
A Long-Term Symbiotic Relationship: Recruitment and Fidelity of the Crab Trapezia on Its Coral Host Pocillopora

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
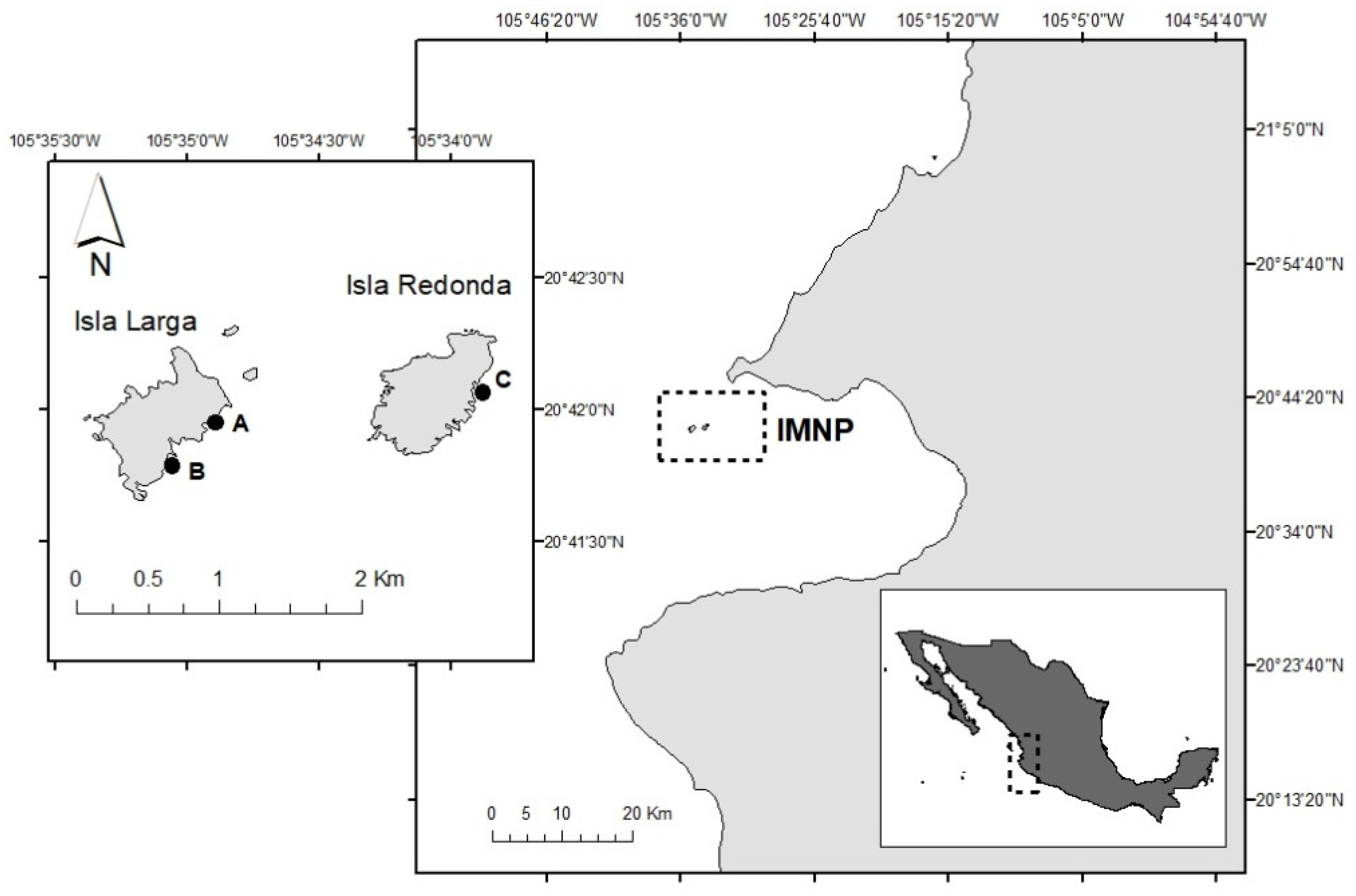
2.1. Study Area
2.2. Field Experiment
2.3. Data Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glynn, P.W.; Enochs, I.C. Invertebrates and their roles in coral reef ecosystems. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer Press: Dordrecht, The Netherlands, 2011; pp. 273–325. [Google Scholar] [CrossRef]
- Enochs, I.C.; Glynn, P.W. Corallivory in the Eastern Pacific. In Coral Reefs of the Eastern Tropical Pacific: Persistence and Loss in a Dynamic Environment; Glynn, P.W., Manzello, D.P., Enochs, I.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 8. [Google Scholar] [CrossRef]
- Garth, J.S. New brachyuran crabs from the Galapagos Islands. Allan Hancock Pac. Exped. 1939, 5, 9–49. [Google Scholar]
- Garth, J.S. The Crustacea Decapoda (Brachyura and Anomura) of Eniwetok Atoll, Marshall Islands, with special reference to the obligate commensals of branching corals. Micronesica 1964, 1, 137–144. [Google Scholar]
- Montano, S. The extraordinary importance of coral-associated fauna. Divers 2020, 12, 357. [Google Scholar] [CrossRef]
- Karlson, R.H. Dynamics of Coral Communities, 1st ed.; Kluwer Academic Publishers: London, UK, 1999; pp. 40–55. [Google Scholar]
- Castro, P. Movements between coral colonies in Trapezia ferruginea (Crustacea: Brachyura), an obligate symbiont of scleractinian corals. Mar. Biol. 1978, 46, 237–245. [Google Scholar] [CrossRef]
- Stella, J.S.; Pratchett, M.; Hutchings, P.; Jones, G. Coral-associated invertebrates: Diversity, ecological importance and vulnerability to disturbance. Oceanogr. Mar. Biol. 2011, 49, 43–104. [Google Scholar]
- Baeza, J.A. Crustaceans as symbionts: An overview of their diversity, host use and life styles. In Lifestyles and Feeding Biology. The Natural History of the Crustacea, 1st ed.; Thiel, M., Watling, L., Eds.; Oxford University Press: New York, NY, USA, 2015; Volume 2, pp. 163–189. [Google Scholar]
- Montano, S.; Fattorini, S.; Parravicini, V.; Berumen, M.L.; Galli, P.; Maggioni, D.; Arrigoni, R.; Seveso, D.; Strona, G. Corals hosting symbiotic hydrozoans are less susceptible to predation and disease. Proc. R. Soc. Lond. B Biol. Sci. 2017, 284, 20172405. [Google Scholar] [CrossRef] [Green Version]
- Pratchett, M.S. Influence of coral symbionts on feeding preferences of crown-of-thorns starfish Acanthaster planci in the western Pacific. Mar. Ecol. Prog. Ser. 2001, 214, 111–119. [Google Scholar] [CrossRef]
- Rouzé, H.; Lecellier, G.; Mills, S.C.; Planes, S.; Berteaux-Lecellier, V.; Stewart, H. Juvenile Trapezia spp. crabs can increase juvenile host coral survival by protection from predation. Mar. Ecol. Prog. Ser. 2014, 515, 151–159. [Google Scholar] [CrossRef]
- Pollock, F.J.; Katz, S.M.; Bourne, D.G.; Willis, B.L. Cymo melanodactylus crabs slow progression of white syndrome lesions on corals. Coral Reefs 2013, 32, 43–48. [Google Scholar] [CrossRef]
