
Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review




Abstract
:1. Introduction
2. Materials and Methods
3. Shelter for the Big and for the Small: Floral Chambers in the Cerrado

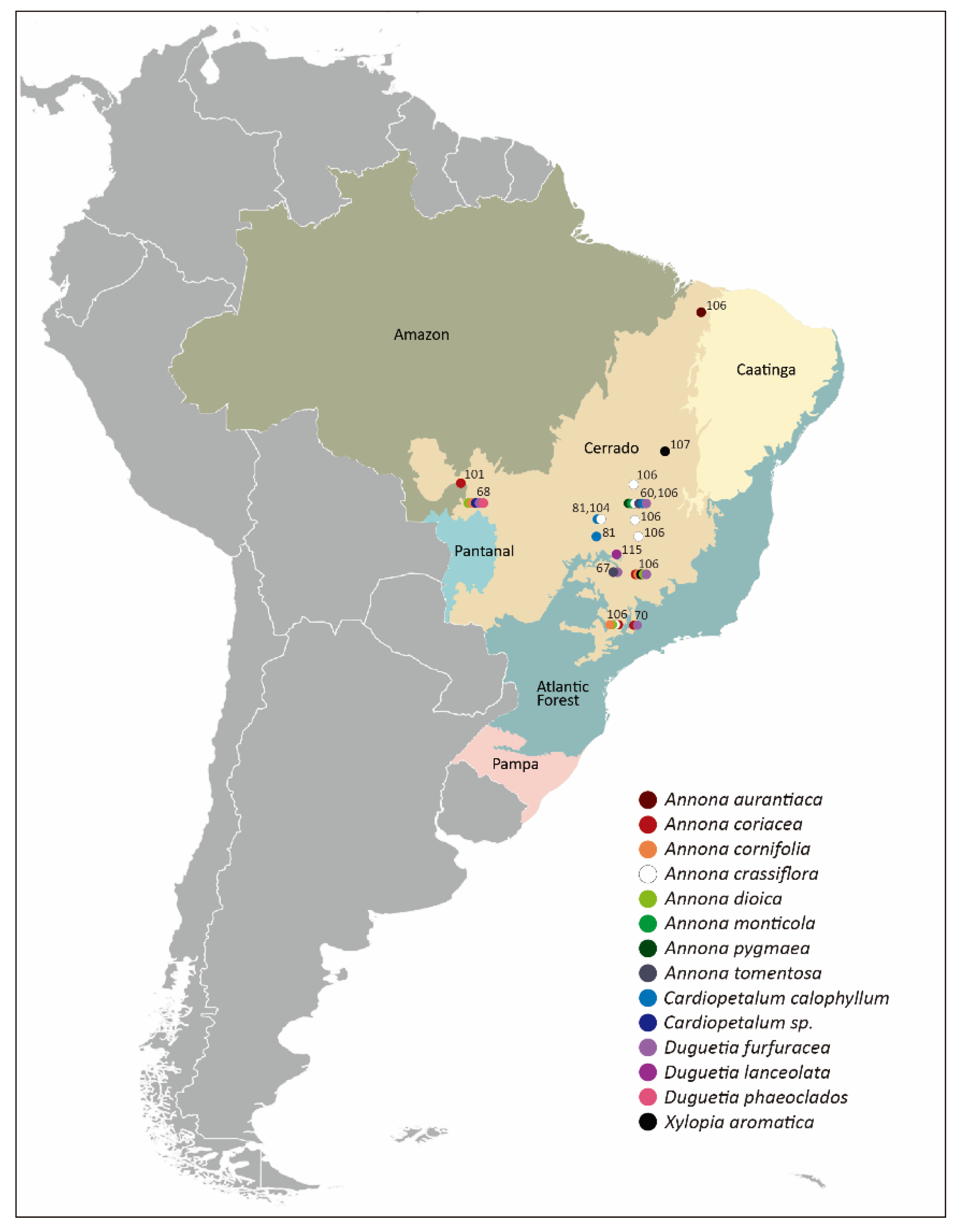{kind=link}
{kind=link}
{kind=link}
| Annonaceae Taxon | Color | Odor | Resources | Time of Anthesis | State | Coleopteran Pollinators | Reference |
|---|---|---|---|---|---|---|---|
| Anaxagorea dolichocarpa 1 | Cream-colored | Banana-like/ acetone-like | Pollen | Diuturnal | Pernambuco | Colopterus sp. a | [88] |
| Annona aurantiaca | Brown/orange yellow | Fruit-like/pungent/disagreeable | Tissue | Nocturnal | Maranhão | Cyclocephala atricapilla b | [106] |
| - | - | - | - | Maranhão | C. atricapilla | [66] | |
| Annona coriacea | Yellow | Pungent | Pollen/ tissue/ exudate/mating site/shelter | Nocturnal | Maranhão | C. atricapilla | [66] |
| Minas Gerais | C. atricapilla | ||||||
| C. quatuordecimpunctata b | |||||||
| São Paulo | C. atricapilla | ||||||
| C. quatuordecimpunctata | |||||||
| C. melanocephala b | |||||||
| C. ohausiana b | |||||||
| Greenish orange/ orange | Sweet | Pollen/ tissue/ mating site/ shelter | Nocturnal | Mato Grosso | C. atricapilla | [101] | |
| C. undata b | |||||||
| C. octopunctata b | |||||||
| C. ohausiana | |||||||
| Arrigutia brevissima b | |||||||
| - | - | Pollen/tissue | - | São Paulo | C. atricapilla | [108] | |
| C. quatuordecimpunctata | |||||||
| Annona cornifolia | Greenish/yellow/creamy | Fruity/pleasant | Tissue/ mating site/shelter | Nocturnal | Minas Gerais | C. quatuordecimpunctata | [66] |
| C. atricapilla | |||||||
| São Paulo | C. quatuordecimpunctata | ||||||
| Yellowish | Present | Pollen/tissue | Nocturnal | Mato Grosso | Coleoptera | [68] | |
| Annona crassiflora | Brownish-yellow | Pungent | Pollen/ tissue/ mating site/shelter | Nocturnal | Distrito Federal | C. atricapilla | [66] |
| Goiás | C. atricapilla | ||||||
| C. quatuordecimpunctata | |||||||
| São Paulo | C. atricapilla | ||||||
| C. literata b | |||||||
| Minas Gerais | C. atricapilla | ||||||
| - | - | - | - | Minas Gerais | C. atricapilla | [104] | |
| C. latericia b | |||||||
| C. octopunctata | |||||||
| Reddish-brown | - | Pollen/petals | - | Distrito Federal | Cyclocephala cf. atricapilla | [60] | |
| Annona dioica | Yellow | Pungent/unpleasant | Pollen/tissue | Nocturnal | Minas Gerais | C. atricapilla | [66] |
| C. quatuordecimpunctata | |||||||
| São Paulo | C. atricapilla | ||||||
| Yellow | Present | Pollen/tissue | Nocturnal | Mato Grosso | Coleoptera | [68] | |
| Annona monticola | - | Unpleasant | - | - | Distrito Federal | C. atricapilla | [66] |
| Annona tomentosa | Yellowish | Fruit-like/ agreeable | - | - | Distrito Federal | C. atricapilla | [106] |
| C. quatuordecimpunctata | |||||||
| Minas Gerais | C. quatuordecimpunctata | ||||||
| C. atricapilla | |||||||
| Creamy | Present | Pollen | Nocturnal | Minas Gerais | Coleoptera | [67] | |
| Annona pygmaea [syn. A. warmingiana] | - | Fruit-like/ unpleasant | - | - | Distrito Federal | C. atricapilla | [106] |
| Cardiopetalum sp. | Yellow | Present | Pollen/tissue | Nocturnal | Mato Grosso | Coleoptera | [68] |
| Cardiopetalum calophyllum | - | Decaying fruit | - | Diurnal | Distrito Federal | Lobiopa spp. a | [66] |
| - | - | - | - | Goiás | Lobiopa insularis a | [81] | |
| Greenish/white | Present | - | Nocturnal | Minas Gerais | Chrysomelidae | [109] | |
| Coccinelidae | |||||||
| Duguetia furfuracea | Red | Fruity/ decaying fruit | - | Diurnal | Distrito Federal | Nitidulidae spp. | [66] |
| Curculionidae aff. Prantisus sp. | |||||||
| Minas Gerais | Lobiopa sp. | ||||||
| Curculionidae aff. Prantisus sp. | |||||||
