

A New Genus and Two New Species of Fireflies from South America (Lampyridae: Lampyrinae: Photinini) †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphology, Terminology, and Illustrations
2.2. Taxon Sampling
2.3. Phylogenetic Analysis
3. Results
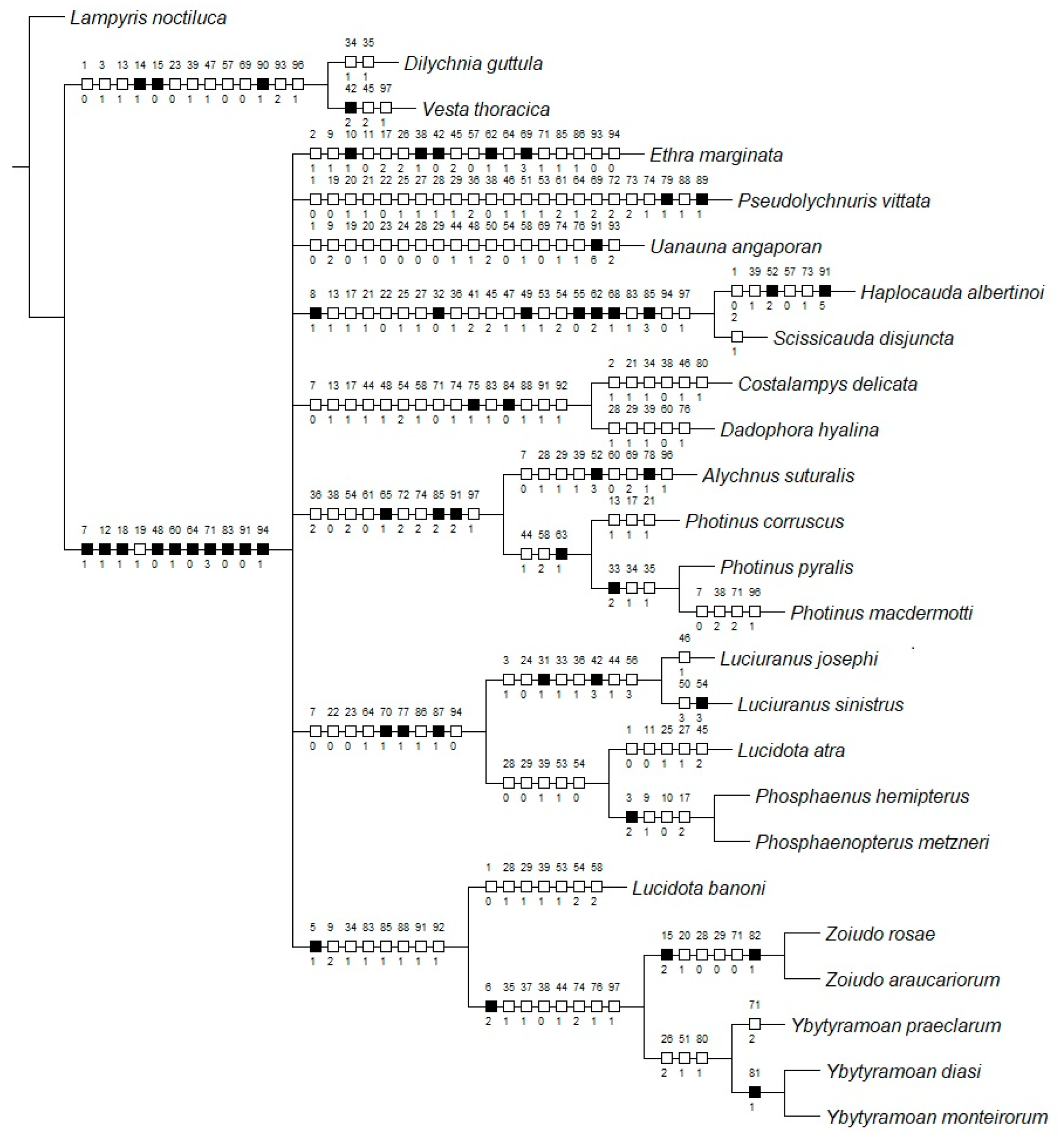
3.1. Characters
- Antenna, antennomeres III-IX, core, shape: (0) serrate; (1) cylindrical (Figure 4F). L = 6; CI = 16; RI = 16.
- Antenna, antennomeres III-IX, single lamellae: (0) absent (Figure 4F); (1) present (branch longer than core antennomere). L = 3; CI = 33; RI = 0.
