

Exploring the Diversity of Elopidae (Teleostei; Elopiformes) Using DNA Barcoding Analysis

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Database Assembly
2.2. Species Delimitation Analysis
2.3. Molecular Analysis
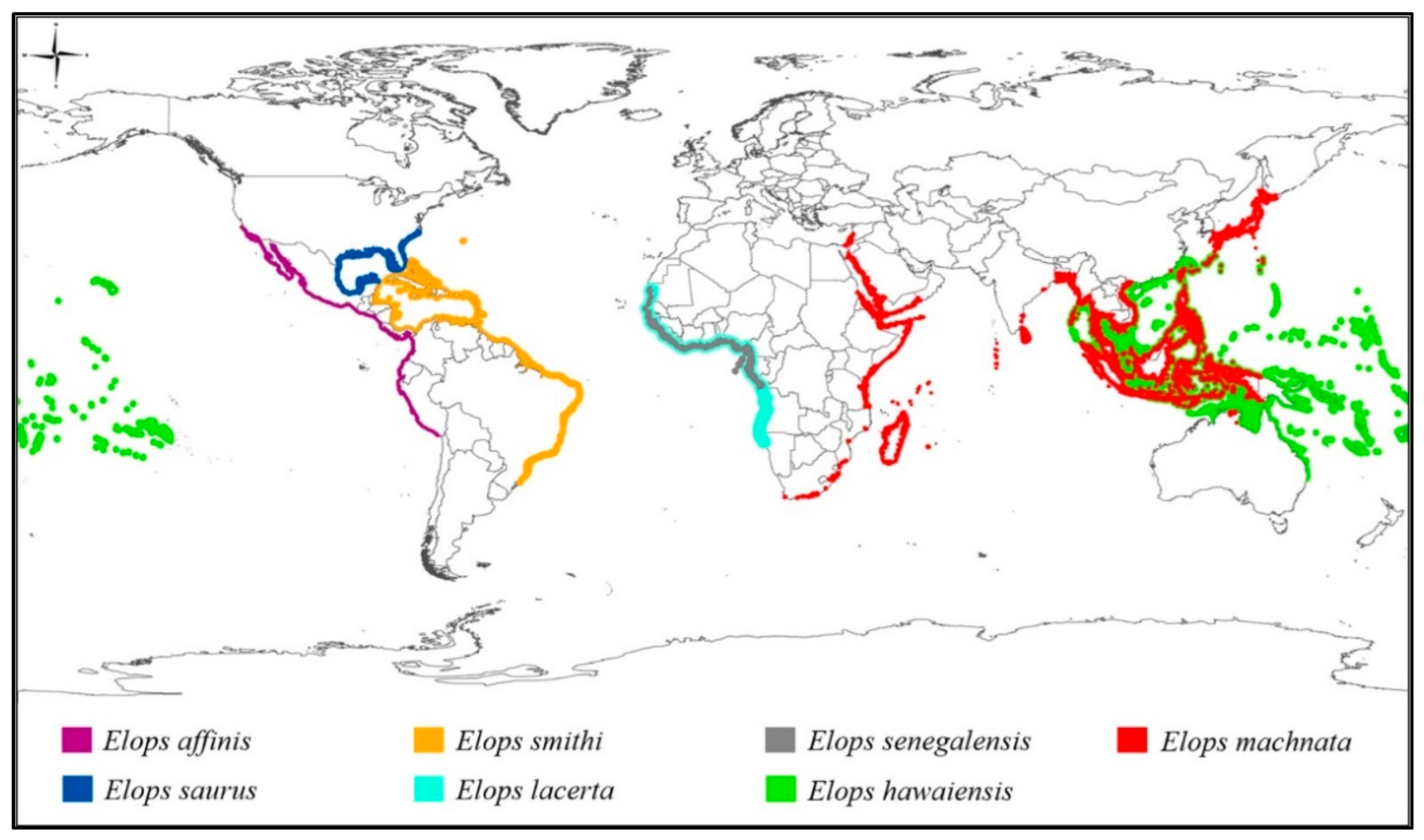
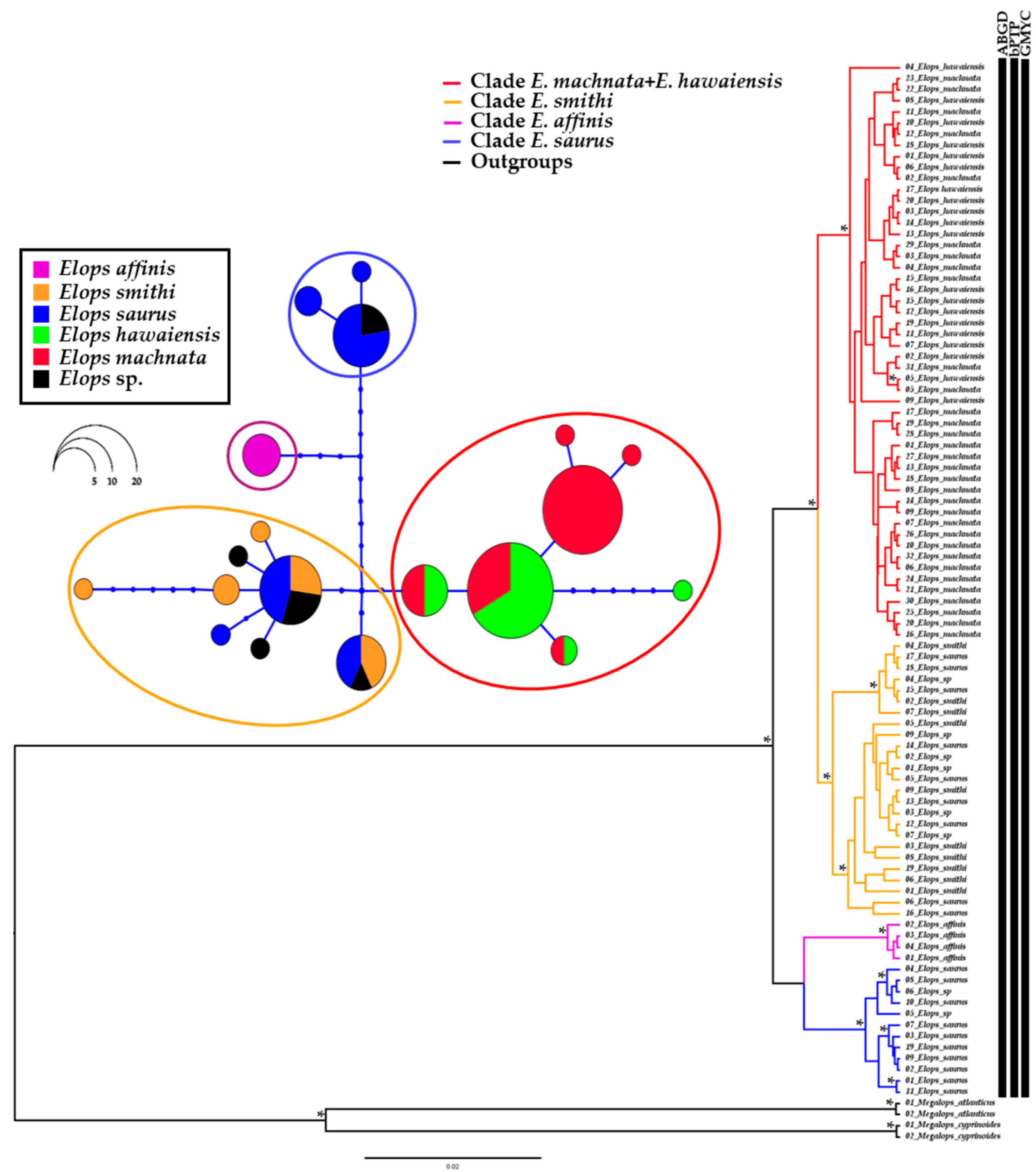
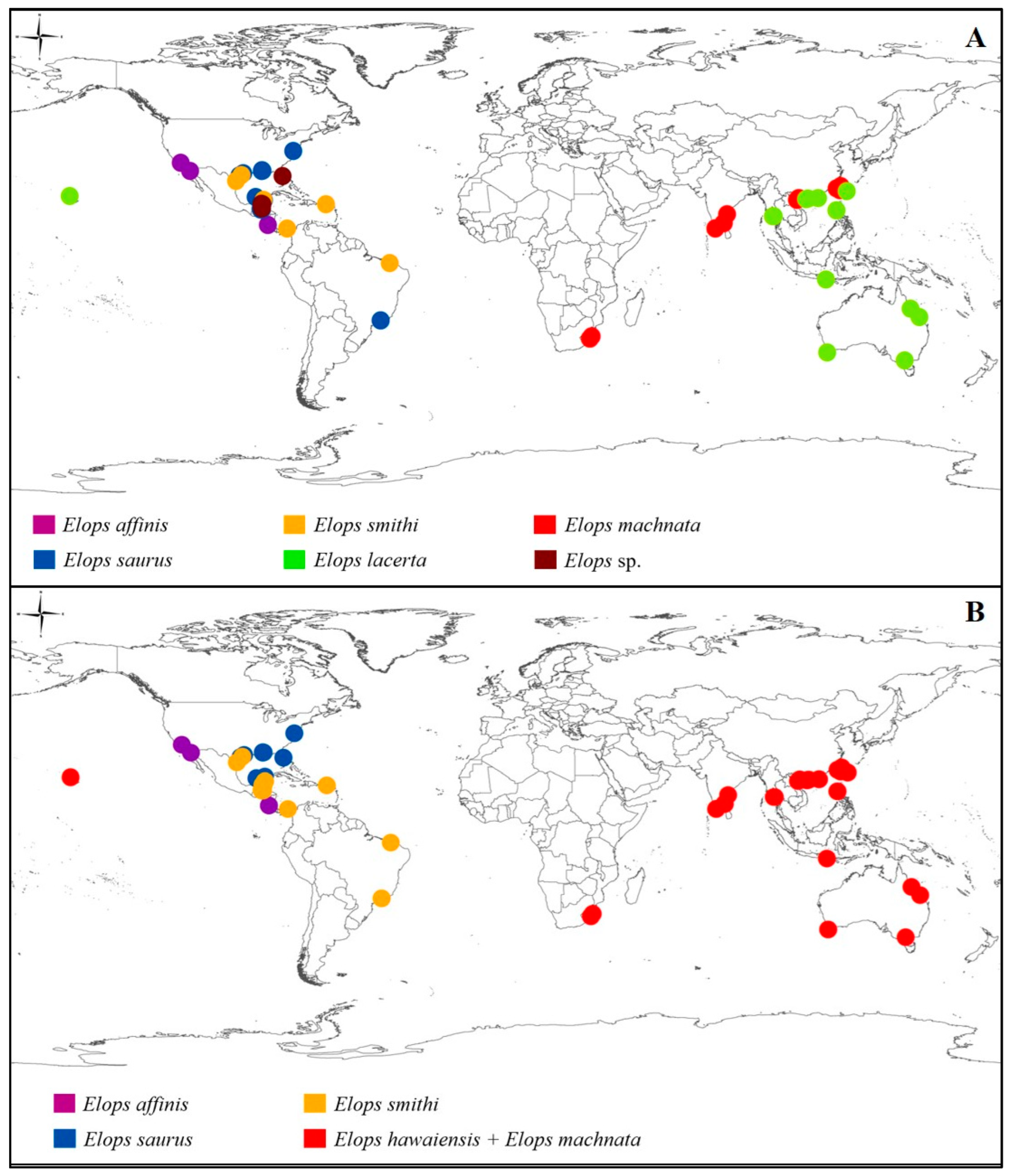
3. Results
4. Discussion
4.1. Low Genetic Diversity Resulted from Recent Radiation
4.2. Taxonomic Conflicts among Elopidae Species
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, A.J.; Horodysky, A.Z.; McBride, R.S.; Guindon, K.; Shenker, J.; MacDonald, T.C.; Harwell, H.D.; Ward, R.; Carpenter, K. Global conservation status and research needs for tarpons (Megalopidae), ladyfishes (Elopidae) and bonefishes (Albulidae). Fish Fish. 2014, 15, 280–311. [Google Scholar] [CrossRef]
- De Sousa, R.P.C.; Silva-Oliveira, G.C.; Furo, I.O.; Oliveira-Filho, A.B.; Brito, C.D.B.; Rabelo, L.; Guimarães-Costa, A.; Oliveira, E.H.C.; Vallinoto, M. The role of the chromosomal rearrangements in the evolution and speciation of Elopiformes fishes (Teleostei; Elopomorpha). Zool. Anz. 2021, 290, 40–48. [Google Scholar] [CrossRef]
- McBride, R.S.; Rocha, C.R.; Ruiz-Carus, R.; Bowen, B.W. A new species of ladyfish, of the genus Elops (Elopiformes: Elopidae), from the western Atlantic Ocean. Zootaxa 2010, 2346, 29–41. [Google Scholar] [CrossRef]
- Ramanadevi, V.; Thangaraj, M. Comparative Phylogenetic Study of Four Genes of Mitochondrial Genome in Tenpounder Fishes (Order: Elopiformes). Not. Sci. Biol. 2013, 5, 282–289. [Google Scholar] [CrossRef]