| Dark red/purple | Mature fruit | - | Diuturnal | São Paulo | Colopterus sp. | [70] | |
| Red | Present | Pollen/tissue | Diuturnal | Mato Grosso | Coleoptera | [68] | |
| Rosy | Undetected | Pollen | Nocturnal | Minas Gerais | Coleoptera | [67] | |
| Duguetia lanceolata | Purple/reddish-orange | Characteristic | - | Diuturnal | Minas Gerais | Chrysomelidae | [109] |
| Coccinelidae | |||||||
| Duguetia phaeoclados [syn. Annona phaeoclados] | Yellow | Present | Pollen/ tissue | Nocturnal | Mato Grosso | Coleoptera | [68] |
| Xylopia aromatica | White | Very sweet | Pollen/tissue | Diurnal | Amazonas | Chrysomelidae sp. | [66] |
| Nitidulidae spp. | |||||||
| Minas Gerais | Thysanoptera spp. | ||||||
| Chrysomelidae sp. | |||||||
| Cillaeus sp. a | |||||||
| Carpophilinae sp. a | |||||||
| Mato Grosso | Cillaeus sp. | ||||||
| São Paulo | Halticinae sp. c | ||||||
| Cillaeus sp. | |||||||
| - | Undetected | - | Diuturnal | Bahia | Curculionidae sp. | [107] |
4. Beetle Marks and Other Mysteries
5. Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Labandeira, C.C. Insect mouthparts: Ascertaining the paleobiology of insect feeding strategies. Annu. Rev. Ecol. Syst. 1997, 28, 153–193. [Google Scholar] [CrossRef] [Green Version]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; St. John, O.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S.; et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Nishikawa, M.; Sota, T. Loss of flight promotes beetle diversification. Nat. Commun. 2012, 3, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, D.D.; Wild, A.L.; Kanda, K.; Bellamy, C.L.; Beutel, R.G.; Caterino, M.S.; Farnum, C.W.; Hawks, D.C.; Ivie, M.A.; Jameson, M.L.; et al. The beetle tree of life reveals that Coleoptera survived end-Permian mass extinction to diversify during the Cretaceous terrestrial revolution. Syst. Entomol. 2015, 40, 835–880. [Google Scholar] [CrossRef] [Green Version]
- Labandeira, C.C. The role of insects in Late Jurassic to Middle Cretaceous Ecosystems. In Lower and Middle Cretaceous Terrestrial Ecosystems; Lucas, S.G., Kirkland, J.I., Estep, J.W., Eds.; New Mexico Museum of Natural History and Science: Albuquerque, NM, USA, 1998; pp. 105–124. [Google Scholar]
- Wang, B.; Zhang, H.; Jarzembowski, E.A. Early Cretaceous angiosperms and beetle evolution. Front. Plant Sci. 2013, 4, 360. [Google Scholar] [CrossRef] [Green Version]
- Saunders, R.M.K. The evolution of key functional floral traits in the early divergent angiosperm family Annonaceae. J. Syst. Evol. 2020, 58, 369–392. [Google Scholar] [CrossRef]
- Gottsberger, G. Comments on flower evolution and beetle pollination in the genera Annona and Rollinia. Plant Syst. Evol. 1989, 167, 189–194. [Google Scholar] [CrossRef]
- Souza, V.C.; Lorenzi, H. Botânica Sistemática: Guia Ilustrada para Identificação das Famílias Nativas e Exóticas no Brasil, Baseado em APG III; Instituto Plantarum: Nova Odessa, Brazil, 2012. [Google Scholar]
- Teixeira, S.P.; Marinho, C.R.; Paulino, J.V. A flor: Aspectos morfofuncionais e evolutivos. In Biologia da Polinização; Rech, A.R., Agostini, K., Oliveira, P.E., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; pp. 45–69. [Google Scholar]
- Gottsberger, G. The reproductive biology of the early-divergent genus Anaxagorea (Annonaceae), and its significance for the evolutionary development of the family. Acta Bot. Bras. 2016, 30, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Couvreur, T.L.P.; Helmstetter, A.J.; Koenen, E.J.M.; Bethune, K.; Brandão, R.D.; Little, S.A.; Sauquet, H.; Erkens, R.H.J. Phylogenomics of the major tropical plant family Annonaceae using targeted enrichment of nuclear genes. Front. Plant Sci. 2019, 9, 1491. [Google Scholar] [CrossRef] [Green Version]
