.10 | 5.86 | ±0.40 | 11.56 | ±2.30 | 23.48 | ±2.32 | 26.79 | ±2.95 | 8.13 | 8.77 | 3.22 | 3.44 |
| Pyc. | 19.95 | ±1.81 | 20.39 | ±1.55 | 8.06 | ±2.22 | 13.57 | ±2.92 | 13.80 | ±5.39 | 21.87 | ±5.06 | 8.26 | 8.74 | 3.26 | 3.43 | |||
| Bot. | 17.18 | ±0.50 | 18.51 | ±0.75 | 12.76 | ±1.14 | 17.44 | ±1.75 | 1.48 | ±1.36 | 11.79 | ±4.33 | 7.88 | 8.27 | 3.13 | 3.27 | |||
| T | 23 | 26 | Surf. | 22.64 | ±0.23 | 22.94 | ±0.07 | 8.22 | ±1.12 | 11.22 | ±1.36 | 17.39 | ±3.93 | 17.14 | ±3.60 | 8.55 | 9.12 | 3.36 | 3.56 |
| Pyc. | 23.14 | ±0.49 | 22.91 | ±0.07 | 11.06 | ±1.92 | 14.30 | ±1.52 | 16.92 | ±5.65 | 17.33 | ±4.17 | 9.13 | 9.62 | 3.57 | 3.74 | |||
| Bot. | 23.61 | ±0.22 | 22.90 | ±0.06 | 13.61 | ±0.93 | 16.42 | ±0.79 | 7.36 | ±1.68 | 16.37 | ±1.36 | 9.73 | 9.95 | 3.78 | 3.85 | |||
| W | 87 | 86 | Surf. | 26.12 | ±1.44 | 26.27 | ±1.80 | 14.35 | ±2.27 | 17.50 | ±2.16 | 15.34 | ±2.82 | 19.18 | ±2.41 | 10.81 | 11.82 | 4.15 | 4.50 |
| Pyc. | 24.87 | ±0.89 | 25.95 | ±1.62 | 15.99 | ±1.51 | 19.05 | ±1.94 | 11.82 | ±4.59 | 15.68 | ±3.74 | 10.72 | 12.09 | 4.12 | 4.60 | |||
| Bot. | 24.39 | ±0.60 | 25.69 | ±1.34 | 17.85 | ±0.93 | 20.85 | ±1.53 | 4.06 | ±4.34 | 8.34 | ±5.61 | 10.91 | 12.41 | 4.19 | 4.71 | |||
References
- Rabalais, N.N.; Diaz, R.J.; Levin, L.A.; Turner, R.E.; Gilbert, D.; Zhang, J. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 2010, 7, 585–619. [Google Scholar] [CrossRef] [Green Version]
- Rhein, M.; Rintoul, S.R.; Aoki, S.; Campos, E.; Chambers, D.; Feely, R.A.; Gulev, S.; Johnson, G.C.; Josey, S.A.; Kostianoy, A.; et al. Observations: Ocean; Cambridge University Press: Cambridge, UK, 2013; pp. 255–315. [Google Scholar]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Libes, S. Introduction to Marine Biogeochemistry; Academic Press: Cambridge, MA, USA, 2011; ISBN 0-08-091664-3. [Google Scholar]
- Cowan, J.L.; Boynton, W.R. Sediment-water oxygen and nutrient exchanges along the longitudinal axis of Chesapeake Bay: Seasonal patterns, controlling factors and ecological significance. Estuaries 1996, 19, 562–580. [Google Scholar] [CrossRef]
- Hagy, J.D.; Boynton, W.R.; Keefe, C.W.; Wood, K.V. Hypoxia in Chesapeake Bay, 1950–2001: Long-term change in relation to nutrient loading and river flow. Estuaries 2004, 27, 634–658. [Google Scholar] [CrossRef]
- Kemp, W.M.; Boynton, W.R.; Adolf, J.E.; Boesch, D.F.; Boicourt, W.C.; Brush, G.; Cornwell, J.C.; Fisher, T.R.; Glibert, P.M.; Hagy, J.D.; et al. Eutrophication of Chesapeake Bay: Historical trends and ecological interactions. Mar. Ecol. Prog. Ser. 2005, 303, 1–29. [Google Scholar] [CrossRef]
- Murphy, R.R.; Kemp, W.M.; Ball, W.P. Long-term trends in Chesapeake Bay seasonal hypoxia, stratification, and nutrient loading. Estuaries Coasts 2011, 34, 1293–1309. [Google Scholar] [CrossRef]
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.; Brix, H.; Ito, T.; Frenzel, H.; Thompson, L. Climate-Forced Variability of Ocean Hypoxia. Science 2011, 333, 336–339. [Google Scholar] [CrossRef] [Green Version]
- Najjar, R.G.; Pyke, C.R.; Adams, M.B.; Breitburg, D.; Hershner, C.; Kemp, M.; Howarth, R.; Mulholland, M.R.; Paolisso, M.; Secor, D.; et al. Potential climate-change impacts on the Chesapeake Bay. Estuar. Coast. Shelf Sci. 2010, 86, 1–20. [Google Scholar] [CrossRef]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive Effects of Hypoxia and Temperature on Coastal Pelagic Zooplankton and Fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Breitburg, D.L.; Loher, T.; Pacey, C.A.; Gerstein, A. Varying Effects of Low Dissolved Oxygen on Trophic Interactions in an Estuarine Food Web. Ecol. Monogr. 1997, 67, 489–507. [Google Scholar] [CrossRef]
- Kimmel, D.G.; Boynton, W.R.; Roman, M.R. Long-term decline in the calanoid copepod Acartia tonsa in central Chesapeake Bay, USA: An indirect effect of eutrophication? Estuar. Coast. Shelf Sci. 2012, 101, 76–85. [Google Scholar] [CrossRef]