- McKeon, C.S.; Moore, J.M. Species and size diversity in protective services offered by coral guard-crabs. PeerJ 2014, 2, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garth, J.S. Decapod crustaceans inhabiting reef-building corals of Ceylon and the Maldive Islands. J. Mar. Biol. Assoc. India. 1974, 15, 195–212. [Google Scholar]
- Glynn, P.W.; Ault, J.S. A biogeographic analysis and review of the far eastern Pacific coral reef region. Coral Reefs 2000, 19, 1–23. [Google Scholar] [CrossRef]
- Lee, S.Y. Ecology of brachyura. In Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea, 1st ed.; Castro, P., Davie, P.J.F., Guinot, D., Schram, F.R., von Vaupel Klein, J.C., Eds.; Brill: Boston, MA, USA, 2015; Volume 9, Part C (2 vols); pp. 469–541. [Google Scholar]
- Stewart, H.L.; Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Symbiotic crabs maintain coral health by clearing sediments. Coral Reefs 2006, 25, 609–615. [Google Scholar] [CrossRef]
- Mclay, C.L.; Becker, C. Reproduction in brachyura. In Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea, 1st ed.; Castro, P., Davie, P.J.F., Guinot, D., Schram, F.R., von Vaupel Klein, J.C., Eds.; Brill: Boston, MA, USA, 2015; Volume 9, Part C (2 vols); pp. 185–243. [Google Scholar]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Annu. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef]
- Sanford, E.; Kelly, M.W. Local adaptation in marine invertebrates. Annu. Rev. Mar. Sci. 2011, 3, 509–535. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Mayfield, A.B.; Cupul-Magaña, A.L. Temporal variation in invertebrate recruitment on an Eastern Pacific coral reef. J. Sea Res. 2019, 145, 8–15. [Google Scholar] [CrossRef]
- Patton, W.K. Community structure among the animals inhabiting the coral Pocillopora damicornis at Heron Island Australia. In Symbiosis in the Sea, 1st ed.; Vernberg, W., Ed.; Univ. South Carolina Press: Columbia, SC, USA, 1974; pp. 219–243. [Google Scholar]
- CONANP (Comisión Nacional de Áreas Naturales Protegidas). Programa de Conservación Y Manejo, Parque Nacional Islas Marietas; Secretaria de Medio Ambiente y Recursos Naturales (SEMARENAT): México City, México, 2007; pp. 21–57. [Google Scholar]
- Wyrtki, K. Surface currents of the eastern tropical Pacific Ocean. Bull. Inter-Am. Trop. Tuna Commn. 1965, 9, 268–305. [Google Scholar]
- Plata, L.; Filonov, A. Marea interna en la parte noroeste de la Bahía de Banderas México. Cienc. Mar. 2007, 33, 197–215. [Google Scholar]
- Portela, W.; Beier, E.; Barton, E.D.; Castro, R.; Godínez, V.; Palacios Hernández, E.; Fiedler, P.C.; Sánchez-Velazco, L.; Trasviña, A. Water masses and circulation in the tropical pacific off Central Mexico and surrounding areas. J. Phys. Oceanogr. 2016, 46, 3069–3081. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Bonilla, H.; Calderón Aguilera, L.E.; Cruz-Piñón, G.; Medina-Rosas, P.; López-Perez, R.A.; Herrero-Perezrul, M.D.; Leyte-Morales, G.E.; Cupul-Magaña, A.L.; Carriquiry-Beltran, J.D. Atlas de Corales Pétreos (Anthozoa: Scleractinia) del Pacífico Mexicano, 1st ed.; Sociedad Mexicana de Arrecifes Coralinos AC, CICESE, CONABIO, CONACYT, DBM/UABCS, CUC/UdeG, Umar: Mexico City, Mexico, 2005; p. 129. [Google Scholar]