- Xue, B.; Guo, X.; Landis, J.B.; Sun, M.; Tang, C.C.; Soltis, P.S.; Soltis, D.E.; Saunders, R.M.K. Accelerated diversification correlated with functional traits shapes extant diversity of the early-divergent angiosperm family Annonaceae. Mol. Phylogenet. Evol. 2020, 142, 106659. [Google Scholar] [CrossRef]
- Gottsberger, G. Pollination and evolution in Neotropical Annonaceae. Plant Species Biol. 1999, 14, 143–152. [Google Scholar] [CrossRef]
- Paulino-Neto, H.F. Polinização por besouros. In Biologia da Polinização; Rech, A.R., Agostini, K., Oliveira, P.E., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; pp. 259–276. [Google Scholar]
- Gottsberger, G. Flowers and beetles in the South American Tropics. Bot. Acta 1990, 103, 360–365. [Google Scholar] [CrossRef]
- Gibernau, M.; Barabé, D.; Cerdan, P.; Dejean, A. Beetle pollination of Philodendron solimoesense (Araceae) in French Guiana. Int. J. Plant Sci. 1999, 160, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, A.C.D.; Gibernau, M.; Dötterl, S.; Navarro, D.M.A.F.; Seifert, K.; Müller, T.; Schlindwein, C. The floral scent of Taccarum ulei (Araceae): Attraction of scarab beetle pollinators to an unusual aliphatic acyloin. Phytochemistry 2013, 93, 71–78. [Google Scholar] [CrossRef]
- Pereira, J.; Schlindwein, C.; Antonini, Y.; Maia, A.C.D.; Dötterl, S.; Martins, C.; Navarro, D.M.A.F.; Oliveira, R. Philodendron adamantinum (Araceae) lures its single cyclocephaline scarab pollinator with specific dominant floral scent volatiles. Biol. J. Linn. Soc. Lond. 2014, 111, 679–691. [Google Scholar] [CrossRef] [Green Version]
- Milet-Pinheiro, P.; Gonçalves, E.G.; Navarro, D.M.A.F.; Nuñez-Avellaneda, L.A.; Maia, A.C.D. Floral scent chemistry and pollination in the Neotropical aroid genus Xanthosoma (Araceae). Flora 2017, 231, 1–10. [Google Scholar] [CrossRef]
- Silberbauer-Gottsberger, I. Pollination and evolution in palms. Phyton 1990, 30, 213–233. [Google Scholar]
- Küchmeister, H.; Webber, A.C.; Silberbauer-Gottsberger, I.; Gottsberger, G. A polinização e sua relação com a termogênese em espécies de Arecaceae e Annonaceae da Amazônia Central. Acta Amazon. 1998, 28, 217–245. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.S.P.; Couturier, G.; Beserra, P. Biologia da polinização da palmeira tucumã (Astrocaryum vulgare Mart.) em Belém, Pará, Brasil. Acta Bot. Bras. 2003, 17, 343–353. [Google Scholar] [CrossRef] [Green Version]
- De Medeiros, B.A.S.; Núñez-Avellaneda, L.A.; Hernandez, A.M.; Farrell, B.D. Flower visitors of the licuri palm (Syagrus coronata): Brood pollinators coexist with a diverse community of antagonists and mutualists. Biol. J. Linn. Soc. Lond. 2019, 126, 666–687. [Google Scholar] [CrossRef]
- Eriksson, R. The remarkable weevil pollination of the Neotropical Carludovicoideae (Cyclanthaceae). Plant Syst. Evol. 1994, 189, 75–81. [Google Scholar] [CrossRef]
- Franz, N.M.; Valente, R.M. Evolutionary trends in derelomine flower weevils (Coleoptera: Curculionidae): From associations to homology. Invertebr. Syst. 2005, 19, 499–530. [Google Scholar] [CrossRef]
- Moore, M.R.; Jameson, M.L. Floral associations of Cyclocephaline scarab beetles. J. Insect Sci. 2013, 13, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valente, R.M.; Da Silva, P.A.L.; De Medeiros, B.A.S. The first species of Cotithene Voss (Coleoptera: Curculionidae: Curculioninae) from Amazonian Brazil, with notes on its role as a pollinator of Evodianthus funifer (Poit.) Lindm. (Cyclanthaceae). Zootaxa 2019, 4576, 461–482. [Google Scholar] [CrossRef] [PubMed]