- Wu, R.S. Hypoxia: From molecular responses to ecosystem responses. Mar. Pollut. Bull. 2002, 45, 35–45. [Google Scholar] [CrossRef]
- Noone, K.J.; Diaz, R.J.; Sumaila, U.R. Managing Ocean. Environments in a Changing Climate: Sustainability and Economic Perspectives, 1st ed.; Elsevier: Burlington, MA, USA, 2013; ISBN 978-0-12-407668-6. [Google Scholar]
- Buchheister, A.; Bonzek, C.F.; Gartland, J.; Latour, R.J. Patterns and drivers of the demersal fish community of Chesapeake Bay. Mar. Ecol. Prog. Ser. 2013, 481, 161–180. [Google Scholar] [CrossRef]
- Lipton, D.; Hicks, R. The cost of stress: Low dissolved oxygen and economic benefits of recreational striped bass (Morone saxatilis) fishing in the Patuxent River. Estuaries 2003, 26, 310–315. [Google Scholar] [CrossRef]
- Diaz, R.J.; Rosenberg, R. Marine benthic hypoxia: A review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanogr. Mar. Biol. Annu. Rev. 1995, 33, 245–303. [Google Scholar]
- Heinle, D.R. Production of a calanoid copepod, Acartia tonsa, in the Patuxent River estuary. Chesap. Sci. 1966, 7, 59–74. [Google Scholar] [CrossRef]
- Lippson, A.J.; Lippson, R.L. Life in the Chesapeake Bay; JHU Press: Baltimore, MD, USA, 2006; ISBN 0-8018-8337-7. [Google Scholar]
- Olson, M.M. Ecological Studies in the Middle Reach of Chesapeake Bay; Springer: Berlin/Heidelberg, Germany, 1987; Volume 23. [Google Scholar]
- Steinberg, D.K.; Condon, R.H. Zooplankton of the York River. J. Coast. Res. 2009, 14, 66–69. [Google Scholar] [CrossRef]
- Wilson, C.B. The copepod crustaceans of Chesapeake Bay. Proc. U. S. Natl. Mus. 1932, 80, 1–54. [Google Scholar] [CrossRef]
- Alekseev, V.R.; Souissi, A. A new species within the Eurytemora affinis complex (Copepoda: Calanoida) from the Atlantic Coast of USA, with observations on eight morphologically different European populations. Zootaxa 2011, 2767, 41–56. [Google Scholar] [CrossRef]
- Kimmel, D.G.; Roman, M.R. Long-term trends in mesozooplankton abundance in Chesapeake Bay, USA: Influence of freshwater input. Mar. Ecol. Prog. Ser. 2004, 267, 71–83. [Google Scholar] [CrossRef]
- White, J.R.; Roman, M.R. Seasonal study of grazing by metazoan zooplankton in the mesohaline Chesapeake Bay. Mar. Ecol. Prog. Ser. 1992, 86, 251–261. [Google Scholar] [CrossRef]
- Bayha, K.M.; Collins, A.G.; Gaffney, P.M. Multigene phylogeny of the scyphozoan jellyfish family Pelagiidae reveals that the common U.S. Atlantic sea nettle comprises two distinct species (Chrysaora quinquecirrha and C. chesapeakei). PeerJ 2017, 5, e3863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrell, V.G.; Van Engel, W.A. Predation by and distribution of a ctenophore, Mnemiopsis leidyi A. Agassiz, in the York River estuary. Estuar. Coast. Mar. Sci. 1976, 4, 235–242. [Google Scholar] [CrossRef]
- Calder, D.R. Hydroids and Hydromedusae of Southern Chesapeake Bay; CAB International: Wallingford, UK, 1971. [Google Scholar]
- Mayer, A.G. Medusae of the World: The Scyphomedusae; Carnegie Institution of Washington: Washington, DC, USA, 1910. [Google Scholar]
- Mayor, A.G. Ctenophores of the Atlantic Coast of North. America; Carnegie Institution of Washington: Washington, DC, USA, 1912. [Google Scholar]
- Morales-Alamo, R.; Haven, D.S. Atypical mouth shape of polyps of the jellyfish, Aurelia aurita, from Chesapeake Bay, Delaware Bay, and Gulf of Mexico. Chesap. Sci. 1974, 15, 22–29. [Google Scholar] [CrossRef]
- Purcell, J.E.; Nemazie, D.A. Quantitative feeding ecology of the hydromedusan Nemopsis bachei in Chesapeake Bay. Mar. Biol. 1992, 113, 305–311. [Google Scholar]
- Stone, S.L. Distribution and Abundance of Fishes and Invertebrates in Mid-Atlantic Estuaries; US Department of Commerce; National Oceanic and Atmospheric Administration: Silver Spring, MD, USA, 1994.