- Hernández-Zulueta, J.; Rodríguez-Zaragoza, F.A.; Araya, R.; Vargas-Ponce, O.; Rodríguez-Troncoso, A.P.; Cupul-Magaña, A.L.; Díaz-Pérez, L.; Ríos-Jara, E.; Ortiz, M. Multi-scale analysis of hermatypic coral assemblages at Mexican Central Pacific. Sci. Mar. 2017, 81, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Sotelo-Casas, R.C.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Solís-Marín, F.A.; Godínez-Domínguez, E.; Cupul-Magaña, A.L. Spatial-temporal variations in echinoderm diversity within coral communities in a transitional region of the northeast of the eastern pacific. Estuar. Coast. Shelf Sci. 2019, 227, 106346. [Google Scholar] [CrossRef]
- Cruz-García, R.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Mayfield, A.; Cupul-Magaña, A.L. Ephemeral effects of El Niño–Southern Oscillation events on an eastern tropical Pacific coral community. Mar. Freshw. Res. 2020, 71, 1259–1268. [Google Scholar] [CrossRef]
- Edwards, A.J. Reef Rehabilitation Manual; The Coral Reef Targeted Research and Capacity Building for Management Program: Queensland, Australia, 2010; pp. 64–108. [Google Scholar]
- Tortolero-Langarica, J.J.A.; Cupul-Magaña, A.L.; Rodríguez-Troncoso, A.P. Restoration of a degraded coral reef using a natural remediation process: A case study from a Central Mexican Pacific National Park. Ocean Coast. Manag. 2014, 96, 12–19. [Google Scholar] [CrossRef]
- Barry, C.K. Ecological Study of the Decapod Crustaceans Commensal with the Branching Coral Pocillopora meandrina var. Nobilis Verrill. Master’s Thesis, University of Hawaii, Honolulu, HI, USA, 1965. [Google Scholar]
- Highsmith, R.C. Reproduction by fragmentation in corals. Mar. Ecol. Prog. Ser. 1982, 7, 207–226. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008; p. 214. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; p. 192. [Google Scholar]
- Castro, P. Animal symbioses in coral reef communities: A review. Symbiosis 1988, 5, 161–184. [Google Scholar]
- Abele, L.G.; Patton, W.K. The size of coral heads and the community biology of associated decapod crustaceans. J. Biogeogr. 1976, 35–47. [Google Scholar] [CrossRef]
- Stewart, H.L.; Price, N.N.; Holbrook, S.J.; Schmitt, R.J.; Brooks, A.J. Determinants of the onset and strength of mutualistic interactions between branching corals and associate crabs. Mar. Ecol. Prog. Ser. 2013, 493, 155–163. [Google Scholar] [CrossRef]
- Preston, E.M. A computer simulation of competition among five sympatric congeneric species of xanthid crabs. Ecology 1973, 54, 469–483. [Google Scholar] [CrossRef]
- Huber, M.E. Aggressive behavior of Trapezia intermedia miers and T. digitalis Latreille (Brachyura: Xanthidae). J. Crust. Biol. 1987, 7, 238–248. [Google Scholar] [CrossRef]
- Vannini, M. A shrimp that speaks crab-ese. J. Crust. Biol. 1985, 5, 160–167. [Google Scholar] [CrossRef]
- Roughgarden, J. Evolution of marine symbiosis—A simple cost-benefit model. Ecology 1975, 56, 1201–1208. [Google Scholar] [CrossRef]
- Thiel, M.; Zander, A.; Baeza, J.A. Movements of the symbiotic crab Liopetrolisthes mitra between hosts, black sea urchins Tetrapygus niger. Bull. Mar. Sci. 2003, 72, 89–101. [Google Scholar]