- Dieringer, G.; Espinosa, J.E. Reproductive ecology of Magnolia schiedeana (Magnoliaceae), a threatened cloud forest tree species in Veracruz, Mexico. Bull. Torrey Bot. Club. 1994, 121, 154–159. [Google Scholar] [CrossRef]
- Thien, L.B.; Kawano, S.; Devall, M.; Latimer, S.; Rosso, S.; Azuma, H.; Jobes, D.V. Fluorescent Magnolia flowers. Plant Species Biol. 1995, 10, 61–64. [Google Scholar] [CrossRef]
- Dieringer, G.; Cabrera, R.L.; Lara, M.; Loya, L.; Reyes-Castillo, P. Beetle pollination and floral thermogenicity in Magnolia tamaulipana (Magnoliaceae). Int. J. Plant Sci. 1999, 160, 64–71. [Google Scholar]
- Gottsberger, G.; Silberbauer-Gottsberger, I.; Dötterl, S. Pollination ecology of Magnolia ovata may explain the overall large flower size of the genus. Flora 2012, 207, 107–118. [Google Scholar] [CrossRef]
- Singer, R.B.; Cocucci, A.A. Pollination of Pteroglossaspis ruwenzoriensis (Rendle) Rolfe (Orchidaceae) by beetles in Argentina. Bot. Acta 1997, 110, 338–342. [Google Scholar] [CrossRef]
- Steiner, K.E. The evolution of beetle pollination in a South African Orchid. Am. J. Bot. 1998, 85, 1180–1193. [Google Scholar]
- Peter, C.I.; Johnson, S.D. Pollination by flower chafer beetles in Eulophia ensata and Eulophia welwitschii (Orchidaceae). S. Afr. J. Bot. 2009, 75, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Tang, W. Insect pollination in the cycad Zamia pumila (Zamiaceae). Am. J. Bot. 1987, 74, 90–99. [Google Scholar] [CrossRef]
- Norstog, K.J.; Fawcett, P.K.S.; Vovides, A.P. Beetle pollination of two species of Zamia: Evolutionary and ecological considerations. Palaeobotanist 1992, 41, 149–158. [Google Scholar]
- Sakai, S. A review of brood-site pollination mutualism: Plants providing breeding sites for their pollinators. J. Plant Res. 2002, 115, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Tobe, H. Is Cycas revoluta (Cycadaceae) wind- or insect-pollinated? Am. J. Bot. 2007, 94, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Cousins, S.R.; Witkowski, E.T.F. African cycad ecology, ethnobotany and conservation: A synthesis. Bot. Rev. 2017, 83, 152–194. [Google Scholar] [CrossRef]
- Rech, A.R.; Bergamo, P.J.; Figueiredo, R.A. Polinização abiótica. In Biologia da Polinização; Rech, A.R., Agostini, K., Oliveira, P.E., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; pp. 183–204. [Google Scholar]
- Henderson, A. A review of pollination studies in the Palmae. Bot. Rev. 1986, 52, 221–259. [Google Scholar] [CrossRef]
- Hu, S.; Dilcher, D.L.; Taylor, D.W. Pollen evidence for the pollination biology of early flowering plants. In Evolution of Plant–Pollinator Relationships; Patiny, S., Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 165–236. [Google Scholar]
- Li, H.T.; Yi, T.S.; Gao, L.M.; Ma, P.F.; Zhang, T.; Yang, J.B.; Gitzendanner, M.A.; Fritsch, P.W.; Cai, J.; Luo, Y.; et al. Origin of angiosperms and the puzzle of the Jurassic gap. Nat. Plants 2019, 5, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooi, C.J.; Ollerton, J. The origins of flowering plants and pollinators. Science 2020, 368, 1306–1308. [Google Scholar]
- Thien, L.B.; Bernhardt, P.; Devall, M.S.; Chen, Z.D.; Luo, Y.B.; Fan, J.H.; Yuan, L.C.; Williams, J.H. Pollination biology of basal angiosperms (ANITA Grade). Am. J. Bot. 2009, 96, 166–182. [Google Scholar] [CrossRef]
- Labandeira, C.C. Fossil history and evolutionary ecology of Diptera and their associations with plants. In The Evolutionary Biology of Flies; Yeats, D.K., Wiegmann, D.M., Eds.; Columbia University Press: New York, NY, USA, 2005; pp. 217–273. [Google Scholar]
- Paccini, E.; Hesse, M. Pollenkitt—its composition, forms and functions. Flora 2005, 200, 399–415. [Google Scholar] [CrossRef]