- Kwun, H.J.; Kang, C.B.; Kim, J.K. Redescription of the Hawaiian ladyfish Elops hawaiensis from Korea. Korean J. Ichthyol. 2011, 23, 80–84. [Google Scholar]
- Whitehead, P.J.P. The species of Elops (Pisces: Elopidae). J. Nat. Hist. 1962, 5, 321–329. [Google Scholar] [CrossRef]
- Regan, C.T. VII—A revision of the fishes of the genus Elops. Ann. Mag. Nat. Hist. 1909, 3, 37–40. [Google Scholar] [CrossRef] [Green Version]
- De Sousa, R.P.C.; Sodre, D.; Costa, R.M.; Vallinoto, M.; Oliveira, E.H.C.; Silva-Oliveira, G.C.; Sampaio, I.; Guimarães-Costa, A. Range distribution and contributions to taxonomy of Elops smithi (Elopiformes: Elopidae). An. Acad. Bras. Ciências 2019, 91, e20181240. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- De Carlos, A.; Bañón, R.; Cobo-Arroyo, S.; Arronte, J.C.; Del Río, J.L.; Barros-García, D. DNA barcoding flags the existence of sympatric cryptic species in the slender codling Halargyreus johnsonii Günther, 1862 (Gadiformes, Moridae). Mar. Biodivers. 2020, 50, 49. [Google Scholar] [CrossRef]
- Da Silva, R.; Peloso, P.L.; Sturaro, M.J.; Veneza, I.; Sampaio, I.; Schneider, H.; Gomes, G. Comparative analyses of species delimitation methods with molecular data in snappers (Perciformes: Lutjaninae). Mitochondrial DNA Part A 2018, 29, 1108–1114. [Google Scholar] [CrossRef]
- Araújo, R.G.; Dda Silva, R.D.F.; Sampaio, I.; Guimarães-Costa, A. Does DNA barcoding offer meaningful insights into the diversity of the parrotfish of the genus Sparisoma (Scaridae). J. Appl. Ichthyol 2019, 35, 1029–1033. [Google Scholar] [CrossRef]