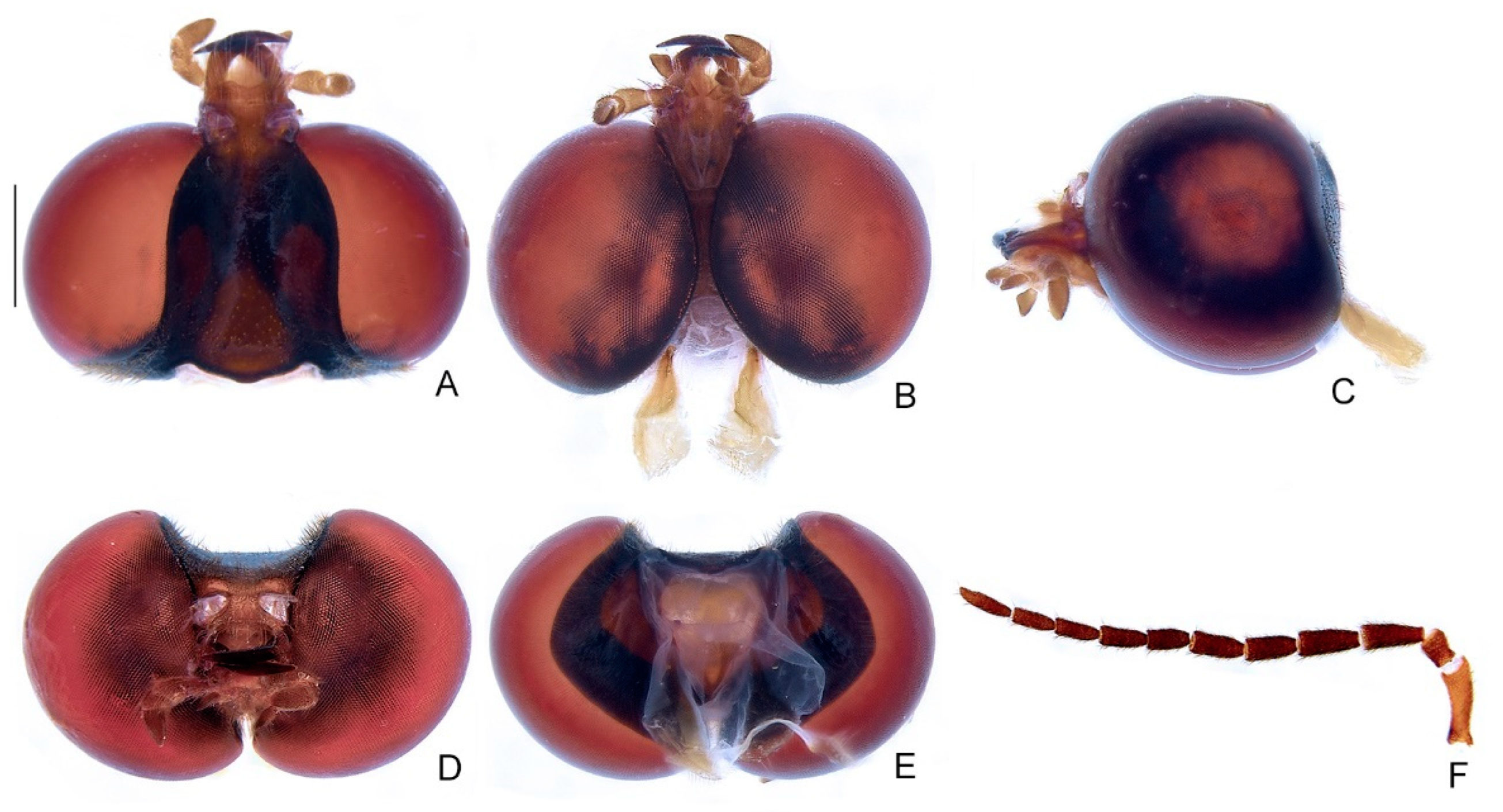
- Clypeus, connection to frons: (0) connected by membrane throughout (Figure 4D); (1) completely obliterate; (2) connate by median third. L = 3; CI = 66; RI = 75.
- Mandibles, orientation in frontal view: (0) overlapping (Figure 4D); (1) crossed (Supplementary Material Figure S4A); (2) convergent (Supplementary Material Figure S4C). L = 2; CI = 100; RI = 100.
- Mandible, apex, shape: (0) sharp (Figure 4A); (1) blunt (Supplementary Material S4B); (2) rounded (Supplementary Material S4C). L = 3; CI = 66; RI = 80.
- Mandible, curvature: (0) straight (Supplementary Material Figure S4C); (1) evenly curved (Supplementary Material S4B); (2) almost right-angled (basal half straight, Figure 4C). L = 2; CI = 100; RI = 100.
- Labrum, sclerite, anterior margin, shape: (0) straight; (1) emarginate (Figure 4A). L = 5; CI = 20; RI = 63.
- Labium, submentum, anterior margin, shape: (0) straight (Figure 4B); (1) notched. L = 1; CI = 100; RI = 100.
- Labium, submentum, lateral margins, shape: (0) subparallel to slightly convergent posteriorly; (1) abruptly constrained posteriorly; (2) strongly convergent posteriorly (Figure 4B). L = 8; CI = 25; RI = 45.
- Labium, palp, palpomere III, lateral margins, shape: (0) obconical; (1) subparallel; (2) divergent apically (Figure 4B). L = 3; CI = 66; RI = 50.
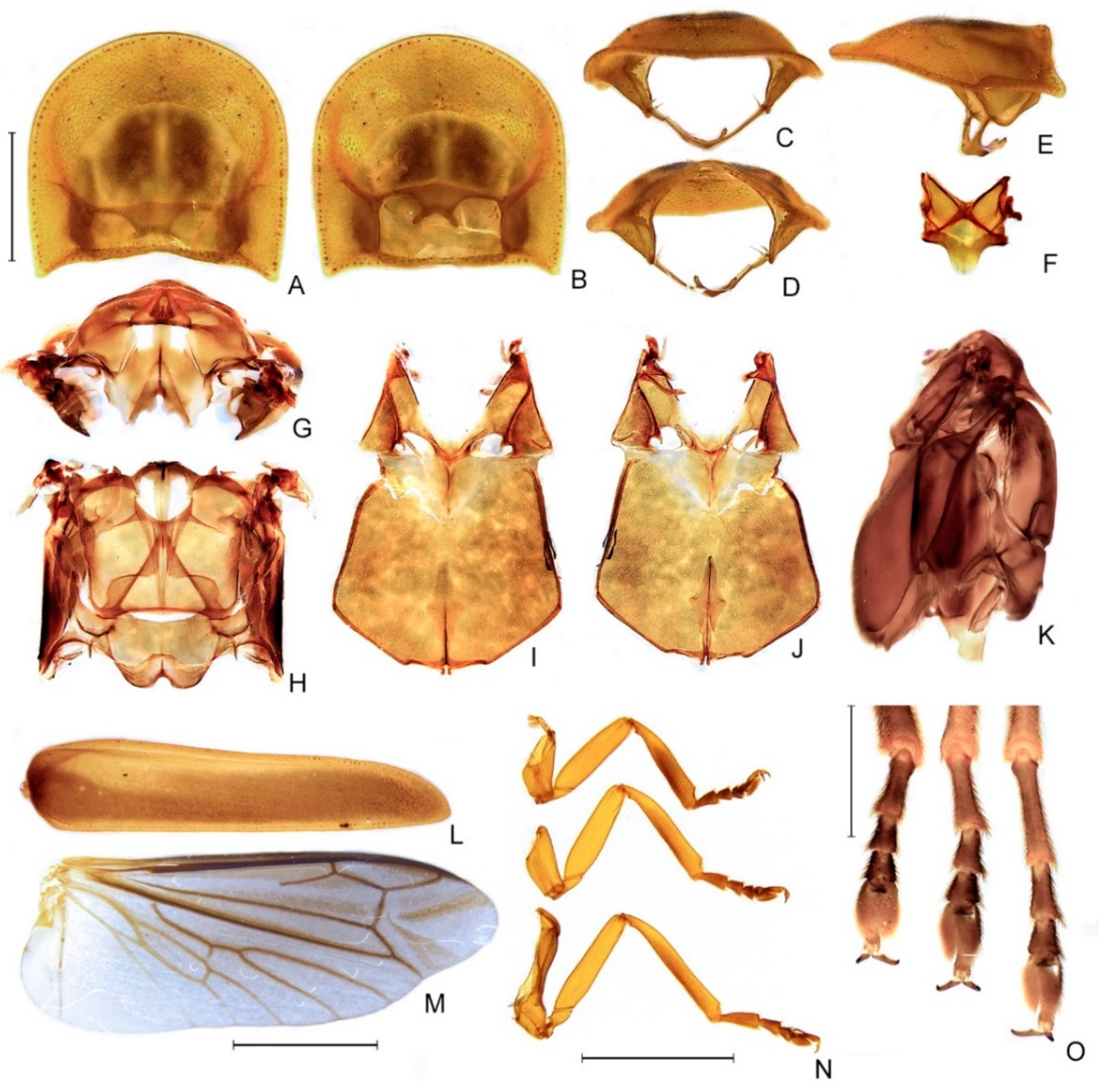
- Pronotum, anterior margin, shape: (0) acuminate anteriorly; (1) evenly rounded (Figure 5B). L = 4; CI = 25; RI = 40.
- Pronotum (lateral view), anterior expansion, curvature: (0) curved upwards; (1) straight (Figure 5E). L = 1; CI = 100; RI = 100.
- Pronotum, lateral margin, length relative to disc: (0) less than a third (Figure 5E); (1) nearly half; (2) at least 1. L = 4; CI = 25; RI = 50.
- Pronotum, disc, sagittal depression: (0) absent (Figure 5A); (1) present. L = 1; CI = 100; RI = 100.