- Gandolfo, M.A.; Nixon, K.C.; Crepet, W.L. Cretaceous flowers of Nymphaeaceae and implications for complex insect entrapment pollination mechanisms in early angiosperms. Proc. Natl. Acad. Sci. USA 2004, 101, 8056–8060. [Google Scholar] [CrossRef] [Green Version]
- Ervik, F.; Knudsen, J.T. Water lilies and scarabs: Faithful partners for 100 million years? Biol. J. Linn. Soc. Lond. 2003, 80, 539–543. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, P. Convergent evolution and adaptive radiation of beetle-pollinated angiosperms. Plant Syst. Evol. 2000, 222, 293–320. [Google Scholar] [CrossRef]
- Friedman, J.; Barrett, S.P.C. A phylogenetic analyses of the evolution of wind pollination in the angiosperms. Int. J. Plant Sci. 2008, 169, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Webber, A.C.; Gottsberger, G. Floral biology and pollination of Bocageopsis multiflora and Oxandra euneura in Central Amazonia, with remarks on the evolution of stamens in Annonaceae. Feddes Repert. 1995, 106, 515–524. [Google Scholar] [CrossRef]
- Momose, K.; Nagamitsu, T.; Inoue, T. Thrips cross-pollination of Popowia pisocarpa (Annonaceae) in a lowland dipterocarp forest in Sarawak. Biotropica 1998, 30, 444–448. [Google Scholar] [CrossRef]
- Silberbauer-Gottsberger, I.; Gottsberger, G.; Webber, A.C. Morphological and functional flower characteristics of New and Old World Annonaceae with respect to their mode of pollination. Taxon 2003, 52, 701–718. [Google Scholar] [CrossRef]
- Gottsberger, G.; Meinke, S.; Porembski, S. First records of flower biology and pollination in African Annonaceae: Isolona, Piptostigma, Uvariodendron, Monodora and Uvariopsis. Flora 2011, 206, 498–510. [Google Scholar] [CrossRef]
- Carvalho, R.; Webber, A.C. Biologia floral de Unonopsis guatterioides (A.D.C.) R.E. Fr., uma Annonaceae polinizada por Euglossini. Rev. Bras. Bot. 2000, 23, 421–425. [Google Scholar] [CrossRef]
- Teichert, H.; Dötterl, S.; Zimma, B.; Ayasse, M.; Gottsberger, G. Perfume-collecting male euglossine bees as pollinators of a basal angiosperm—the case of Unonopsis stipitata. Plant Biol. 2008, 11, 29–37. [Google Scholar] [CrossRef]
- Nagamitsu, T.; Inoue, T. Cockroach pollination and breeding system of Uvaria elmeri (Annonaceae) in a lowland mixed-dipterocarp forest in Sarawak. Am. J. Bot. 1997, 84, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, P.E.; Gibbs, P.E. Reproductive biology of woody plants in a cerrado community of Central Brazil. Flora 2000, 195, 311–329. [Google Scholar] [CrossRef]
- Caetano, A.P.S.; Cortez, P.A. Reprodução assexuada. In Biologia da Polinização; Rech, A.R., Agostini, K., Oliveira, P.E., Machado, I.C., Eds.; Projeto Cultural: Rio de Janeiro, Brazil, 2014; pp. 93–111. [Google Scholar]
- Alves, R.M.; Artero, A.S.; Sebbenn, A.M.; Figueira, A. Mating system in a natural population of Theobroma grandiflorum (Willd. ex Spreng.) Schum., by microsatellite markers. Genet. Mol. Biol. 2003, 26, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Rech, A.R. Reproductive biology of Davilla kunthii A. St-Hil. (Dilleniaceae) in Central Amazonia. Acta Bot. Bras. 2011, 25, 487–496. [Google Scholar] [CrossRef]
- Saraiva, L.C.; Cesar, O.; Monteiro, R. Breeding systems of shrubs and trees of a Brazilian savanna. Arq. Biol. Tecnol. 1996, 39, 751–763. [Google Scholar]
- Ishara, K.L.; Maimoni-Rodella, C.S. Pollination and dispersal systems in a Cerrado remnant (Brazilian Savanna) in Southeastern Brazil. Braz. Arch. Biol. Technol. 2011, 54, 629–642. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G.; Silberbauer-Gottsberger, I. Pollination and floral biology in one hectare of Cerrado. In Life in the Cerrado: A South American Tropical Seasonal Vegetation; Reta Verlag: Ulm, Germany, 2006; Volume 2. [Google Scholar]
