
Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling


Abstract
:1. Introduction
2. Materials and Methods
2.1. Occurrence Data
2.2. Bioclimatic Covariates
2.3. Habitat Suitability Modeling
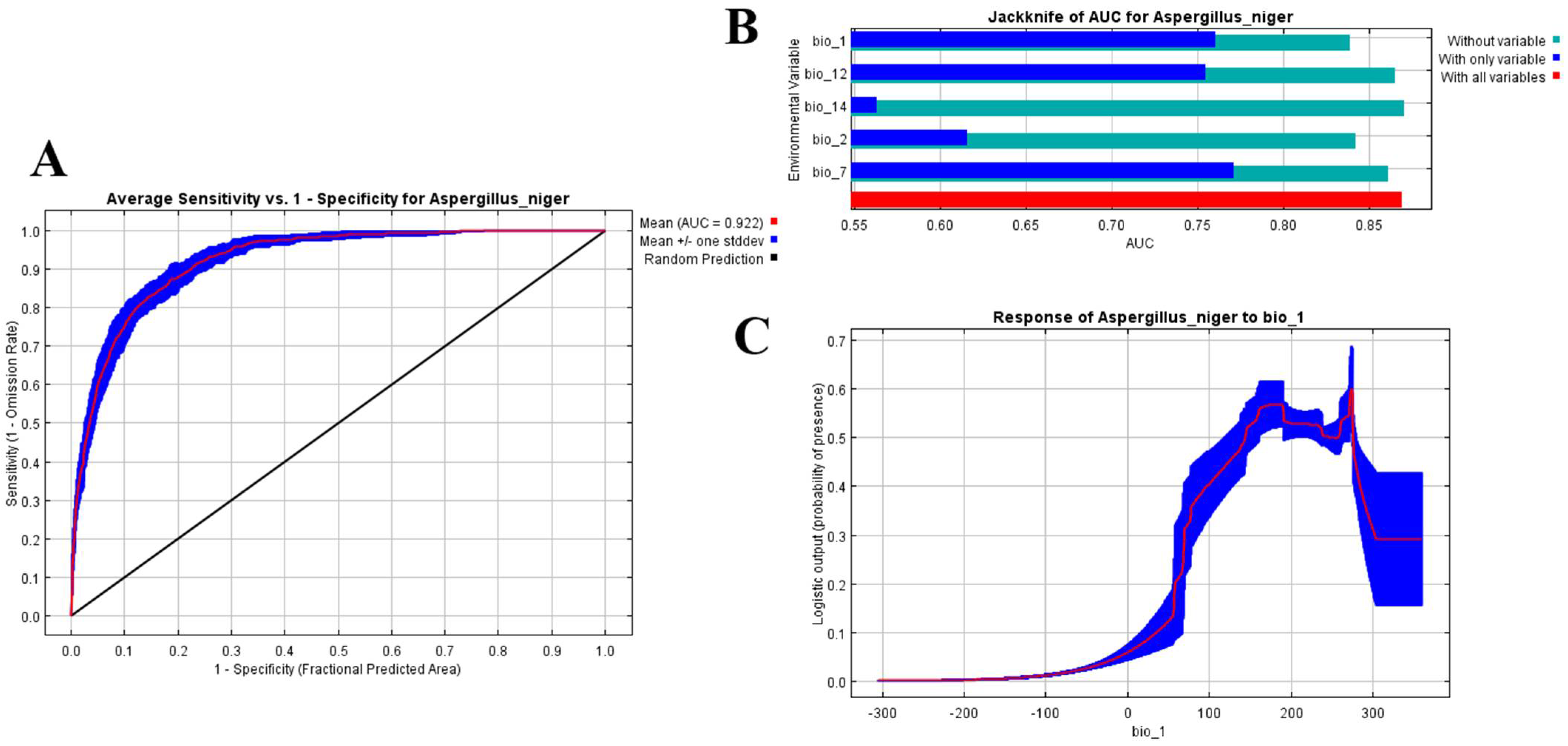
2.4. Model Evaluation
3. Results
3.1. Modeling Performance
3.2. Contribution of Bioclimatic Variables
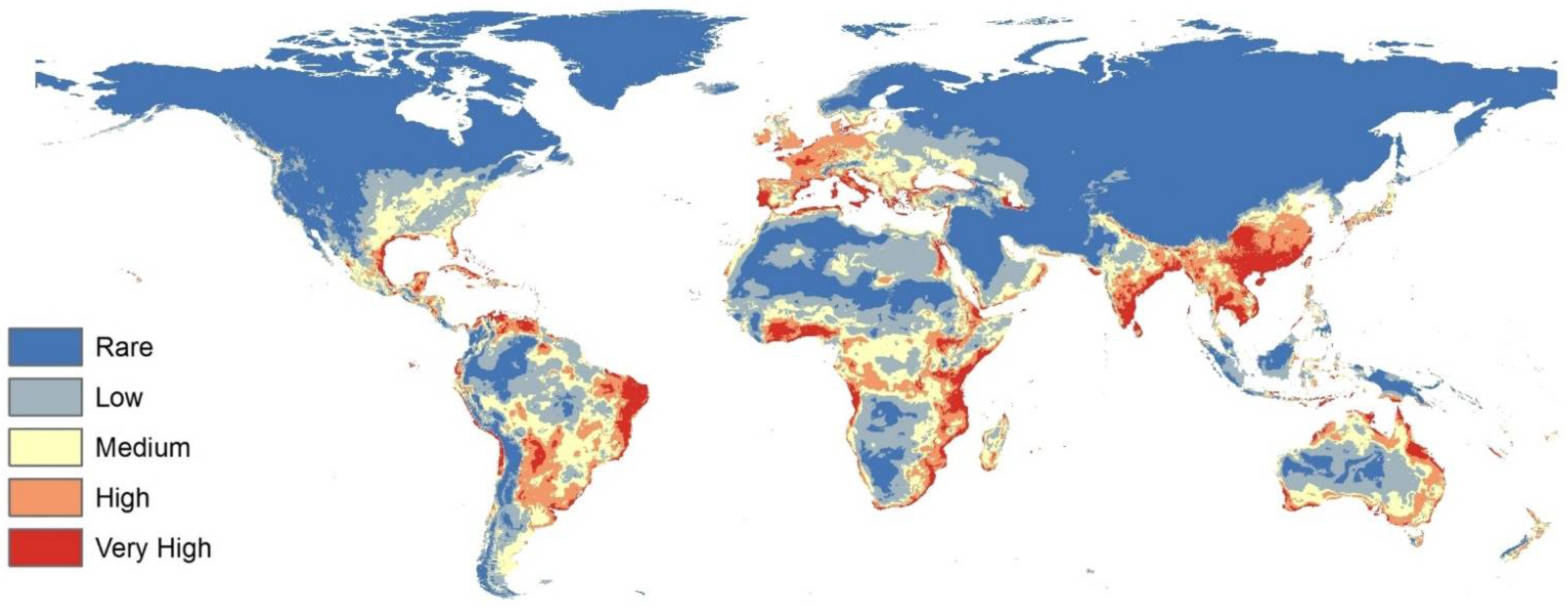
3.3. Predicted Current Potential Distribution of Black Mold
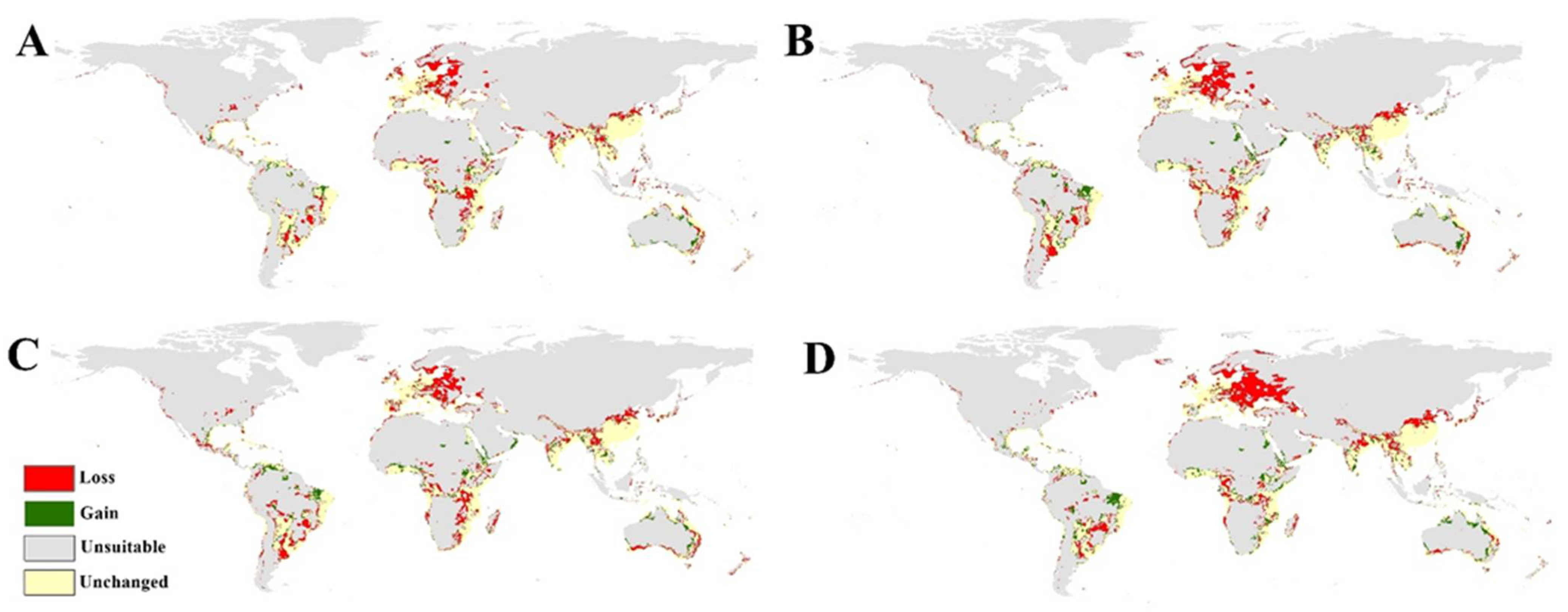
3.4. The Predicted Future Potential Distribution of Black Mold in 2050 and 2070
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, R. Pathogenecity of Aspergillus niger in plants. Cibtech J. Microbiol. 2012, 1, 47–51. [Google Scholar]
- Mateescu, C.; Buruntea, N.; Stancu, N. Investigation of Aspergillus niger growth and activity in a static magnetic flux density field. Rom. Biotechnol. Lett. 2011, 16, 6364–6368. [Google Scholar]
- Shabani, F.; Kumar, L.; Esmaeili, A. A modelling implementation of climate change on biodegradation of low-density polyethylene (LDPE) by Aspergillus Niger in soil undefined. Glob. Ecol. Conserv. 2015, 4, 388–398. [Google Scholar] [CrossRef] [Green Version]
- Ardestani, F.; Kasebkar, R. Non-structured kinetic model of Aspergillus Niger growth and substrate uptake in a batch submerged culture. Br. Biotechnol. J. 2014, 4, 970. [Google Scholar] [CrossRef]
- Person, A.K.; Chudgar, S.M.; Norton, B.L.; Tong, B.C.; Stout, J.E. Aspergillus niger: An unusual cause of invasive pulmonary aspergillosis. J. Med. Microbiol. 2010, 59, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.; Rogers, T.R.; Talento, A.F. COVID-19 associated invasive pulmonary aspergillosis: Diagnostic and therapeutic challenges. J. Fungi 2020, 6, 115. [Google Scholar] [CrossRef]
- Arastehfar, A.; Carvalho, A.; van de Veerdonk, F.L.; Jenks, J.D.; Koehler, P.; Krause, R.; Cornely, O.A.; Perlin, D.S.; Lass-Flörl, C.; Hoenigl, M. COVID-19 associated pulmonary aspergillosis (CAPA)-From immunology to treatment. J. Fungi 2020, 6, 91. [Google Scholar] [CrossRef]