- Pronotum, by the disc, posterior margin, shape: (0) strongly sinuose; (1) almost straight; (2) slightly emarginate (Figure 5A). L = 2; CI = 100; RI = 100.
- Pronotum, posterior corner, notch, presence: (0) absent (Figure 5A); (1) present. L = 5; CI = 20; RI = 50.
- Hypomeron (lateral view), ratio between hypomeron depth and pronotal lateral expansion width: (0) about as long (Figure 5E); (1) at least a 1/5 shorter; (2) at least a 1/5 longer. L = 5; CI = 40; RI = 50.
- Hypomeron (ventral view), area anterior to prosternal insertior, shape: (0) projecting outwards; (1) straight (Figure 5B). L = 1; CI = 100; RI = 100.
- Prosternum, anterior margin, shape: (0) medially sinuose; (1) straight (Figure 5B). L = 5; CI = 20; RI = 50.
- Mesoscutellum, posterior margin, shape: (0) rounded; (1) truncate (Figure 5F). L = 5; CI = 20; RI = 42.
- Elytron, outer margin, shape: (0) straight; (1) rounded; (2) convergent posteriorly (Figure 5L); (3) widest at apical third. L = 7; CI = 42; RI = 50.
- Wing, position of MP3+4 split, relative to CuA1: (0) more basal; (1) more apical (Figure 5M). L = 4; CI = 25; RI = 57.
- Wing, AA3, shape: (0) short (almost as long as wide) and almost perpendicular to AA4; (1) elongate and with an acute angle to AA4 (Figure 5M). L = 3; CI = 33; RI = 60.
- Wing, r3: (0) absent; (1) present (Figure 5M). L = 2; CI = 50; RI = 50.
- Proleg, anterior claw, tooth: (0) absent (Figure 5N); (1) present. L = 3; CI = 33; RI = 33.
- Proleg, tibial spurs, count: (0) zero (Figure 5N); (1) one; (2) two. L = 9; CI = 22; RI = 46.
- Mesoleg, anterior claw, tooth: (0) absent (Figure 5N); (1) present. L = 3; CI = 33; RI = 33.
- Mesoleg, tibial spurs, count: (0) zero (Figure 5N); (1) one; (2) two. L = 7; CI = 28; RI = 37.
- Metaleg, tibial spurs, count: (0) zero (Figure 5N); (1) one; (2) two. L = 7; CI = 28; RI = 37.