- Barbosa, A.A.A. Biologia Reprodutiva de Uma Comunidade de Campo Sujo, Uberlândia/MG. Ph.D. Thesis, Universidade Estadual de Campinas, Campinas, Brazil, 1997. [Google Scholar]
- Borges, H.B.N. Biologia Reprodutiva e Onservação do Estrato Lenhoso Numa Comunidade do Cerrado. Ph.D. Thesis, Universidade Estadual de Campinas, Campinas, Brazil, 2000. [Google Scholar]
- Martins, F.Q.; Batalha, M.A. Pollination systems and floral traits in Cerrado woody species of the Upper Taquari Region (Central Brazil). Braz. J. Biol. 2006, 66, 543–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulino-Neto, H.F. História Natural e Interação Flores-Besouros em Espécies de Cerrado. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2009. [Google Scholar]
- Batalha, M.A.; Mantovani, W. Reproductive phenological patterns of Cerrado plant species at the Pé-de-Gigante Reserve (Santa Rita do Passa Quatro, SP, Brasil): A comparison between the herbaceous and woody floras. Braz. J. Biol. 2000, 60, 129–145. [Google Scholar]
- Gottsberger, G. As anonáceas do cerrado e a sua polinização. Braz. J. Biol. 1994, 54, 391–402. [Google Scholar]
- Riehs, P.J. Fenologia de algumas espécies do Gênero Cyclocephala (Coleoptera, Scarabaeidae) do este e centro-oeste do Paraná, Brasil. Rev. Ciênc. Exatas Nat. 2006, 8, 201–223. [Google Scholar]
- Silva, N.A.P.; Frizzas, M.A.; Oliveira, C.M. Seasonality in insect abundance in the Cerrado of Goiás State, Brazil. Rev. Bras. Entomol. 2011, 55, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Hausmann, C.; Samietz, J.; Dorn, S. Visual orientation of overwintered Anthonomus pomorum (Coleoptera: Curculionidae). Environ. Entomol. 2004, 33, 1410–1415. [Google Scholar] [CrossRef] [Green Version]
- Döring, D.F.; Skellern, M.; Watts, N.; Cook, S.M. Colour choice behaviour in the pollen beetle Meligethes aeneus (Coleoptera: Nitidulidae). Physiol. Entomol. 2012, 37, 360–368. [Google Scholar] [CrossRef]
- Sharkey, C.R.; Fujimoto, M.S.; Lord, N.P.; Shin, S.; McKenna, D.; Suvorov, A.; Martin, G.J.; Bybee, S.M. Overcoming the loss of blue sensitivity through opsin duplication in the largest animal group, beetles. Sci. Rep. 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Harms, J.; Vorobyev, M.; Schron, J.; Shmida, A.; Keasar, T.; Homberg, U.; Schmeling, F.; Menzel, R. Evidence of red sensitive photoreceptors in Pygopleurus israelitus (Glaphyridae: Coleoptera) and its implications for beetle pollination in the southeast Mediterranean. J. Comp. Physiol. A 2012, 198, 451–463. [Google Scholar] [CrossRef]
- Warrant, E.; Dacke, M. Vision and visual navigation in nocturnal insects. Annu. Rev. Entomol. 2011, 56, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Teichert, H.; Dötterl, S.; Frame, D.; Kirejtshuk, A.; Gottsberger, G. A novel pollination mode, saprocantharophily, in Duguetia cadaverica (Annonaceae): A stinkhorn (Phallales) flower mimic. Flora 2012, 207, 522–529. [Google Scholar] [CrossRef]
- Elias, M.A.S.; Franceschinelli, E.V.; Juen, L.; Borges, F.J.A.; Ferreira, G.M.; Carvalho, F.M.V. Reproductive success of Cardiopetalum calophyllum (Annonaceae) treelets in fragments of Brazilian savanna. J. Trop. Ecol. 2012, 28, 317–320. [Google Scholar] [CrossRef]
- Cline, A.R.; Kinnee, S.A. A new species of sap beetle (Coleoptera: Nitidulidae) from Baja California Sur, Mexico, with a review of the genus Lobiopa Erichson. Pan-Pac. Entomol. 2012, 88, 202–211. [Google Scholar] [CrossRef]
- Jürgens, A.; Webber, A.C.; Gottsberger, G. Floral scent compounds of Amazonian Annonaceae species pollinated by small beetles and thrips. Phytochemistry 2000, 55, 551–558. [Google Scholar] [CrossRef]