- Wang, S.B.; Houde, E.D. Energy storage and dynamics in bay anchovy Anchoa mitchilli. Mar. Biol. 1994, 121, 219–227. [Google Scholar] [CrossRef]
- Newberger, T.A.; Houde, E.D. Population biology of bay anchovy Anchoa mitchilli in the mid Chesapeake Bay. Mar. Ecol. Prog. Ser. 1995, 116, 25–37. [Google Scholar] [CrossRef]
- Murdy, E.O.; Birdsong, R.S.; Musick, J.A. Fishes of Chesapeake Bay; Smithsonian Institution Press: Washington, DC, USA, 1997; ISBN 1-56098-638-7. [Google Scholar]
- Jung, S.; Houde, E.D. Spatial and temporal variabilities of pelagic fish community structure and distribution in Chesapeake Bay, USA. Estuar. Coast. Shelf Sci. 2003, 58, 335–351. [Google Scholar] [CrossRef]
- Olney, J.E. Eggs and early larvae of the bay anchovy, Anchoa mitchilli, and the weakfish, Cynoscion regalis, in lower Chesapeake Bay with notes on associated ichthyoplankton. Estuaries 1983, 6, 20–35. [Google Scholar] [CrossRef]
- Roman, M.R.; Gauzens, A.L.; Rhinehart, W.K.; White, J.R. Effects of low oxygen waters on Chesapeake Bay zooplankton. Limnol. Oceanogr. 1993, 38, 1603–1614. [Google Scholar] [CrossRef]
- Elliott, D.T.; Pierson, J.J.; Roman, M.R. Copepods and hypoxia in Chesapeake Bay: Abundance, vertical position and non-predatory mortality. J. Plankton Res. 2013, 35, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Ekau, W.; Auel, H.; Poertner, H.; Gilbert, D. Impacts of hypoxia on the structure and processes in pelagic communities (zooplankton, macro-invertebrates and fish). Biogeosciences 2010, 7, 1669–1699. [Google Scholar] [CrossRef] [Green Version]
- Lutz, R.V.; Marcus, N.H.; Chanton, J.P. Effects of low oxygen concentrations on the hatching and viability of eggs of marine calanoid copepods. Mar. Biol. 1992, 114, 241–247. [Google Scholar] [CrossRef]
- Marcus, N.H.; Richmond, C.; Sedlacek, C.; Miller, G.A.; Oppert, C. Impact of hypoxia on the survival, egg production and population dynamics of Acartia tonsa Dana. J. Exp. Mar. Biol. Ecol. 2004, 301, 111–128. [Google Scholar] [CrossRef]
- Richmond, C.; Marcus, N.H.; Sedlacek, C.; Miller, G.A.; Oppert, C. Hypoxia and seasonal temperature: Short-term effects and long-term implications for Acartia tonsa Dana. J. Exp. Mar. Biol. Ecol. 2006, 328, 177–196. [Google Scholar] [CrossRef]
- Vargo, S.L.; Sastry, A.N. Acute temperature and low dissolved oxygen tolerances of brachyuran crab (Cancer irroratus) larvae. Mar. Biol. 1977, 40, 165–171. [Google Scholar] [CrossRef]
- Chesney, E.J.; Houde, E.D. Laboratory Studies on the Effects of Hypoxic Waters on the Survival of Eggs and Yolk-sac Larvae of the Bay Anchovy, Anchoa mitchilli; Center for Environmental Science, Chesapeake Biological Laboratory: Solomons, MD, USA, 1989; pp. 184–191. [Google Scholar]
- Houde, E.D.; Zastrow, C.E. Bay anchovy Anchoa mitchilli. In Habitat Requirements for Chesapeake Bay Living Resources; Chesapeake Bay Program Living Resources Subcommittee: Annapolis, MD, USA, 1991; pp. 8-1–8-14. [Google Scholar]
- MacGregor, J.M.; Houde, E.D. Onshore-offshore pattern and variability in distribution and abundance of bay anchovy Anchoa mitchilli eggs and larvae in Chesapeake Bay. Mar. Ecol. Prog. Ser. 1996, 138, 15–25. [Google Scholar] [CrossRef]
- Jung, S.; Houde, E.D. Recruitment and spawning-stock biomass distribution of bay anchovy (Anchoa mitchilli) in Chesapeake Bay. Fish. Bull. 2004, 102, 63–77. [Google Scholar]