- Koehler, P.; Cornely, O.A.; Böttiger, B.W.; Dusse, F.; Eichenauer, D.A.; Fuchs, F.; Hallek, M.; Jung, N.; Klein, F.; Persigehl, T.; et al. COVID-19 associated pulmonary aspergillosis. Mycoses 2020, 63, 528–534. [Google Scholar] [CrossRef]
- Trovato, L.; Calvo, M.; Migliorisi, G.; Astuto, M.; Oliveri, F.; Oliveri, F. Fatal VAP-related pulmonary aspergillosis by Aspergillus niger in a positive COVID-19 patient. Respir. Med. Case Rep. 2021, 32, 101367. [Google Scholar] [CrossRef]
- Ababutain, I.M. Effect of some ecological factors on the growth of Aspergillus niger and Cladosporium sphaerospermum. Amer. J. Appl. Sci. 2013, 10, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Richard, S.J.T.; Downingd, T.E.; Kuik, O.J.; Smithe, J.B. Distributional aspects of climate change impacts. Glob. Environ. Chang. 2004, 14, 259–272. [Google Scholar]
- Smith, J.B.; Schellnhuber, H.J.; Mirza, M.M.; Fankhauser, S.; Leemans, R.; Erda, L.; Ogallo, L.; Pittock, B.; Richels, R.; Rosenzweig, C.; et al. Vulnerability to climate change and reasons for concern: A synthesis. In Climate Change 2001: Impacts, Adaptation, and Vulnerability; McCarthy, J.J., Canziani, O.F., Leary, N.A., Dokken, D.J., White, K.S., Eds.; Press Syndicate of the University of Cambridge: Cambridge, UK, 2001; pp. 913–967. [Google Scholar]
- IPCC. Climate Change. The Physical Science Basis: Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Hitz, S.; Smith, J. Estimating global impacts from climate change. Glob. Environ. Chang. 2004, 14, 201–218. [Google Scholar] [CrossRef]
- Dillon, M.E.; Wang, G.; Huey, R.B. Global metabolic impacts of recent climate warming. Nature 2010, 467, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.M.; Leadley, P.W.; Proença, V.; Alkemade, R.; Scharlemann, J.P.W.; Fernandez-Manjarrés, J.F.; Araújo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for global biodiversity in the 21st century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef] [Green Version]
