
Multi-Gene Phylogeny and Taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Sources
2.2. Morphological Characterization
2.3. DNA Extraction, PCR Amplification, and Sequencing
2.4. Phylogenetic Analysis
3. Results
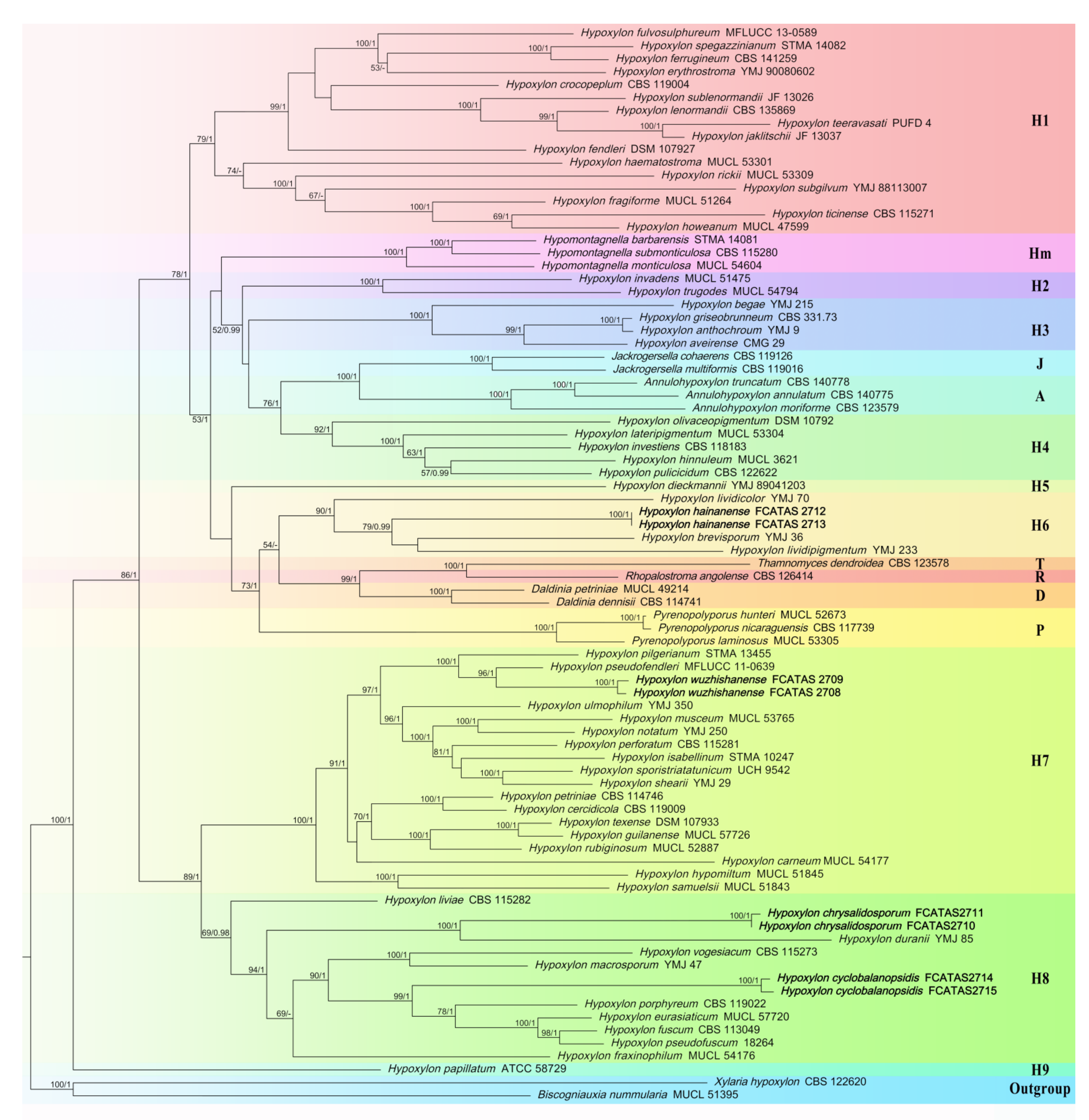
3.1. Phylogenetic Analysis
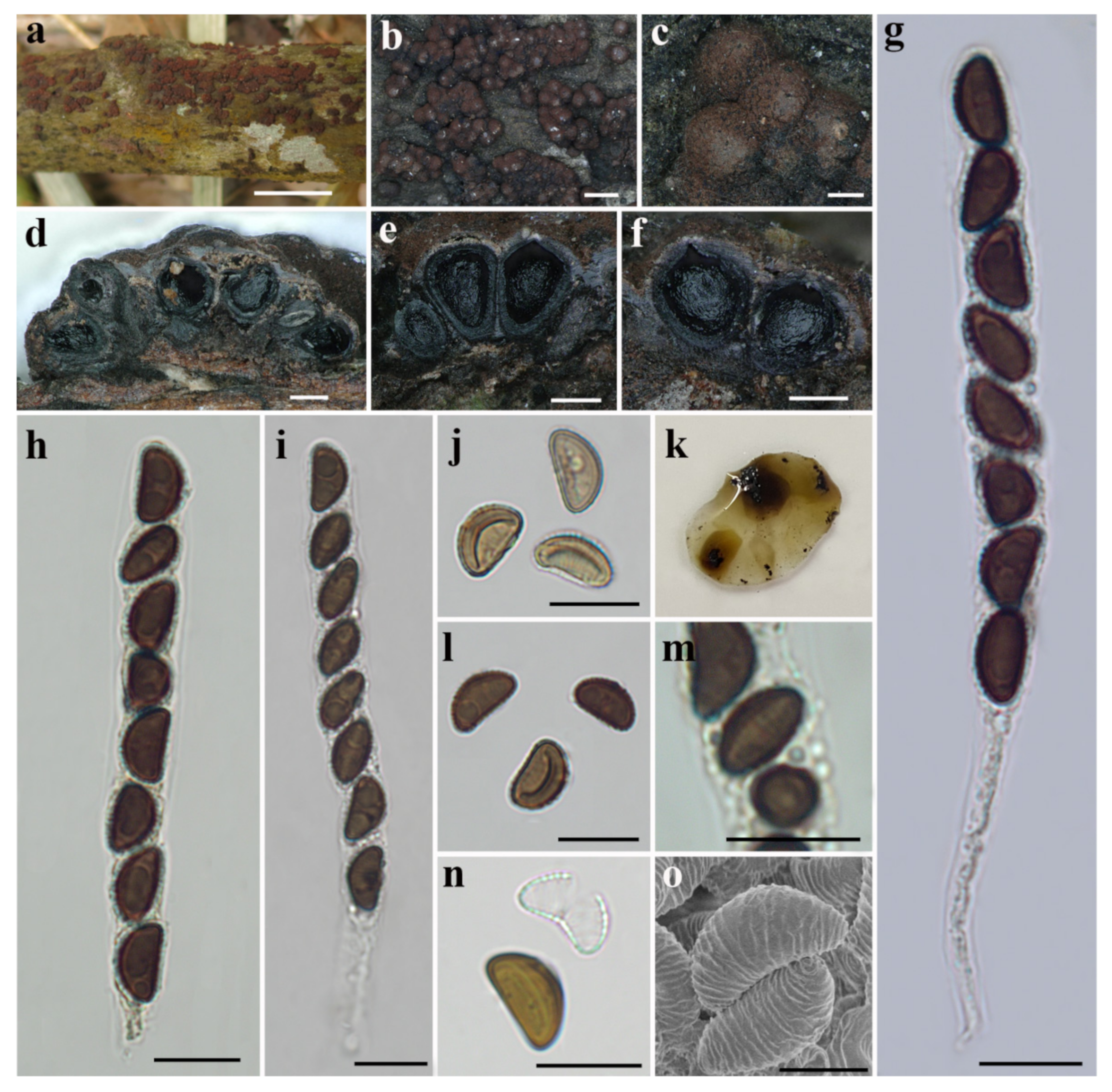
3.2. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, J.H. A Monograph of the World Species of Hypoxylon; University Georgia Press: Athens, Greece, 1961; p. 158. [Google Scholar]
- Rogers, J.D. The Xylariaceae: Systematic, biological and evolutionary aspects. Mycologia 1979, 71, 1–42. [Google Scholar] [CrossRef]
- Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny, and functional biodiversity. Curr. Res. Envion. Appl. Mycol. 2011, 1, 75–133. [Google Scholar] [CrossRef]
- Ju, Y.M.; Rogers, J.D. A revision of the Genus Hypoxylon; American Phytopathological Society Press: St. Paul, MN, USA, 1996; p. 365. [Google Scholar]
- Rogers, J.D. Thoughts and musings on tropical Xylariaceae. Mycol. Res. 2000, 104, 1412–1420. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Per, D.; Luangsaard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Pourmoghaddam, M.J.; Lambert, C.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stadler, M. Discovery of a new species of the Hypoxylon rubiginosum complex from Iran and antagonistic activities of Hypoxylon spp. Against the Ash Dieback pathogen, Hymenoscyphus fraxineus, in dual culture. MycoKeys 2020, 66, 105–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, J.; Lechat, C.; Courtecuisse, R. The genus Hypoxylon (Xylariaceae) in Guadeloupe and Martinique (French West Indies). Ascomycete.org 2016, 7, 145–212. [Google Scholar]
- Sir, E.B.; Kuhnert, E.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Stadler, M. New species and reports of Hypoxylon from Argentina recognized by a polyphasic approach. Mycol. Prog. 2016, 15, 42. [Google Scholar] [CrossRef]
- Rogers, J.D. Two new Hypoxylon species from Gabon. Can. J. Bot. 1981, 59, 1363–1364. [Google Scholar] [CrossRef]
- Rogers, J.D.; Candoussau, F. A new variety of Hypoxylon cohaerens from France. Mycologia 1980, 72, 826–829. [Google Scholar] [CrossRef]