- Taylor, J.C.; Rand, P.S.; Jenkins, J. Swimming behavior of juvenile anchovies (Anchoa spp.) in an episodically hypoxic estuary: Implications for individual energetics and trophic dynamics. Mar. Biol. 2007, 152, 939–957. [Google Scholar] [CrossRef]
- Ludsin, S.A.; Zhang, X.; Brandt, S.B.; Roman, M.R.; Boicourt, W.C.; Mason, D.M.; Costantini, M. Hypoxia-avoidance by planktivorous fish in Chesapeake Bay: Implications for food web interactions and fish recruitment. J. Exp. Mar. Biol. Ecol. 2009, 381, S121–S131. [Google Scholar] [CrossRef]
- Adamack, A.; Rose, K.; Breitburg, D.; Nice, A.; Lung, W. Simulating the effect of hypoxia on bay anchovy egg and larval mortality using coupled watershed, water quality, and individual-based predation models. Mar. Ecol. Prog. Ser. 2012, 445, 141–160. [Google Scholar] [CrossRef] [Green Version]
- North, E.W.; Houde, E.D. Distribution and transport of bay anchovy (Anchoa mitchilli) eggs and larvae in Chesapeake Bay. Estuar. Coast. Shelf Sci. 2004, 60, 409–429. [Google Scholar] [CrossRef]
- Breitburg, D.L. Behavioral response of fish larvae to low dissolved oxygen concentrations in a stratified water column. Mar. Biol. 1994, 120, 615–625. [Google Scholar] [CrossRef]
- Purcell, J.E.; Breitburg, D.L.; Decker, M.B.; Graham, W.M.; Youngbluth, M.J.; Raskoff, K.A. Pelagic cnidarians and ctenophores in low dissolved oxygen environments: A review. In Coastal Hypoxia: Consequences for Living Resources and Ecosystems; American Geophyical Union: Washington, DC, USA, 2001; pp. 77–100. [Google Scholar]
- Breitburg, D.L.; Adamack, A.; Rose, K.A.; Kolesar, S.E.; Decker, B.; Purcell, J.E.; Keister, J.E.; Cowan, J.H. The pattern and influence of low dissolved oxygen in the Patuxent River, a seasonally hypoxic estuary. Estuaries 2003, 26, 280–297. [Google Scholar] [CrossRef]
- Keister, J.E.; Houde, E.D.; Breitburg, D.L. Effects of bottom-layer hypoxia on abundances and depth distributions of organisms in Patuxent River, Chesapeake Bay. Mar. Ecol. Prog. Ser. 2000, 205, 43–59. [Google Scholar] [CrossRef]
- Breitburg, D.L.; Steinberg, N.; DuBean, S.; Cooksey, C.; Houde, E.D. Effects of low dissolved oxygen on predation on estuarine fish larvae. Mar. Ecol. Prog. Ser. 1994, 104, 235–246. [Google Scholar] [CrossRef]
- Decker, M.B.; Breitburg, D.; Purcell, J.E. Effects of low dissolved oxygen on zooplankton predation by the ctenophore Mnemiopsis leidyi. Mar. Ecol. Prog. Ser. 2004, 280, 163–172. [Google Scholar] [CrossRef]
- Shoji, J.; Masuda, R.; Yamashita, Y.; Tanaka, M. Predation on fish larvae by moon jellyfish Aurelia aurita under low dissolved oxygen concentrations. Fish. Sci. 2005, 71, 748–753. [Google Scholar] [CrossRef]
- Shoji, J.; Masuda, R.; Yamashita, Y.; Tanaka, M. Effect of low dissolved oxygen concentrations on behavior and predation rates on red sea bream Pagrus major larvae by the jellyfish Aurelia aurita and by juvenile Spanish mackerel Scomberomorus niphonius. Mar. Biol. 2005, 147, 863–868. [Google Scholar] [CrossRef]
- Pierson, J.; Slater, W.; Elliott, D.; Roman, M. Synergistic effects of seasonal deoxygenation and temperature truncate copepod vertical migration and distribution. Mar. Ecol. Prog. Ser. 2017, 575, 57–68. [Google Scholar] [CrossRef]
- Elliott, D.T.; Pierson, J.J.; Roman, M.R. Predicting the Effects of Coastal Hypoxia on Vital Rates of the Planktonic Copepod Acartia tonsa Dana. PLoS ONE 2013, 8, e63987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.F. The solubility of nitrogen, oxygen and argon in water and seawater. In Proceedings of the Deep Sea Research and Oceanographic Abstracts; Elsevier: Amsterdam, The Netherlands, 1970; Volume 17, pp. 721–735. [Google Scholar]