- Hosni, E.M.; Nasser, M.G.; Al-Ashaal, S.A.; Rady, M.H.; Kenawy, M.A. Modeling current and future global distribution of Chrysomya bezziana under changing climate. Sci. Rep. 2020, 10, 4947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Liu, G.; Bu, W.; Gao, Y. Ecological niche modeling and its applications in biodiversity conservation. Biodivers. Sci. 2013, 21, 90. [Google Scholar]
- Al Ahmed, A.M.; Naeem, M.; Kheir, S.M.; Sallam, M.F. Ecological distribution modeling of two malaria mosquito vectors using geographical information system in Al-Baha Province, Kingdom of Saudi Arabia. Pak. J. Zool. 2015, 47, 1797–1806. [Google Scholar]
- Naeem, M.; Yuan, X.; Huang, J.; An, J. Habitat suitability for the invasion of Bombus terrestris in East Asian countries: A case study of spatial overlap with local Chinese bumblebees. Sci. Rep. 2018, 8, 11035. [Google Scholar] [CrossRef] [Green Version]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A standard protocol for reporting species distribution models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Maldonado, C.; Molina, C.I.; Zizka, A.; Persson, C.; Taylor, C.M.; Albán, J.; Chilquillo, E.; Rønsted, N.; Antonelli, A. Estimating species diversity and distribution in the era of Big Data: To what extent can we trust public databases? Glob. Ecol. Biogeogr. 2015, 24, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Shabani, F.; Kumar, L.; Esmaeili, A. Future Distributions of Fusarium oxysporum f. spp. in European, middle eastern and north African agricultural regions under climate change. Agric. Ecosyst. Environ. 2014, 197, 96–105. [Google Scholar] [CrossRef]