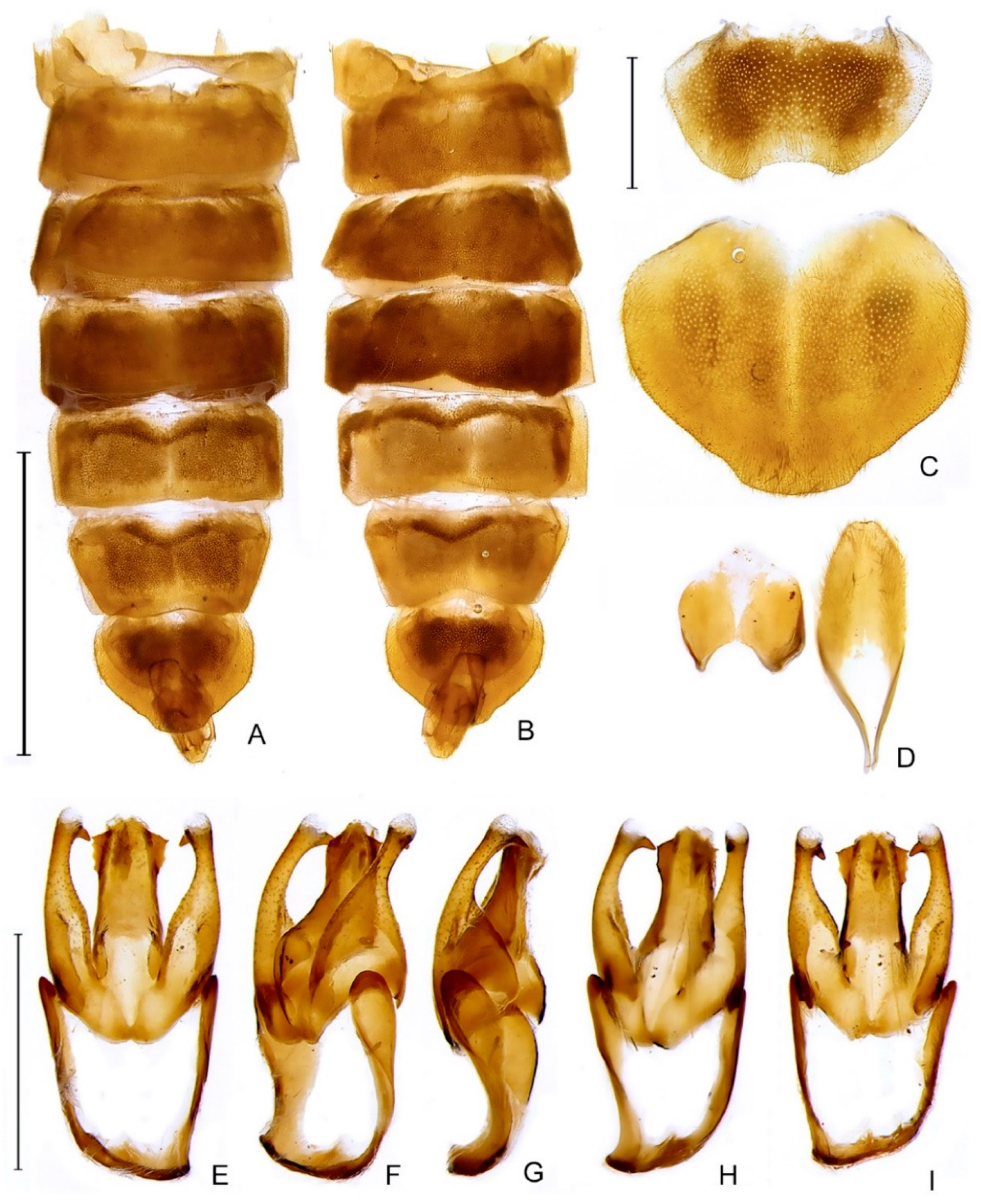
- Tergum I, laterotergite, shape: (0) triangular; (1) quadrangular (Figure 6A). L = 1; CI = 100; RI = 100.
- Tergum I, spiracle, shape: (0) reniform (Figure 6A); (1) subcircular. L = 1; CI = 100; RI = 100.
- Tergum VII, posterior angles, shape: (0) projected, embracing anterior angles of pygidium; (1) rudimentary, slightly projected backwards (Figure 6A). L = 1; CI = 100; RI = 100.
- Sterna II-VIII, width variation: (0) progressively narrow (Figure 6B); (1) widest by sterna III-IV; (2) widest by sternum V. L = 3; CI = 66; RI = 66.
- Sternum VI, lantern: (0) absent; (1) present (Figure 6B). L = 5; CI = 20; RI = 55.
- Sternum VII, lantern: (0) absent; (1) present (Figure 6B). L = 3; CI = 33; RI = 71.
- Sternum VIII, length relative to VII: (0) as long as (Figure 6B); (1) slightly longer; (2) at least a fifth shorter; (3) 2x as long; (4) at least 3x longer. L = 4; CI = 50; RI = 71.
- Sternum VIII, lateral margins, shape: (0) rounded; (1) divergent up to basal 1/4, then convergent posteriorly (Figure 6B). L = 2; CI = 50; RI = 80.
- Sternum VIII, posterior margin, shape: (0) almost straight (Figure 6B); (1) sinuose; (2) emarginate; (4) emarginate with a tiny mucronate projection. L = 6; CI = 33; RI = 55.
- Sternum VIII, posterior margin, median projection: (0) absent (Figure 6B); (1) present. L = 6; CI = 16; RI = 37.
- Sternum VIII, posterior margin, median projection, shape: (0) tiny; (1) elongate; (2) wide, triangular. L = 2; CI = 100; RI = 100.
- Pygidium, relation between length and width: (0) at least a fifth wider than long; (1) as long as wide (Figure 6C); (2) at least a fifth longer than wide. L = 6; CI = 33; RI = 66.
- Pygidium, lateral margins, shape: (0) subparallel; (1) rounded (Figure 6C); (2) divergent posteriorly; (3) convergent posteriorly. L = 3; CI = 100; RI = 100.
- Pygidium, posterior margin, central third, shape: (0) almost straight; (1) rounded (Figure 6C); (2) emarginate; (3) medially notched. L = 6; CI = 50; RI = 25.
- Pygidium, posterolateral corners, degree of development: (0) well-developed; (1) barely conspicuous (Figure 6C). L = 6; CI = 16; RI = 50.
- Pygidium, posterolateral corners, length relative to central third: (0) shorter (Figure 6C); (1) as long as; (2) longer. L = 9; CI = 22; RI = 12.
- Syntergite, shape (proportion): (0) longer than wide (Figure 6D); (1) wider than long. L = 4; CI = 25; RI = 0.