- McDougall, T.J.; Barker, P.M. Getting started with TEOS-10 and the Gibbs Seawater (GSW) oceanographic toolbox. SCOR/IAPSO WG 2011, 127, 1–28. [Google Scholar]
- Pierson, J.J.; Houde, E.D. Anchovy Data of Chesapeake Bay. Biological and Chemical Oceanography Data Management Office Data Sets. 2017. Available online: https://hdl.handle.net/1912/8925 (accessed on 7 January 2020). [CrossRef]
- Pierson, J.J. Zooplankton from Hypoxic Waters of Chesapeake Bay. Biological and Chemical Oceanography Data Management Office Data Sets. 2017. Available online: https://hdl.handle.net/1912/8934 (accessed on 7 January 2020). [CrossRef]
- Pierson, J.J.; Decker, M.B. DeZoZoo Cruise Data. Biological and Chemical Oceanography Data Management Office Data Sets. 2017. Available online: https://hdl.handle.net/1912/8928 (accessed on 7 January 2020). [CrossRef]
- Barba, A.P. Responses of the Copepod Acartia tonsa to Hypoxia in Chesapeake Bay; University of Maryland: College Park, MD, USA, 2015. [Google Scholar]
- Wiebe, P.H. A multiple opening/closing net and environment sensing system for sampling zooplankton. J. Mar. Res. 1976, 34, 313–326. [Google Scholar]
- Elliott, D.T.; Tang, K.W. Simple staining method for differentiating live and dead marine zooplankton in field samples. Limnol. Oceanogr. Methods 2009, 7, 585–594. [Google Scholar] [CrossRef] [Green Version]
- Elliott, D.T. Copepod Carcasses, Mortality and Population Dynamics in the Tributaries of the Lower Chesapeake Bay; The College of William and Mary: Williamsburg, VA, USA, 2010. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Gray, J.S.; Wu, R.S.; Or, Y.Y. Effects of hypoxia and organic enrichment on the coastal marine environment. Mar. Ecol. Prog. Ser. 2002, 238, 249–279. [Google Scholar] [CrossRef]
- Lange, R.; Staaland, H.; Mostad, A. The effect of salinity and temperature on solubility of oxygen and respiratory rate in oxygen-dependent marine invertebrates. J. Exp. Mar. Biol. Ecol. 1972, 9, 217–229. [Google Scholar] [CrossRef]
- Diaz, R.J. Overview of hypoxia around the world. J. Environ. Qual. 2001, 30, 275–281. [Google Scholar] [CrossRef]
- Thuesen, E.V.; Rutherford, L.D.; Brommer, P.L. The role of aerobic metabolism and intragel oxygen in hypoxia tolerance of three ctenophores: Pleurobrachia bachei, Bolinopsis infundibulum and Mnemiopsis leidyi. J. Mar. Biol. Assoc. U. K. 2005, 85, 627–633. [Google Scholar] [CrossRef] [Green Version]
- Gracey, A.Y.; Troll, J.V.; Somero, G.N. Hypoxia-induced gene expression profiling in the euryoxic fish Gillichthys mirabilis. Proc. Natl. Acad. Sci. USA 2001, 98, 1993–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandic, M.; Todgham, A.E.; Richards, J.G. Mechanisms and evolution of hypoxia tolerance in fish. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 735–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, D.A. Molecular ecology of teleost fish hemoglobins: Strategies for adapting to changing environments. Am. Zool. 1980, 20, 139–162. [Google Scholar] [CrossRef]
- Wood, S.C.; Johansen, K. Adaptation to hypoxia by increased HbO2 affinity and decreased red cell ATP concentration. Nature 1972, 237, 278–279. [Google Scholar] [CrossRef] [PubMed]