- Rogers, J.D. Hypoxylon weldenii var. microsporum and H. punctidiscum. Mycologia 1980, 72, 829–832. [Google Scholar] [CrossRef]
- Rogers, J.D.; Ju, Y.M.; Hemmes, D.E. Hypoxylon rectangulosporum sp. nov., Xylaria psidii sp. nov., and comments on taxa of Podosordaria and Stromatoneurospora. Mycologia 1992, 84, 166–172. [Google Scholar] [CrossRef]
- Gucht, K.V.; Ju, Y.M.; Rogers, J.D. New Hypoxylon species from Papua New Guinea and notes on some other taxa. Mycologia 1997, 89, 503–511. [Google Scholar] [CrossRef]
- Hsieh, H.; Ju, Y.M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef]
- Lambert, C.; Wendt, L.; Hladki, A.I.; Stadler, M.; Sir, E.B. Hypomontagnella (Hypoxylaceae): A new genus segregated from Hypoxylon by a polyphasic taxonomic approach. Mycol. Prog. 2019, 18, 187–201. [Google Scholar] [CrossRef]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsaard, J.J.; Srikitikulchai, P.; Per, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef] [Green Version]
- Teng, S.C. Fungi of China; Science Press: Beijing, China, 1963; pp. 182–191. [Google Scholar]
- Tai, F.L. Sylloge Fungorum Sinicorum; Science Press: Beijing, China, 1979; pp. 163–167. [Google Scholar]
- Ma, H.X. Taxonomy and Molecular Phylogeny of Several Genera of Xylariaceae from China. Ph.D. Thesis, Jilin Agricultural University, Changchun, China, June 2011. [Google Scholar]
- Chi, S.Q.; Xu, J.; Lu, B.S. Three New Chinese Records of Hypoxylon. J. Fungal Res. 2016, 14, 218–221. [Google Scholar]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute: London, UK, 1970. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- O’donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics—Science Direct; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.X.; Qiu, J.Z.; Xu, B.; Li, Y. Two Hypoxylon species from Yunnan Province based on morphological and molecular characters. Phytotaxa 2018, 376, 027–036. [Google Scholar] [CrossRef]
- Kuhnert, E.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Rohde, M.; Stadler, M. Phylogenetic and chemotaxonomic resolution of the genus Annulohypoxylon (Xylariaceae) including four new species. Fungal Divers. 2017, 85, 1–43. [Google Scholar] [CrossRef]