- Syntergite, lateral margin, shape: (0) convergent posteriorly (Figure 6D); (1) subparallel. L = 2; CI = 50; RI = 66.
- Syntergite, anterior margin, shape: (0) mildly emarginated (Figure 6D); (1) strongly indented; (2) almost straight. L = 3; CI = 66; RI = 75.
- Syntergite, pattern of sclerotization: (0) evenly sclerotized (Figure 6D); (1) completely divided by a membranous line. L = 1; CI = 100; RI = 100.
- Syntergite, length relative to sternum IX: (0) 1/3; (1) 1/2 (Figure 6D); (2) 2/3; (3) 1/5. L = 7; CI = 42; RI = 20.
- Syntergite, posterolateral corners, chaetotaxy: (0) glabrous; (1) covered in setae; (2) with dome-shaped sensillae (Figure 6D). L = 8; CI = 25; RI = 53.
- Sternum IX, lateral rods, shape: (0) biconcave (Figure 6D); (1) evenly convergent (Figure 6D); (2) abruptly convergent. L = 2; CI = 100; RI = 100.
- Sternum IX, lateral rods, tips, connection: (0) separated (Figure 6D); (1) fused. L = 5; CI = 20; RI = 42. L = 4; CI = 25; RI = 57.
- Sternum IX, length relative to aedeagus (including phallobase): (0) slightly shorter; (1) slightly longer (Figure 6D); (2) 1/3 longer. L = 6; CI = 50; RI = 72.
- Sternum IX, position relative to VIII: (0) completely covered; (1) partially exposed (Figure 6D). L = 1; CI = 100; RI = 100.
- Sternum IX, posterior half, degree of excavation: (0) evenly sclerotized (Figure 6D); (1) completely membranous; (2) emarginated; (3) deeply clefted (to at least a fifth sternum length). L = 2; CI = 50; RI = 50.
- Phallobase, bilateral symmetry: (0) symmetrical; (1) asymmetrical (Figure 6E). L = 3; CI = 33; RI = 33.
- Phallobase, length relative to phallus: (0) at least a fourth shorter (Figure 6E); (1) as long as; (2) at least a fourth longer. L = 6; CI = 33; RI = 42.
- Phallobase, sagittal line: (0) absent (Figure 6E); (1) present. L = 5; CI = 20; RI = 60.
- Phallobase, sagittal line, extension: (0) throughout phallobase; (1) not reaching apical margin. L = 3; CI = 33; RI = 50.
- Phallobase, apical margin, shape: (0) slightly emarginate; (1) deeply emarginate (C-shaped, Figure 6E–I); (2) medially clefted. L = 8; CI = 25; RI = 33.
- Phallus, dorsal plate, median connection to parameres: (0) connected by membrane (Figure 6E–I); (1) connate; (2) fused. L = 2; CI = 100; RI = 100.
- Phallus, dorsal plate, base, pattern of sclerotization: (0) evenly sclerotized (Figure 6E–I); (1) widely membranous. L = 1; CI = 100; RI = 100.
- Phallus, struts, condition: (0) absent (Figure 6E–I); (1) present (visible through the phallobase). L = 2; CI = 50; RI = 88.
- Phallus, dorsal plate, ventrobasal processes, presence: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, ventrobasal processes, shape: (0) wider than long; (1) globose. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, ventrobasal processes, shape (in apical view): (0) divergent; (1) convergent. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, subcleft transverse groove: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, length relative to parameres: (0) nearly a fifth longer; (1) at least a fifth shorter; (2) as long as (Figure 6E–I); (3) twice as long. L = 7; CI = 42; RI = 50.
- Phallus, dorsal plate, condition: (0) entire (Figure 6E–I); (1) medially split. L = 1; CI = 100; RI = 100.
- Phallus (dorsal view), dorsal plate, anterior margin, shape: (0) rounded (Figure 6E–I); (1) pointed; (2) truncate; (3) clefted. L = 5; CI = 60; RI = 71.
- Phallus, dorsal plate, degree of medial indentation: (0) nearly a 1/3 plate length; (1) nearly 1/2 plate length; (2) nearly 2/3 plate length; (3) completely divided; (4) slightly emarginate. L = 2; CI = 50; RI = 75.
- Phallus, dorsal plate, apical arms (of indented Phallus), shape: (0) widely distanced and slightly convergent; (1) contiguous; (2) fused; (3) apically divergent. L = 7; CI = 42; RI = 20.
- Phallus (lateral view), dorsal plate, overall shape: (0) straight; (1) bent dorsally; (2) slightly curved ventrally (Figure 6E–I); (3) sinuose; (4) with a basal acute angle. L = 6; CI = 50; RI = 72.
- Phallus, dorsal plate, basal abrupt constriction: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, subapical outer teeth: (0) absent; (1) present (Figure 6E–I). L = 3; CI = 33; RI = 66.
- Phallus, dorsal plate, basal joint: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, sides, texture: (0) smooth (Figure 6E–I); (1) toothed. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, longitudinal window: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, subapical keel: (0) absent (Figure 6E–I); (1) present. L = 2; CI = 50; RI = 66.
- Phallus, dorsal plate, subapical keel, extent: (0) entire; (1) medially interrupted. L = 1; CI = 100; RI = 100.
- Phallus, dorsal plate, lateral keel: (0) absent; (1) present (Figure 6E–I). L = 1; CI = 100; RI = 100.
- Phallus, ventral plate: (0) absent; (1) present (Figure 6E–I). L = 4; CI = 25; RI = 72.
- Phallus, ventral plate, length relative to dorsal plate: (0) half as long; (1) as long as or slightly longer; (2) a third shorter; (3) rudimentary (less than a fifth) (Figure 6E–I). L = 4; CI = 75; RI = 80.
- Phallus, endossac opening relative to the apex of dorsal plate: (0) at it; (1) a 1/5 beyond it (Figure 6E–I). L = 5; CI = 60; RI = 80.