- Abou-Shaara, H.; Alashaal, S.A.; Hosni, E.M.; Nasser, M.G.; Ansari, M.J.; Alharbi, S.A. Modeling the Invasion of the Large Hive Beetle, Oplostomus fuligineus, into North Africa and South Europe under a Changing Climate. Insects 2021, 12, 275. [Google Scholar] [CrossRef] [PubMed]
- Hosni, E.M.; Nasser, M.; Al-Khalaf, A.A.; Al-Shammery, K.A.; Al-Ashaal, S.; Soliman, D. Invasion of the Land of Samurai: Potential Spread of Old-World Screwworm to Japan under Climate Change. Diversity 2022, 14, 99. [Google Scholar] [CrossRef]
- Hosni, E.M.; Al-Khalaf, A.A.; Nasser, M.G.; Abou-Shaara, H.F.; Radwan, M.H. Modeling the Potential Global Distribution of Honeybee Pest, Galleria mellonella under Changing Climate. Insects 2022, 13, 484. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Ge, X.; Zou, Y.; Zhou, Y.; Wang, T.; Zong, S. Projecting the Potential Global Distribution of Carpomya vesuviana (Diptera: Tephritidae), Considering Climate Change and Irrigation Patterns. Forests 2019, 10, 355. [Google Scholar] [CrossRef]
- Byeon, D.H.; Jung, J.M.; Jung, S.; Lee, W.H. Prediction of global geographic distribution of Metcalfa pruinosa using CLIMEX. Entomol. Res. 2018, 48, 99–107. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 20 March 2020).
- Hosni, E.M.; Al-Khalaf, A.A.; Naguib, R.M.; Afify, A.E.; Abdalgawad, A.A.; Faltas, E.M.; Hassan, M.A.; Mahmoud, M.A.; Naeem, O.M.; Hassan, Y.M.; et al. Evaluation of Climate Change Impacts on the Global Distribution of the Calliphorid Fly Chrysomya albiceps Using GIS. Diversity 2022, 14, 578. [Google Scholar] [CrossRef]
- Kessler, W.H.; Ganser, C.; Glass, G.E. Modeling the Distribution of Medically Important Tick Species in Florida. Insects 2019, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Bari, M.S.; Hossain, M.J.; Akhter, S.; Emran, T.B. Delta variant and black fungal invasion: A bidirectional assault might worsen the massive second/third stream of COVID-19 outbreak in South-Asia. Ethics Med. Public Health 2021, 19, 100722. [Google Scholar] [CrossRef]