- Gilly, W.F.; Beman, J.M.; Litvin, S.Y.; Robison, B.H. Oceanographic and Biological Effects of Shoaling of the Oxygen Minimum Zone. Annu. Rev. Mar. Sci. 2013, 5, 393–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childress, J.J.; Seibel, B.A. Life at stable low oxygen levels: Adaptations of animals to oceanic oxygen minimum layers. J. Exp. Biol. 1998, 201, 1223–1232. [Google Scholar]
- Breitburg, D.L. Episodic hypoxia in Chesapeake Bay: Interacting effects of recruitment, behavior, and physical disturbance. Ecol. Monogr. 1992, 62, 525–546. [Google Scholar] [CrossRef]
- Purcell, J.; Decker, M.; Breitburg, D.; Broughton, K. Fine-scale vertical distributions of Mnemiopsis leidyi ctenophores: Predation on copepods relative to stratification and hypoxia. Mar. Ecol. Prog. Ser. 2014, 500, 103–120. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.R.; Brush, G.S. A 2500-year history of anoxia and eutrophication in Chesapeake Bay. Estuaries 1993, 16, 617–626. [Google Scholar] [CrossRef]
- Palinkas, C.M.; Halka, J.P.; Li, M.; Sanford, L.P.; Cheng, P. Sediment deposition from tropical storms in the upper Chesapeake Bay: Field observations and model simulations. Cont. Shelf Res. 2014, 86, 6–16. [Google Scholar] [CrossRef]
- Hirsch, R.M. Flux of Nitrogen, Phosphorus, and Suspended Sediment from the Susquehanna River Basin to the Chesapeake Bay during Tropical Storm Lee, September 2011, as an Indicator of the Effects of Reservoir Sedimentation on Water Quality; Scientific Investigations Report; United States Department of the Interior: Reston, VA, USA, 2012; p. 28.
- Peierls, B.L.; Christian, R.R.; Paerl, H.W. Water quality and phytoplankton as indicators of hurricane impacts on a large estuarine ecosystem. Estuaries 2003, 26, 1329–1343. [Google Scholar] [CrossRef]
- Stevens, P.W.; Blewett, D.A.; Casey, J.P. Short-term effects of a low dissolved oxygen event on estuarine fish assemblages following the passage of hurricane Charley. Estuaries Coasts 2006, 29, 997–1003. [Google Scholar] [CrossRef]
- Mallin, M.A.; Posey, M.H.; Shank, G.C.; McIver, M.R.; Ensign, S.H.; Alphin, T.D. Hurricane effects on water quality and benthos in the cape fear watershed: Natural and anthropogenic impacts. Ecol. Appl. 1999, 9, 350–362. [Google Scholar] [CrossRef]
- Roman, M.R.; Boicourt, W.C.; Kimmel, D.G.; Miller, W.D.; Adolf, J.E.; Bichy, J.; Harding, L.W.; Houde, E.D.; Jung, S.; Zhang, X. Chesapeake Bay plankton and fish abundance enhanced by Hurricane Isabel. Eos Trans. Am. Geophys. Union 2005, 86, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Cervetto, G.; Gaudy, R.; Pagano, M. Influence of salinity on the distribution of Acartia tonsa (Copepoda, Calanoida). J. Exp. Mar. Biol. Ecol. 1999, 239, 33–45. [Google Scholar] [CrossRef]
- Miller, R.J. Distribution and Biomass of an Estuarine Ctenophore Population, Mnemiopis leidyi (A. Agassiz). Chesap. Sci. 1974, 15, 1–8. [Google Scholar] [CrossRef]
- Purcell, J.E.; White, J.R.; Nemazie, D.A.; Wright, D.A. Temperature, salinity and food effects on asexual reproduction and abundance of the scyphozoan Chrysaora quinquecirrha. Mar. Ecol. Prog. Ser. 1999, 180, 187–196. [Google Scholar] [CrossRef]
- Robinette, H.R. Species Profiles: Life Histories and Environmental Requirements of Coastal Fishes and Invertebrates (Gulf of Mexico): Bay Anchovy and Striped Anchovy; U.S. Army Corps of Engineers Coastal Ecology Group: Washington, DC, USA; Waterways Experiment Station: Vicksburg, MS, USA, 1983. [Google Scholar]
- Bishop, J.W. Ctenophores of the Chesapeake Bay. Chesap. Sci. 1972, 13, S98. [Google Scholar] [CrossRef]