- Phallus, endossac, opening, shape: (0) cylindrical (Figure 6E–I); (1) cul-de-sac. L = 1; CI = 100; RI = 100.
- Base of paramere, basal projection: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Paramere, subapical ventral spike, presence: (0) absent; (1) present (Figure 6E–I). L = 4; CI = 25; RI = 62.
- Paramere, subapical ventral spike, shape: (0) pointy (Figure 6E–I); (1) elongate. L = 1; CI = 100; RI = 100.
- Paramere, subapical membranous appendage: (0) absent (Figure 6E–I); (1) present. L = 1; CI = 100; RI = 100.
- Paramere, apex (tip), curvature in lateral view: (0) straight; (1) slightly curved ventrally (Figure 6E–I); (2) evenly curved inwards; (3) curved dorsally; (4) embracing phallus ventrally; (5) sinuose, curved inwards; (6) strongly curved dorsal. L = 6; CI = 66; RI = 80.
- Paramere, apex (tip), sclerotization relative to core paramere: (0) as sclerotized; (1) distinctly membranous (Figure 6E–I); (2) more sclerotized (darker). L = 6; CI = 16; RI = 50.
- Paramere, apex (tip), shape: (0) rounded; (1) blunt (Figure 6E–I); (2) pointed; (3) truncated. L = 4; CI = 50; RI = 71.
- Paramere (lateral view), basal lobe: (0) absent; (1) present (Figure 6E–I). L = 2; CI = 50; RI = 90.
- Paramere (lateral view), basal lobe, shape: (0) robust, rounded (Figure 6E–I); (1) narrow and acute; (2) rudimentary. L = 2; CI = 100; RI = 100.
- Paramere, inner face, shape: (0) smooth (Figure 6E–I); (1) excavate. L = 3; CI = 33; RI = 33.
- Paramere, base, orientation relative to phallus: (0) dorsal; (1) lateral (coplanar) (Figure 6E–I); (2) ventral. L = 4; CI = 25; RI = 72.
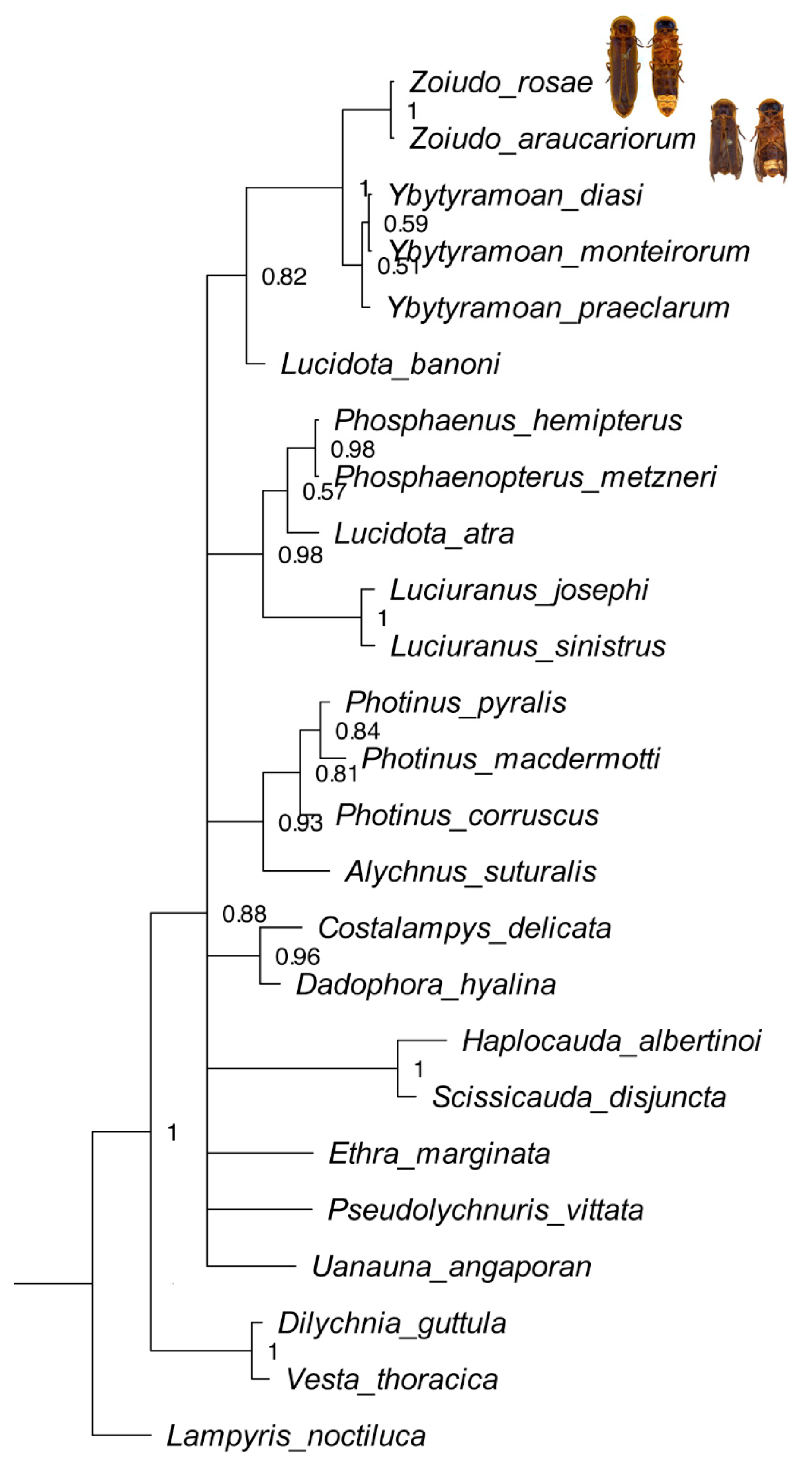
3.2. Phylogenetic Analyses
3.3. Taxonomy
- Elateroidea Leach, 1815
- Lampyridae Rafinesque, 1815
- Lampyrinae Rafinesque, 1815
- Photinini Olivier, 1907
- Zoiudo Roza, Mermudes and Silveira gen. nov.