- Maia, A.C.D.; Schlindwein, C.; Navarro, D.M.A.F.; Gibernau, M. Pollination of Philodendron acutatum (Araceae) in the Atlantic Forest of northeastern Brazil: A single scarab beetle species guarantees high fruit set. Int. J. Plant Sci. 2010, 171, 740–748. [Google Scholar] [CrossRef]
- Goodrich, K.R. Floral scent in Annonaceae. Bot. J. Linn. Soc. 2012, 169, 262–279. [Google Scholar] [CrossRef] [Green Version]
- Tsukada, M.; Inui, M.; Suzaki, N. Do beetles prefer the odor of female-stage to male-stage flowers in Atemoya, a cantharophylous protogynous fruit tree (Annonaceae)? J. Entomol. Res. Soc. 2017, 19, 43–52. [Google Scholar]
- Schatz, G.E. Some aspects of pollination biology in Central American forests. In Reproductive Ecology of Tropical Forest Plants; Bawa, K.S., Hadley, M., Eds.; UNESCO and The Parthenon Publishing Group: Paris, France, 1990; pp. 69–84. [Google Scholar]
- Braun, M.; Gottsberger, G. Floral biology and breeding system of Anaxagorea dolichocarpa (Annonaceae), with observations on the interval between anthesis and fruit formation. Phyton 2011, 51, 315–327. [Google Scholar]
- Habeck, D.H. Nitidulidae Latreille 1802. In American Beetles; Arnet, R.H., Thomas, M.C., Skelley, P.E., Frank, J.H., Eds.; CRC Press: Boca Ratón, FL, USA, 2002; Volume 2, pp. 311–315. [Google Scholar]
- Almeida, E.B., Jr.; Collevatti, R.G.; Telles, M.P.C.; Chaves, L.J.; Neres, D.F.; Soares, T.N. Short-distance pollen dispersal in a protogynous Annonaceae tree species from the Brazilian Cerrado. Plant Syst. Evol. 2018, 304, 1091–1099. [Google Scholar]
- Gibernau, M.; Barabé, D.; Labat, D. Flowering and pollination of Philodendron melinonii (Araceae) in French Guiana. Plant Biol. 2000, 2, 331–334. [Google Scholar] [CrossRef]
- Gottsberger, G. Pollination of some species of Carludovicoideae and remarks on the origin and evolution of Cyclanthaceae. Bot. Jahrb. Syst. 1991, 113, 221–235. [Google Scholar]
- Meeuse, B.J.D.; Raskin, I. Sexual reproduction in the arum lily family, with emphasis on thermogenicity. Sex. Plant Reprod. 1988, 1, 3–15. [Google Scholar] [CrossRef]
- Seymour, R.S.; Schultze-Motel, P. Heat-producing flowers. Endeavour 1997, 21, 125–129. [Google Scholar] [CrossRef]
- Dieringer, G.; Reyes-Castillo, P.; Lara, M.; Cabrera, R.L.; Loya, L. Endothermy and floral utilization of Cyclocephala caelestis (Coleoptera: Scarabaeoidea; Melolonthidae); a cloud forest endemic beetle. Acta Zool. Mex. 1998, 73, 145–153. [Google Scholar]
- Seymour, R.S.; Schultze-Motel, P. Physiological temperature regulation by flowers of the sacred lotus. Philos. Trans. R. Soc. 1998, 353, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Seymour, R.S.; White, C.R.; Gibernau, M. Heat reward for insect pollinators. Nature 2003, 426, 243–244. [Google Scholar] [CrossRef]
- Rands, S.A.; Whitney, H.M. Floral temperature and optimal foraging: Is heat a feasible floral reward for pollinators? PLoS ONE 2008, 3, e2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, R.S.; White, C.R.; Gibernau, M. Endothermy of dynastine scarab beetles (Cyclocephala colasi) associated with pollination biology of a thermogenic arum lily (Philodendron solimoesense). J. Exp. Biol. 2009, 212, 2960–2968. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.A.; Domingues-Neta, A.M. Aspectos reprodutivos e visitantes florais de Duguetia marcgraviana Mart. (Annonaceae) na região sudoeste de Mato Grosso. Biotemas 2010, 23, 69–76. [Google Scholar]
- Costa, M.S.; Silva, R.J.; Paulino-Neto, H.F.; Ferreira, M.J.B. Beetle pollination and flowering rhythm of Annona coriacea Mart. (Annonaceae) in Brazilian Cerrado: Behavioral features of its principal pollinators. PLoS ONE 2017, 12, e0171092. [Google Scholar]