- Stadler, M.; Læssøe, T.; Fournier, J.; Decock, C.; Schmieschek, B.; Tichy, H.V.; Peršoh, D. A polyphasic taxonomy of Daldinia (Xylariaceae). Stud. Mycol. 2014, 77, 1–143. [Google Scholar] [CrossRef] [Green Version]
- Vicente, T.F.L.; Goncalves, M.F.M.; Brandao, C.; Fidalgo, C.; Alves, A. Diversity of fungi associated with macroalgae from an estuarine environment and description of Cladosporium rubrum sp. nov. and Hypoxylon aveirense sp. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 004630. [Google Scholar] [CrossRef]
- Lambert, C.; Pourmoghaddam, M.J.; Cedeño-Sanchez, M.; Surup, F.; Khodaparast, S.A.; Krisai-Greilhuber, I.; Voglmayr, H.; Stradal, T.E.B.; Stadler, M. Resolution of the Hypoxylon fuscum complex (Hypoxylaceae, Xylariales) and discovery and biological characterization of two of its prominent secondary metabolites. J. Fungi 2021, 7, 131. [Google Scholar] [CrossRef]
- Sir, E.B.; Becker, K.; Lambert, C.; Bills, G.F.; Kuhnert, E. Observations on Texas hypoxylons, including two new Hypoxylon species and widespread environmental isolates of the H. croceum complex identified by a polyphasic approach. Mycologia 2019, 11, 832–856. [Google Scholar] [CrossRef]
- Friebes, G.; Wendelin, I. Studies on Hypoxylon ferrugineum (Xylariaceae), a rarely reported species collected in the urban area of Graz (Austria). Ascomycete.org 2016, 8, 83–90. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.Z.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Sir, E.B.; Kuhnert, E.; Surup, F.; Hyde, K.D.; Stadler, M. Discovery of new mitorubrin derivatives from Hypoxylon fulvosulphureum sp. nov. (Ascomycota, Xylariales). Mycol. Prog. 2015, 14, 28. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Bitzer, J.; Læssøe, T.; Fournier, J.; Kummer, V.; Decock, C.; Tichy, H.V.; Piepenbring, M.; Peršoh, D.; Stadler, M. Affinities of Phylacia and the daldinoid Xylariaceae, inferred from chemotypes of cultures and ribosomal DNA sequences. Mycol. Res. 2008, 112, 251–270. [Google Scholar] [CrossRef]