- Farmer, L.; Reeve, M.R. Role of the free amino acid pool of the copepod Acartia tonsa in adjustment to salinity change. Mar. Biol. 1978, 48, 311–316. [Google Scholar] [CrossRef]
- Lance, J. Respiration and osmotic behaviour of the copepod Acartia tonsa in diluted sea water. Comp. Biochem. Physiol. 1965, 14, 155–165. [Google Scholar] [CrossRef]
- Skjoldal, H.R.; Wiebe, P.H.; Postel, L.; Knutsen, T.; Kaartvedt, S.; Sameoto, D.D. Intercomparison of zooplankton (net) sampling systems: Results from the ICES/GLOBEC sea-going workshop. Prog. Oceanogr. 2013, 108, 1–42. [Google Scholar] [CrossRef]
- Breitburg, D.L.; Pihl, L.; Kolesar, S.E. Effects of Low Dissolved Oxygen on the Behavior, Ecology and Harvest of Fishes: A Comparison of the Chesapeake Bay and Baltic-Kattegat Systems. Coast. Estuar. Stud. 2001, 9, 241–267. [Google Scholar]
- Eby, L.A.; Crowder, L.B.; McClellan, C.M.; Peterson, C.H.; Powers, M.J. Habitat degradation from intermittent hypoxia: Impacts on demersal fishes. Mar. Ecol. Prog. Ser. 2005, 291, 249–262. [Google Scholar] [CrossRef]
- Pollock, M.S.; Clarke, L.M.J.; Dubé, M.G. The effects of hypoxia on fishes: From ecological relevance to physiological effects. Environ. Rev. 2007, 15, 1–14. [Google Scholar] [CrossRef]
- Purcell, J.E.; Decker, M.B. Effects of Climate on Relative Predation by Scyphomedusae and Ctenophores on Copepods in Chesapeake Bay during 1987–2000. Limnol. Oceanogr. 2005, 50, 376–387. [Google Scholar] [CrossRef]
- Condon, R.H.; Decker, M.B.; Purcell, J.E. Effects of low dissolved oxygen on survival and asexual reproduction of scyphozoan polyps (Chrysaora quinquecirrha). In Jellyfish Blooms: Ecological and Societal Importance; Springer: Berlin/Heidelberg, Germany, 2001; pp. 89–95. [Google Scholar]
- Breitburg, D.L.; Fulford, R.S. Oyster-sea nettle interdependence and altered control within the Chesapeake Bay ecosystem. Estuaries Coasts 2006, 29, 776–784. [Google Scholar] [CrossRef]
- Grove, M.; Breitburg, D.L. Growth and reproduction of gelatinous zooplankton exposed to low dissolved oxygen. Mar. Ecol. Prog. Ser. 2005, 301, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Newell, R.I. Ecological changes in Chesapeake Bay: Are they the result of overharvesting the American oyster, Crassostrea virginica. Underst. Estuary Adv. Chesap. Bay Res. 1988, 129, 536–546. [Google Scholar]
- McNamara, M.; Lonsdale, D.; Cerrato, R. Role of eutrophication in structuring planktonic communities in the presence of the ctenophore Mnemiopsis leidyi. Mar. Ecol. Prog. Ser. 2014, 510, 151–165. [Google Scholar] [CrossRef]
- Sullivan, B.K.; Van Keuren, D.; Clancy, M. Timing and size of blooms of the ctenophore Mnemiopsis leidyi in relation to temperature in Narragansett Bay, RI. Hydrobiologia 2001, 451, 113–120. [Google Scholar] [CrossRef]
- Oviatt, C.A. The changing ecology of temperate coastal waters during a warming trend. Estuaries 2004, 27, 895–904. [Google Scholar] [CrossRef]
- Purcell, J.E.; Shiganova, T.A.; Decker, M.B.; Houde, E.D. The ctenophore Mnemiopsis in native and exotic habitats: US estuaries versus the Black Sea basin. Hydrobiologia 2001, 451, 145–176. [Google Scholar] [CrossRef]
- McClelland, J.W.; Valiela, I. Changes in food web structure under the influence of increased anthropogenic nitrogen inputs to estuaries. Mar. Ecol. Prog. Ser. 1998, 168, 259–271. [Google Scholar] [CrossRef]
- Breitburg, D. Effects of Hypoxia, and the Balance between Hypoxia and Enrichment, on Coastal Fishes and Fisheries. Estuaries 2002, 25, 767–781. [Google Scholar] [CrossRef]