- Ward, R.D. DNA barcode divergence among species and genera of birds and fishes. Mol. Ecol. Resour. 2009, 9, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; de Maio, N.; et al. Beast 2.5: An advanced software platform for bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 2 January 2022).
- Salzburger, W.; Ewing, G.B.; Von-Haeseler, A. The performance of phylogenetic algorithms in estimating haplotype genealogies with migration. Mol. Ecol. 2011, 20, 1952–1963. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubert, N.; Hanner, R. DNA barcoding, species delineation and taxonomy: A historical perspective. DNA Barcodes 2015, 3, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Lara, A.; Ponce de León, J.L.; Rodriguez, R.; Casane, D.; Cote, G.; Bernatchez, L.; García-Machado, E. DNA barcoding of Cuban freshwater fishes: Evidence for cryptic species and taxonomic conflicts. Mol. Ecol. Resour. 2010, 10, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Geiger, M.F.; Herder, F.; Monaghan, M.T.; Almada, V.; Barbieri, R.; Bariche, M.; Berrebi, P.; Bohlen, J.; Casal-Lopez, M.; Delmastro, G.B.; et al. Spatial heterogeneity in the Mediterranean Biodiversity Hotspot affects barcoding accuracy of its freshwater fishes. Mol. Ecol. Resour. 2014, 14, 1210–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bingpeng, X.; Heshan, L.; Zhilan, Z.; Chunguang, W.; Yanguo, W.; Jianjun, W. DNA barcoding for identification of fish species in the Taiwan Strait. PLoS ONE 2018, 13, e0198109. [Google Scholar] [CrossRef]