- Silva-Júnior, M.C. 100 Árvores do Cerrado—Guia de Campo; Rede de sementes do Cerrado: Brasília, Brazil, 2005. [Google Scholar]
- Lorenzi, H. Árvores Brasileiras: Manual de Identificação e Cultivo de Plantas Arbóreas Nativas do Brasil; Instituto Plantarum: Nova Odessa, Brazil, 2009; Volume 1. [Google Scholar]
- Cavalcante, T.R.M.; Naves, R.V.; Franceschinelli, R.V.; da Silva, R.P. Polinização e formação de frutos em araticum. Bragantia 2009, 68, 13–21. [Google Scholar] [CrossRef]
- Gottsberger, G.; Webber, A.C. Nutritious tissue in petals of Annonaceae and its function in pollination by scarab beetles. Acta Bot. Bras. 2018, 32, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Gottsberger, G. Beetle pollination and flowering rhythm of Annona spp. (Annonaceae) in Brazil. Plant Syst. Evol. 1989, 167, 165–187. [Google Scholar] [CrossRef]
- Aragão, D.S.; Costa, C.B.N.; Nascimento, V.T. Biologia floral, fenologia reprodutiva e polinização de Xylopia aromatica (Lam.) Mart. (Annonaceae) em uma área de Cerrado no oeste da Bahia. Paubrasilia 2019, 2, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Paulino-Neto, H.F. Polinização e biologia reprodutiva do araticum-liso (Annona coriacea Mart.: Annonaceae) em uma área de Cerrado paulista: Implicações para fruticultura. Rev. Bras. Frutic. 2014, 36, 132–140. [Google Scholar] [CrossRef] [Green Version]
- Paulino-Neto, H.F. Biologia de Reprodução de Annonaceae em Matas do Brasil Central. Undergraduate Thesis, Universidade Federal de Uberlândia, Uberlândia, Brazil, 1999. [Google Scholar]
- van Kleunen, M.; Nänni, I.; Donaldson, J.S.; Manning, J.C. The role of beetle marks and flower colour on visitation by Monkey Beetles (Hopliini) in the Greater Cape Floral Region, South Africa. Ann. Bot. 2007, 100, 1483–1489. [Google Scholar] [CrossRef] [Green Version]
- Aquino, F.G.; Miranda, G.H.B. Consequências ambientais da fragmentação de habitats no Cerrado. In Cerrado—Ecologia e Flora; Sano, S.M., Almeida, S.P., Ribeiro, J.F., Eds.; Embrapa Informação Tecnológica: Brasília, Brazil, 2008; Volume 1, pp. 383–398. [Google Scholar]
- Moore, M.R.; Cave, R.D.; Branham, M.A. Synopsis of the cyclocephaline scarab beetles (Coleoptera, Scarabaeidae, Dynastinae). Zookeys 2018, 745, 1–99. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.C.R.; Pinto, A.C.Q.; Ramos, V.H.V. O Cultivo da Pinha, Fruta-do-Conde ou Ata no Brasil; Embrapa Cerrados: Planaltina, Brazil, 2000. [Google Scholar]
- Lemos, E.E.P. A Produção de Anonáceas no Brasil. Rev. Bras. Frutic. 2014, 36, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Parizotto, D.R.; Grossi, P.C. Revisiting pollinating Cyclocephala scarab beetles (Coleoptera: Melolonthidae: Dynastinae) associated with the soursop (Annona muricata, Annonaceae). Neotrop. Entomol. 2019, 48, 415–421. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saravy, F.P.; Marques, M.I.; Schuchmann, K.-L. Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review. Diversity 2021, 13, 438. https://doi.org/10.3390/d13090438
Saravy FP, Marques MI, Schuchmann K-L. Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review. Diversity. 2021; 13(9):438. https://doi.org/10.3390/d13090438
Chicago/Turabian StyleSaravy, Fábio Pinheiro, Marinez Isaac Marques, and Karl-L. Schuchmann. 2021. "Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review" Diversity 13, no. 9: 438. https://doi.org/10.3390/d13090438
APA StyleSaravy, F. P., Marques, M. I., & Schuchmann, K. -L. (2021). Coleopteran Pollinators of Annonaceae in the Brazilian Cerrado—A Review. Diversity, 13(9), 438. https://doi.org/10.3390/d13090438