- Seyed Alinaghi, S.; Karimi, A.; Barzegary, A.; Pashaei, Z.; Afsahi, A.M.; Alilou, S.; Janfaza, N.; Shojaei, A.; Afroughi, F.; Mohammadi, P.; et al. Mucormycosis infection in patients with COVID-19: A systematic review. Health Sci. Rep. 2022, 5, e529. [Google Scholar]
- Chakraborty, S.; Veleri, S.; Kateriya, S. Mucociliary Respiratory Epithelium Integrity in Molecular Defense and Susceptibility to Pulmonary Viral Infections. Biology 2021, 10, 95. [Google Scholar]
- Ruijter, G.J.G.; Bax, M.; Patel, H.; Flitter, S.J.; van de Vondervoort, P.J.I.; de Vries, R.P.; Vankuyk, P.A.; Visser, J. Mannitol is required for stress tolerance in Aspergillus niger conidiospores. Eukaryot. Cell 2003, 2, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczyk, K.W.; Myszka, L. The effect of temperature on the growth of A. niger in solid state fermentation. Bioprocess Eng. 1994, 10, 123–126. [Google Scholar] [CrossRef]
- Sohail, M.; Siddiqi, R.; Ahmad, A.; Khan, S.A. Cellulase production from Aspergillus niger MS82: Effect of temperature and pH. New Biotechnol. 2009, 25, 437–441. [Google Scholar] [CrossRef]
- Padhi, S.; Masi, M.; Panda, S.K.; Luyten, W.; Cimmino, A.; Tayung, K.; Evidente, A. Antimicrobial secondary metabolites of an endolichenic Aspergillus niger isolated from lichen thallus of Parmotrema ravum. Nat. Prod. Res. 2020, 34, 2573–2580. [Google Scholar] [CrossRef]
- Han, X.; Jiang, H.; Li, F. Dynamic ochratoxin A production by strains of Aspergillus niger intended used in food industry of China. Toxins 2019, 11, 122. [Google Scholar] [CrossRef]
- Arit, H.S.; Jarapa, D.G.; Serrano, H.R. Phytochemical and Anti-Fungal Screening of the Averrhoa bilimbi Fruit Extract against Aspergillus Niger. 8isc Proc. Allied Health 2022, 92–102. [Google Scholar]
- Shyu, D.; Patil, S.; Wilhite, R.; Karle, E.; Beck, P.; Athey, M.; Nelson, T.; Holliday, Z. Invasive pulmonary aspergillosis due to aspergillus niger and COVID-19 pneumonia. In Tp98. Tp098 Fungus among-Us-Rare Fungal Case Reports; American Thoracic Society: New York, NY, USA, 2021; p. A4011. [Google Scholar]