- Becker, K.; Lambert, C.; Wieschhaus, J.; Stadler, M. Phylogenetic assignment of the fungicolous Hypoxylon invadens (Ascomycota, Xylariales) and investigation of its secondary metabolites. Microorganisms 2020, 8, 1397. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, E.; Surup, F.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Stadler, M. Lenormandins A—G, new azaphilones from Hypoxylon lenormandii and Hypoxylon jaklitschii sp. nov., recognised by chemotaxonomic data. Fungal Divers. 2015, 71, 165–184. [Google Scholar] [CrossRef]
- Dai, D.Q.; Phookamsak, R.; Wijayawardene, N.N.; Li, W.J.; Bhat, D.J.; Xu, J.C.; Taylor, J.E.; Hyde, K.D.; Chukeatirote, E. Bambusicolous fungi. Fungal Divers. 2017, 82, 1–105. [Google Scholar] [CrossRef]
- Bills, G.F.; González-Menéndez, V.; Martín, J.; Platas, G.; Fournier, J.; Peršoh, D.; Stadler, M. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 2012, 7, e46687. [Google Scholar] [CrossRef] [PubMed]
- Stadler, M.; Kuhnert, E.; Peršoh, D.; Fournier, J. The Xylariaceae as model example for a unified nomenclature following the “One Fungus-One Name” (1F1N) concept. Mycology 2013, 4, 5–21. [Google Scholar]
- Cedeño-Sanchez, M.; Wendt, L.; Stadler, M.; Mejía, L.C. Three new species of Hypoxylon and new records of Xylariales from Panama. Mycosphere 2020, 11, 1457–1476. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.; Raspe, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Stadler, M.; Fournier, J.; Laessøe, T.; Chlebicki, A.; Lechat, C.; Flessa, F.; Rambold, G.; Peršoh, D. Chemotaxonomic and phylogenetic studies of Thamnomyces (Xylariaceae). Mycoscience 2010, 51, 189–207. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 9, 1312–1313. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, H.; Lin, C.; Fang, M.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Daranagama, D.A.; Hyde, K.D.; Sir, E.B.; Thambugala, K.M.; Stadler, M. Towards a natural classification and backbone tree for Graphostromataceae, Hypoxylaceae, Lopadostomataceae and Xylariaceae. Fungal Divers. 2018, 88, 1–165. [Google Scholar] [CrossRef]
- Long, Q.D.; Liu, L.L.; Zhang, X.; Wen, T.C.; Kang, J.C.; Hyde, K.D.; Shen, X.C.; Li, Q.R. Contributions to species of Xylariales in China-1. Durotheca species. Mycol. Prog. 2019, 18, 495–510. [Google Scholar] [CrossRef]
- Rambaut, A. Figtree Version 1.4.2. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 19 January 2018).