- Knowlton, N. Sexual selection and dimorphism in two demes of a symbiotic, pair-bonding snapping shrimp. Evolution 1980, 34, 161–173. [Google Scholar] [CrossRef]
- Bell, J.L. Changing residence: Dynamics of the symbiotic relationship between Dissodactylus mellitae Rathbun (Pinnotheridae) and Mellita quinquiesperforata (Leske) (Echinodermata). J. Exp. Mar. Biol. Ecol. 1984, 82, 101–115. [Google Scholar] [CrossRef]
- Hartnoll, R.G.; Bryant, A.D. Growth to maturity of juveniles of the spider crabs Hyas coarctatus Leach and Inachus dorsettensis (Pennant) (Brachyura: Majidae). J. Exp. Mar. Biol. Ecol. 2001, 263, 143–158. [Google Scholar] [CrossRef]
- Azra, M.N.; Aaqillah-Amr, M.A.; Ikhwanuddin, M.; Ma, H.; Waiho, K.; Ostrensky, A.; Prestes dos Santos Tavares, C.; Abol-Munafi, A.B. Effects of climate-induced water temperature changes on the life history of brachyuran crabs. Rev. Aquac. 2020, 12, 1211–1216. [Google Scholar] [CrossRef]
- Stella, J.S.; Munday, P.L.; Walker, S.P.W.; Pratchett, M.S.; Jones, G.P. From cooperation to combat: Adverse effect of thermal stress in a symbiotic coral-crustacean community. Oecologia 2013, 174, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, M.; Taira, A. Population structure of six sympatric species of Trapezia associated with the hermatypic coral Pociliopora damicornis with a hypothesis of mechanisms promoting their coexistence. J. Jpn. Coral Reef Soc. 1999, 9–17. [Google Scholar] [CrossRef]
- Knudsen, J.W. Trapezia and Tetralia (Decapoda, Brachyura, Xanthidae) as obligate ectoparasites of pocilloporid and acroporid corals. Pac. Sci. 1967, 21, 51–57. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Habitat Space | Recruitment | ||||
|---|---|---|---|---|---|---|
| Pseudo-F | P (perm) | C.V.(%) | Pseudo-F | P (perm) | C.V.(%) | |
| Time | 145.44 | 0.0001 * | 71.7 | 108.1 | 0.0001 * | 70.9 |
| Residuals | 21.9 | 29.1 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canizales-Flores, H.M.; Rodríguez-Troncoso, A.P.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L. A Long-Term Symbiotic Relationship: Recruitment and Fidelity of the Crab Trapezia on Its Coral Host Pocillopora. Diversity 2021, 13, 450. https://doi.org/10.3390/d13090450
Canizales-Flores HM, Rodríguez-Troncoso AP, Rodríguez-Zaragoza FA, Cupul-Magaña AL. A Long-Term Symbiotic Relationship: Recruitment and Fidelity of the Crab Trapezia on Its Coral Host Pocillopora. Diversity. 2021; 13(9):450. https://doi.org/10.3390/d13090450
Chicago/Turabian StyleCanizales-Flores, H. M., A. P. Rodríguez-Troncoso, F. A. Rodríguez-Zaragoza, and A. L. Cupul-Magaña. 2021. "A Long-Term Symbiotic Relationship: Recruitment and Fidelity of the Crab Trapezia on Its Coral Host Pocillopora" Diversity 13, no. 9: 450. https://doi.org/10.3390/d13090450
APA StyleCanizales-Flores, H. M., Rodríguez-Troncoso, A. P., Rodríguez-Zaragoza, F. A., & Cupul-Magaña, A. L. (2021). A Long-Term Symbiotic Relationship: Recruitment and Fidelity of the Crab Trapezia on Its Coral Host Pocillopora. Diversity, 13(9), 450. https://doi.org/10.3390/d13090450