- Fiema, M.; Wlodarczyk, A.; Wojkowska-Mach, J.; Garlicki, J.; Gregorczyk-Maga, I. Atypical Presentation of Aspergillus niger Infection in the Oral Cavity as a Prediction of Invasive Pulmonary Aspergillosis in a Patient with COVID-19: Case Report and Literature Review. Microorganisms 2022, 10, 1630. [Google Scholar] [CrossRef]
- Takazono, T.; Mukae, H.; Izumikawa, K.; Hasegawa, N.; Yokoyama, A. COVID-19 associated pulmonary aspergillosis: A nationwide survey by the Japanese Respiratory Society. ERJ Open Res. 2021, 7, 00402-2021. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioclimatic Variables | Description | Contribution Percentages |
|---|---|---|
| Bio 1 | Annual mean temperature | 36.5% |
| Bio 2 | Mean diurnal range | 13.4% |
| Bio7 | Temperature annual range | 26.6% |
| Bio 12 | Annual precipitation | 17% |
| Bio 14 | Precipitation of driest month | 6.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhalifah, D.H.M.; Damra, E.; Khalaf, S.M.H.; Hozzein, W.N. Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling. Diversity 2022, 14, 845. https://doi.org/10.3390/d14100845
Alkhalifah DHM, Damra E, Khalaf SMH, Hozzein WN. Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling. Diversity. 2022; 14(10):845. https://doi.org/10.3390/d14100845
Chicago/Turabian StyleAlkhalifah, Dalal Hussien M., Eman Damra, Sameh M. H. Khalaf, and Wael N. Hozzein. 2022. "Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling" Diversity 14, no. 10: 845. https://doi.org/10.3390/d14100845
APA StyleAlkhalifah, D. H. M., Damra, E., Khalaf, S. M. H., & Hozzein, W. N. (2022). Biogeography of Black Mold Aspergillus niger: Global Situation and Future Perspective under Several Climate Change Scenarios Using MaxEnt Modeling. Diversity, 14(10), 845. https://doi.org/10.3390/d14100845