4. Discussion
4.1. Phylogeny and Classification of Photinini
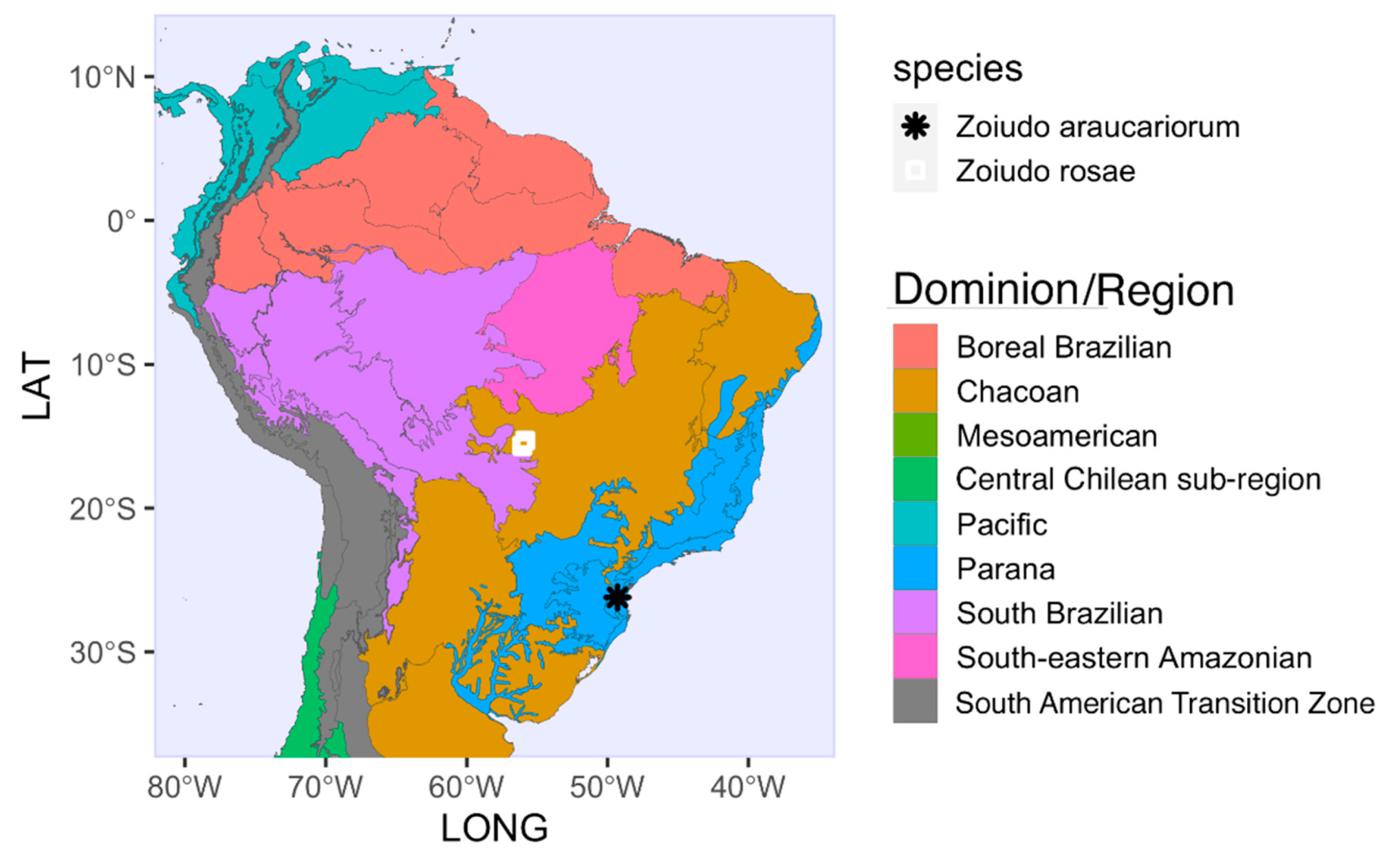
4.2. Hypothesized Habits of Zoiudo
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Branham, M.A.; Wenzel, J.W. The origin of photic behavior and the evolution of sexual communication in fireflies (Coleoptera: Lampyridae). Cladistics 2003, 19, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Vaz, S.; Mermudes, J.R.M.; Paiva, P.C.; Silveira, L.F.L. Systematic review and phylogeny of the firefly genus Dilychnia (Lampyridae: Lampyrinae), with notes on geographical range. Zool. J. Linn. Soc. 2020, 190, 844–888. [Google Scholar] [CrossRef]
- Silveira, L.F.L.; Rosa, S.P.; Mermudes, J.R. Systematic review of the firefly genus Lucernuta Laporte, 1833 (Coleoptera: Lampyridae). Ann. Zool. 2019, 69, 293–314. [Google Scholar] [CrossRef]
- Silveira, L.F.L.; Roza, A.S.; Vaz, S.; Mermudes, J.R.M. Description and phylogenetic analysis of a new firefly genus from the Atlantic Rainforest, with five new species and new combinations (Coleoptera: Lampyridae: Lampyrinae). Arthropod Syst. Phylogeny 2021, 79, 115–120. [Google Scholar] [CrossRef]
- Silveira, L.F.L.; Lima, W.; Fonseca, C.R.V.D.; McHugh, J. Haplocauda, a New Genus of Fireflies Endemic to the Amazon Rainforest (Coleoptera: Lampyridae). Insects 2022, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- McDermott, F.A. The taxonomy of the Lampyridae (Coleoptera). Trans. Am. Entomol. Soc. 1964, 90, 1–72. [Google Scholar]
- Ladino-Peñuela, A.G.; Botero, J.P.; Silveira, L.F.L. First Phylogeny of Pseudolychnuris Reveals Its Polyphyly and a Staggering Case of Convergence at the Andean Paramos (Lampyridae: Lampyrini). Insects 2022, 13, 697. [Google Scholar] [CrossRef]
- Kazantsev, S.V. New firefly taxa from Hispaniola and Puerto Rico (Coleoptera: Lampyridae), with notes on biogeography. Russ. Entomol. J. 2006, 15, 367–392. [Google Scholar] [CrossRef]
- Silveira, L.F.L.; Mermudes, J.R.M. Ybytyramoan, a new genus of fireflies (Coleoptera: Lampyridae, Lampyrinae, Photinini) endemic to the Brazilian Atlantic Rainforest, with description of three new species. Zootaxa 2014, 3835, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Zaragoza-Caballero, S.; Navarrete-Heredia, J.L. Descripción de cuatro especies de Ankonophallus gen. nov. (Coleoptera: Lampyridae: Photinini). Dugesiana 2014, 21, 125–130. [Google Scholar]
- Costa, C. Neotropical Coleoptera, state of knowledge. In Hacia un Proyecto CYTED para el Inventario y Estimación de la Diversidad Entomológica en Iberoamérica: PrIBES 2000. Trabajos del 1er Taller Iberoamericano de Entomología Sistemática, Villa de Leyva, Colombia; Martín-Piera, F., Morrone, J.J., Melic, A., Eds.; Monografías Tercer Milenio: Zaragoza, Mexico, 2000; Volume 1, pp. 99–114. [Google Scholar]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity hotspots; Zachos, F., Habel, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–22. [Google Scholar] [CrossRef]
- Machado, R.B.; Ramos, M.B.N.; Pereira, P.; Caldas, E.; Gonçalves, D.; Santos, N.; Tabor, K.; Steininger, M. Estimativas de Perda da área do Cerrado Brasileiro; Relatório técnico não publicado; Conservação Internacional: Brasília, Brazil, 2004. [Google Scholar]