- Granmo, A. Morphotaxonomy and Chorology of the Genus Hypoxylon (Xylariaceae) in Norway. Sommerfeltia 1999, 26, 1–81. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Beltrán-Tejera, E.; Granmo, A. The “red Hypoxylons” of the temperate and subtropical Northern Hemisphere. N. Am. Fungi 2008, 3, 73–125. [Google Scholar] [CrossRef] [Green Version]
- Stadler, M.; Fournier, J. Pigment chemistry, taxonomy and phylogeny of the Hypoxyloideae (Xylariaceae). Rev. Iberoam. Micol. 2006, 23, 160–170. [Google Scholar] [CrossRef]
- Stadler, M.; Fournier, J.; Quang, D.N.; Akulov, A.Y. Metabolomic studies on the chemical ecology of the Xylariaceae (Ascomycota). Nat. Prod. Commun. 2007, 2, 287–304. [Google Scholar] [CrossRef] [Green Version]
- Hennings, P. Fungi mattogrossenses a Dr. R. Pilger collecti 1899. Hedwigia. 1900, 39, 134–139. [Google Scholar]
- Wibberg, D.; Stadler, M.; Lambert, C.; Bunk, B.; Spröer, C.; Rückert, C.; Kalinowski, J.; Cox, R.J.; Kuhnert, E. High quality genome sequences of thirteen Hypoxylaceae (Ascomycota) strengthen the phylogenetic family backbone and enable the discovery of new taxa. Fungal Divers. 2021, 106, 7–28. [Google Scholar] [CrossRef]
- Kuhnert, E.; Surup, F.; Herrmann, J.; Huch, V.; Müller, R.; Stadler, M. Rickenyls A-E, antioxidative terphenyls from the fungus Hypoxylon rickii (Xylariaceae, Ascomycota). Phytochemistry 2015, 118, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, E.; Navarro-Muñoz, J.C.; Becker, K.; Stadler, M.; Collemare, J.; Cox, R.J. Secondary metabolite biosynthetic diversity in the fungal family Hypoxylaceae and Xylaria hypoxylon. Stud. Mycol. 2021, 99, 100118. [Google Scholar] [CrossRef]
- Leman-Loubiere, C.; Le, G.G.; Debitus, C.; Ouazzani, J. Sporochartines A-E, a new family of natural products from the marine fungus Hypoxylon monticulosum isolated from a Sphaerocladina sponge. Front. Mar. Sci. 2017, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Surup, F.; Kuhnert, E.; Lehmann, E.; Heitkämper, S.; Hyde, K.D.; Fournier, J.; Stadler, M. Sporothriolide derivatives as chemotaxonomic markers for Hypoxylon monticulosum. Mycology 2014, 5, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surup, F.; Kuhnert, E.; Böhm, A.; Pendzialek, T.; Solga, D.; Wiebach, V.; Engler, H.; Berkessel, A.; Stadler, M.; Kalesse, M. The rickiols: 20-, 22-, and 24-membered macrolides from the ascomycete Hypoxylon rickii. Chemistry Eur. J. 2018, 24, 2200–2213. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.X.; Vasilyeva, L.; Li, Y. Hypoxylon from China-2: H. dengii sp. nov and H. crocopeplum new to China. Mycotaxon 2012, 122, 1–5. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Specimen No. | Locality | GenBank Accession No. | Status | Reference | |||
|---|---|---|---|---|---|---|---|---|
| ITS | LSU | RPB2 | β-tubulin | |||||