- Uye, S. Human forcing of the copepod–fish–jellyfish triangular trophic relationship. Hydrobiologia 2011, 666, 71–83. [Google Scholar] [CrossRef]












| Eigenvalue | Difference | Proportion | Cumulative | |
|---|---|---|---|---|
| 1 | 5.01 | 2.68 | 0.56 | 0.56 |
| 2 | 2.33 | 1.68 | 0.26 | 0.82 |
| 3 | 0.66 | 0.16 | 0.07 | 0.89 |
| 4 | 0.50 | 0.21 | 0.06 | 0.95 |
| 5 | 0.30 | 0.21 | 0.03 | 0.98 |
| 6 | 0.08 | 0.04 | 0.01 | 0.99 |
| 7 | 0.05 | 0.01 | 0.01 | 0.99 |
| 8 | 0.04 | 0.02 | 0.00 | 1.00 |
| 9 | - | 0.02 | 0.00 | 1.00 |
| Principal Component 1 | Principal Component 2 | |
|---|---|---|
| DO above pycnocline | 0.35 | 0.20 |
| DO at pycnocline | 0.27 | 0.33 |
| DO below pycnocline | 0.14 | 0.51 |
| Temp. above pycnocline | −0.38 | −0.26 |
| Temp. at pycnocline | −0.39 | −0.12 |
| Temp. below pycnocline | −0.42 | −0.06 |
| Salinity above pycnocline | −0.33 | 0.41 |
| Salinity at pycnocline | −0.33 | 0.39 |
| Salinity below pycnocline | −0.30 | 0.45 |
| LO | MO | |
|---|---|---|
| C | 2011-Spring (N) | 2010-Spring (N, S), 2011-Spring (S) |
| T | 2011-Autumn (N) | 2011-Autumn (S) |
| W | 2011-Summer (N, S), 2010-Summer (N) | 2010-Autumn (N, S), 2010-Summer (S) |
| Zooplankton and Fish Concentration | ||||||
|---|---|---|---|---|---|---|
| Species | Stage | Group | Sample Size | d.f. | Chi-Square | p-Value |
| Acartia tonsa | adult | C | 103 | 1 | 15.1180 | 0.0001 |
| T | 36 | 1 | 9.8108 | 0.0017 | ||
| W | 127 | 1 | 1.6625 | 0.1973 | ||
| Acartia tonsa | copepodite | C | 107 | 1 | 8.5712 | 0.0034 |
| T | 36 | 1 | 8.8460 | 0.0029 | ||
| W | 127 | 1 | 25.785 | <0.0001 | ||
| Anchoa mitchilli | larval | C | 60 | 1 | 6.5997 | 0.0102 |
| T | 24 | 1 | 8.5792 | 0.0034 | ||
| W | 58 | 1 | 0.5652 | 0.4522 | ||
| juvenile | C * | - | - | - | - | |
| T | 24 | 1 | 0.1883 | 0.6643 | ||
| W | 58 | 1 | 4.3211 | 0.0376 | ||
| Mnemiopsis leidyi | adult | C | 119 | 1 | 2.6121 | 0.1061 |
| T | 48 | 1 | 35.225 | <0.0001 | ||
| W | 116 | 1 | 17.545 | <0.0001 | ||
| Chrysaora chesapeakei | medusa | C | 0 | - | - | - |
| T | 48 | 1 | 1 | 0.3171 | ||
| W | 116 | 1 | 2.0241 | 0.1548 | ||
| A. tonsa’s Non-Predatory Mortalities | ||||
|---|---|---|---|---|
| Group | Sample Size | d.f. | Chi-Square | p-Value |
| Cool | 5 | 1 | 0.3333 | 0.5637 |
| Temperate | 6 | 1 | 2.3333 | 0.1266 |
| Warm | 18 | 1 | 6.9286 | 0.008 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
L. Slater, W.; Pierson, J.J.; Decker, M.B.; Houde, E.D.; Lozano, C.; Seuberling, J. Fewer Copepods, Fewer Anchovies, and More Jellyfish: How Does Hypoxia Impact the Chesapeake Bay Zooplankton Community? Diversity 2020, 12, 35. https://doi.org/10.3390/d12010035
L. Slater W, Pierson JJ, Decker MB, Houde ED, Lozano C, Seuberling J. Fewer Copepods, Fewer Anchovies, and More Jellyfish: How Does Hypoxia Impact the Chesapeake Bay Zooplankton Community? Diversity. 2020; 12(1):35. https://doi.org/10.3390/d12010035
Chicago/Turabian StyleL. Slater, Wencheng, James J. Pierson, Mary Beth Decker, Edward D. Houde, Carlos Lozano, and James Seuberling. 2020. "Fewer Copepods, Fewer Anchovies, and More Jellyfish: How Does Hypoxia Impact the Chesapeake Bay Zooplankton Community?" Diversity 12, no. 1: 35. https://doi.org/10.3390/d12010035
APA StyleL. Slater, W., Pierson, J. J., Decker, M. B., Houde, E. D., Lozano, C., & Seuberling, J. (2020). Fewer Copepods, Fewer Anchovies, and More Jellyfish: How Does Hypoxia Impact the Chesapeake Bay Zooplankton Community? Diversity, 12(1), 35. https://doi.org/10.3390/d12010035