- Martin, G.J.; Stanger-Hall, K.F.; Branham, M.A.; Silveira, L.F.L.; Lower, S.E.; Hall, D.W.; Li, X.; Lemmon, A.R.; Lemmon, E.M.; Bybee, S.M. (2019). Higher-level phylogeny and reclassification of Lampyridae (Coleoptera: Elateroidea). Insect Syst. Divers. 2019, 3, 11. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Zhou, Y.L.; Lemann, C.; Sinclair, B.; Ślipiński, A. The hind wing of Coleoptera (Insecta): Morphology, nomeclature and phylogenetic significance. Part 1. General discussion and Archostemata–Elateroidea. Ann. Zool. 2021, 71, 421–606. [Google Scholar] [CrossRef]
- Morrone, J.J.; Escalante, T.; Rodríguez-Tapia, G.; Carmona, A.; Arana, M.; Mercado-Gómez, J.D. Biogeographic regionalization of the Neotropical region: New map and shapefile. An. Da Acad. Bras. De Ciências 2022, 94, e20211167. [Google Scholar] [CrossRef]
- Sereno, P.C. Logical basis for morphological characters in phylogenetics. Cladistics 2007, 23, 565–587. [Google Scholar] [CrossRef] [PubMed]
- Maddison, W.P.; Maddison, D.R. Mesquite Version 3.51. A Modular System for Evolutionary Analysis. Available online: http://www.mesquiteproject.org (accessed on 13 September 2022).
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Carpenter, J.M.; Arias, J.S.; Esquivel, D.R.M. Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics 2008, 24, 758–773. [Google Scholar] [CrossRef]
- Marcos Mirande, J. Weighted parsimony phylogeny of the family Characidae (Teleostei: Characiformes). Cladistics 2009, 25, 574–613. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: Enabling high-impact science for phylogenetics researchers with limited resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the extreme to the campus and beyond, Chicago, IL, USA; 2012; pp. 1–8. [Google Scholar]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Lewis, P.O. A likelihood approach to estimating phylogeny from discrete morphological character data. Syst. Biol. 2001, 50, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, K.C. WinClada, Version 1.00. 08. Published by the Author, Ithaca, NY, 734, 745. Available online: http://www.diversityoflife.org/winclada/ (accessed on 13 September 2022).
- Stanger-Hall, K.F.; Lloyd, J.E. Flash signal evolution in Photinus fireflies: Character displacement and signal exploitation in a visual communication system. Evolution 2015, 69, 666–682. [Google Scholar] [CrossRef] [PubMed]
- Stanger-Hall, K.F.; Sander Lower, S.E.; Lindberg, L.; Hopkins, A.; Pallansch, J.; Hall, D.W. The evolution of sexual signal modes and associated sensor morphology in fireflies (Lampyridae, Coleoptera). Proc. R. Soc. B Biol. Sci. 2018, 285, 20172384. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roza, A.S.; Mermudes, J.R.M.; Silveira, L.F.L.d. A New Genus and Two New Species of Fireflies from South America (Lampyridae: Lampyrinae: Photinini). Diversity 2022, 14, 1005. https://doi.org/10.3390/d14111005
Roza AS, Mermudes JRM, Silveira LFLd. A New Genus and Two New Species of Fireflies from South America (Lampyridae: Lampyrinae: Photinini). Diversity. 2022; 14(11):1005. https://doi.org/10.3390/d14111005
Chicago/Turabian StyleRoza, André Silva, José Ricardo Miras Mermudes, and Luiz Felipe Lima da Silveira. 2022. "A New Genus and Two New Species of Fireflies from South America (Lampyridae: Lampyrinae: Photinini)" Diversity 14, no. 11: 1005. https://doi.org/10.3390/d14111005
APA StyleRoza, A. S., Mermudes, J. R. M., & Silveira, L. F. L. d. (2022). A New Genus and Two New Species of Fireflies from South America (Lampyridae: Lampyrinae: Photinini). Diversity, 14(11), 1005. https://doi.org/10.3390/d14111005