| Annulohypoxylon annulatum | CBS 140775 | USA | KU604559 | KY610418 | KY624263 | KX376353 | ET | [10,18,30] |
| A. moriforme | CBS 123579 | Martinique | KX376321 | KY610425 | KY624289 | KX271261 | [30] | |
| A. truncatum | CBS 140778 | USA | KX376329 | KY610419 | KY624277 | KX376352 | ET | [18,30] |
| Daldinia dennisii | CBS 114741 | Australia | JX658477 | KY610435 | KY624244 | KC977262 | T | [6,18,31] |
| D. petriniae | MUCL 49214 | Austria | JX658512 | KY610439 | KY624248 | KC977261 | ET | [6,18,31] |
| Hypomontagnella barbarensis | STMA 14081 | Argentina | MK131720 | MK131718 | MK135891 | MK135893 | T | [17] |
| Hypom. monticulosa | MUCL 54604 | French Guiana | KY610404 | KY610487 | KY624305 | KX271273 | ET | [18] |
| Hypom. submonticulosa | CBS 115280 | France | KC968923 | KY610457 | KY624226 | KC977267 | [6,18] | |
| Hypoxylon anthochroum | YMJ 9 | Mexico | JN660819 | N/A | N/A | AY951703 | [16] | |
| H. aveirense | CMG 29 | Portugal | MN053021 | N/A | N/A | MN066636 | T | [32] |
| H. begae | YMJ 215 | USA | JN660820 | N/A | N/A | AY951704 | [16] | |
| H. brevisporum | YMJ 36 | Puerto Rico | JN660821 | N/A | N/A | AY951705 | [16] | |
| H. carneum | MUCL 54177 | France | KY610400 | KY610480 | KY624297 | KX271270 | [18] | |
| H. cercidicola | CBS 119009 | France | KC968908 | KY610444 | KY624254 | KX271270 | [6,18] | |
| H. chrysalidosporum | FCATAS2710 | China | OL467294 | OL615106 | OL584222 | OL584229 | T | This study |
| H. chrysalidosporum | FCATAS2711 | China | OL467295 | OL615107 | OL584223 | OL584230 | This study | |
| H. crocopeplum | CBS 119004 | France | KC968907 | KY610445 | KY624255 | KC977268 | [18] | |
| H. cyclobalanopsidis | FCATAS2714 | China | OL467298 | OL615108 | OL584225 | OL584232 | T | This study |
| H. cyclobalanopsidis | FCATAS2715 | China | OL467299 | OL615109 | OL584226 | OL584233 | This study | |
| H. dieckmannii | YMJ 89041203 | China | JN979413 | N/A | N/A | AY951713 | [16] | |
| H. duranii | YMJ 85 | China | JN979414 | N/A | N/A | AY951714 | [16] | |
| H. erythrostroma | YMJ 90080602 | China | JN979416 | N/A | N/A | AY951716 | [16] | |
| H. eurasiaticum | MUCL 57720 | Iran | MW367851 | N/A | MW373852 | MW373861 | [33] | |
| H. fendleri | DSM 107927 | USA | MK287533 | MK287545 | MK287558 | MK287571 | [34] | |
| H. ferrugineum | CBS 141259 | Austria | KX090079 | N/A | N/A | KX090080 | [35] | |
| H. fragiforme | MUCL 51264 | Germany | KM186294 | KM186295 | KM186296 | KM186293 | ET | [36] |
| H. fraxinophilum | MUCL 54176 | France | KC968938 | N/A | N/A | KC977301 | ET | [6] |
| H. fulvosulphureum | MFLUCC 13-0589 | Thailand | KP401576 | N/A | N/A | KP401584 | T | [37] |
| H. fuscum | CBS 113049 | France | KY610401 | KY610482 | KY624299 | KX271271 | ET | [18] |
| H. griseobrunneum | CBS 331.73 | India | KY610402 | MH872399 | KY624300 | KC977303 | T | [6,18,38] |
| H. guilanense | MUCL 57726 | Iran | MT214997 | MT214992 | MT212235 | MT212239 | T | [8] |
| H. haematostroma | MUCL 53301 | Martinique | KC968911 | KY610484 | KY624301 | KC977291 | ET | [6,18] |
| H.hainanense | FCATAS2712 | China | OL467296 | OL616132 | OL584224 | OL584231 | T | This study |
| H.hainanense | FCATAS2713 | China | OL467297 | N/A | N/A | N/A | This study | |
| H. hinnuleum | MUCL 3621 | USA | MK287537 | MK287549 | MK287562 | MK287575 | T | [36] |
| H. howeanum | MUCL 47599 | Germany | AM749928 | KY610448 | KY624258 | KC977277 | [6,18,39] | |
| H. hypomiltum | MUCL 51845 | Guadeloupe | KY610403 | KY610449 | KY624302 | KX271249 | [18] | |
| H. invadens | MUCL 51475 | France | MT809133 | MT809132 | MT813037 | MT813038 | T | [40] |
| H. investiens | CBS 118183 | Malaysia | KC968925 | KY610450 | KY624259 | KC977270 | [6,18] | |
| H. isabellinum | STMA10247 | Martinique | STMA10247 | N/A | N/A | KC977295 | T | [6] |
| H. jaklitschii | JF 13037 | Sri Lanka | KM610290 | N/A | N/A | KM610304 | T | [41] |
| H. lateripigmentum | MUCL 53304 | Martinique | KC968933 | KY610486 | KY624304 | KC977290 | T | [6,18] |
| H. lenormandii | CBS 135869 | Cameroon | KY610390 | KY610453 | KY624262 | KM610295 | [18,41] | |
| H. liviae | CBS 115282 | Norway | NR155154 | N/A | N/A | KC977265 | ET | [6] |
| H. lividicolor | YMJ 70 | China | JN979432 | N/A | N/A | AY951734 | [16] | |
| H. lividipigmentum | YMJ 233 | Mexico | JN979433 | N/A | N/A | AY951735 | [16] | |
| H. macrosporum | YMJ 47 | Canada | JN979434 | N/A | N/A | AY951736 | [16] | |
| H. musceum | MUCL 53765 | Guadeloupe | KC968926 | KY610488 | KY624306 | KC977280 | [6,18] | |
| H. notatum | YMJ 250 | USA | JQ009305 | N/A | N/A | AY951739 | [16] | |
| H. olivaceopigmentum | DSM 10792 | USA | MK287530 | MK287542 | MK287555 | MK287568 | T | [34] |
| H. papillatum | ATCC 58729 | USA | NR155153 | KY610454 | KY624223 | KC977258 | T | [6,18] |
| H. perforatum | CBS 115281 | France | KY610391 | KY610455 | KY624224 | KX271250 | [18] | |
| H. petriniae | CBS 114746 | France | NR155185 | KY610491 | KY624279 | KX271274 | T | [18] |
| H. pilgerianum | STMA 13455 | Martinique | KY610412 | N/A | KY624308 | KY624315 | [18] | |
| H. porphyreum | CBS 119022 | France | KC968921 | KY610456 | KY624225 | KC977264 | [6,18] | |
| H. pseudofendleri | MFLUCC 11-0639 | Thailand | KU940156 | KU863144 | N/A | N/A | [42] | |
| H. pseudofuscum | 18264 | Germany | MW367857 | MW367848 | MW373858 | MW373867 | T | [33] |
| H. pulicicidum | CBS 122622 | Martinique | JX183075 | KY610492 | KY624280 | JX183072 | T | [18,43] |
| H. rickii | MUCL 53309 | Martinique | KC968932 | KY610416 | KY624281 | KC977288 | ET | [18] |
| H. rubiginosum | MUCL 52887 | Germany | KC477232 | KY610469 | KY624266 | KY624311 | ET | [18,44] |
| H. samuelsii | MUCL 51843 | Guadeloupe | KC968916 | KY610466 | KY624269 | KC977286 | ET | [6,18] |
| H. shearii | YMJ 29 | Mexico | EF026142 | N/A | N/A | AY951753 | [16] | |
| H. spegazzinianum | STMA 14082 | Argentina | KU604573 | N/A | N/A | KU604582 | T | [10] |
| H. sporistriatatunicum | UCH9542 | Panama | MN056426 | N/A | N/A | MK908140 | T | [45] |
| H. subgilvum | YMJ 88113007 | China | JQ009315 | N/A | N/A | AY951755 | [16] | |
| H. sublenormandii | JF 13026 | Sri Lanka | KM610291 | N/A | N/A | KM610303 | T | [41] |
| H. teeravasati | PUFD 4 | India | N/A | MF385274 | MG986895 | MG986894 | [46] | |
| H. texense | DSM 107933 | USA | MK287536 | MK287548 | MK287561 | MK287574 | T | [34] |
| H. trugodes | MUCL 54794 | Sri Lanka | KF234422 | NG066380 | KY624282 | KF300548 | ET | [6,18] |
| H. ulmophilum | YMJ 350 | Russia | JQ009320 | N/A | N/A | AY951760 | [16] | |
| H. vogesiacum | CBS 115273 | France | KC968920 | KY610417 | KY624283 | KX271275 | [18] | |
| H.wuzhishanense | FCATAS2708 | China | OL467292 | OL615104 | OL584220 | OL584227 | T | This study |
| H.wuzhishanense | FCATAS2709 | China | OL467293 | OL615105 | OL584221 | OL584228 | This study | |
| Jackrogersella cohaerens | CBS 119126 | Germany | KY610396 | KY610497 | KY624270 | KY624314 | [18] | |
| J. multiformis | CBS 119016 | Germany | KC477234 | KY610473 | KY624290 | KX271262 | ET | [6,18] |
| Pyrenopolyporus hunteri | MUCL 52673 | Ivory Coast | KY610421 | KY610472 | KY624309 | KU159530 | ET | [18,30] |
| P. laminosus | MUCL 53305 | Martinique | KC968934 | KY610485 | KY624303 | KC977292 | T | [6,18] |
| P. nicaraguensis | CBS 117739 | Burkina Faso | AM749922 | KY610489 | KY624307 | KC977272 | [6,18,39] | |
| Rhopalostroma angolense | CBS 126414 | Ivory Coas | KY610420 | KY610459 | KY624228 | KX271277 | [18] | |
| Thamnomyces dendroidea | CBS 123578 | French Guiana | FN428831 | KY610467 | KY624232 | KY624313 | T | [18,47] |
| Xylaria hypoxylon | CBS 122620 | Sweden | KY610407 | KY610495 | KY624231 | KX271279 | ET | [18] |
| Biscogniauxia nummularia | MUCL 51395 | France | KY610382 | KY610427 | KY624236 | KX271241 | [18] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, H.; Song, Z.; Pan, X.; Li, Y.; Yang, Z.; Qu, Z. Multi-Gene Phylogeny and Taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity 2022, 14, 37. https://doi.org/10.3390/d14010037
Ma H, Song Z, Pan X, Li Y, Yang Z, Qu Z. Multi-Gene Phylogeny and Taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity. 2022; 14(1):37. https://doi.org/10.3390/d14010037
Chicago/Turabian StyleMa, Haixia, Zikun Song, Xiaoyan Pan, Yu Li, Zhanen Yang, and Zhi Qu. 2022. "Multi-Gene Phylogeny and Taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China" Diversity 14, no. 1: 37. https://doi.org/10.3390/d14010037
APA StyleMa, H., Song, Z., Pan, X., Li, Y., Yang, Z., & Qu, Z. (2022). Multi-Gene Phylogeny and Taxonomy of Hypoxylon (Hypoxylaceae, Ascomycota) from China. Diversity, 14(1), 37. https://doi.org/10.3390/d14010037