- Esmaeili, H.R.; Teimori, A.; Zarei, F.; Sayyadzadeh, G. DNA barcoding and species delimitation of the Old World tooth-carps, family Aphaniidae Hoedeman, 1949 (Teleostei: Cyprinodontiformes). PLoS ONE 2020, 15, e0231717. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Filho, L.F.D.S.; Rocha, T.C.D.; Rêgo, P.S.D.; Schneider, H.; Sampaio, I.; Vallinoto, M. Identification and phylogenetic inferences on stocks of sharks affected by the fishing industry off the Northern coast of Brazil. Genet. Mol. Biol. 2009, 32, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D.N. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Desalle, R.; Goldstein, P. Review and interpretation of trends in DNA barcoding. Front. Ecol. Evol. 2019, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.; Guindon, K.; Horodysky, A.; MacDonald, T.; McBride, R.; Shenker, J.; Ward, R. Elops saurus. The IUCN Red List of Threatened Species. 2012. Available online: https://www.iucnredlist.org/species/191822/2006513 (accessed on 27 December 2021).
- Deidun, A.; Zava, B. Far from home… the first documented capture of the genus Elops (Actinopterygii, Elopidae) from the Mediterranean. BioInvasions Rec. 2020, 9, 223–227. [Google Scholar] [CrossRef]
- Hastungs, P.A.; Burton, R.S. Establishing a DNA sequence database for the marine fish Fauna of California. UC San Diego: California Sea Grant College Program. 2008, Volume 1, p. 5. Available online: https://escholarship.org/uc/item/1ck9b3qs (accessed on 27 December 2021).
- Zhang, J.; Hanner, R. DNA barcoding is a useful tool for the identification of marine fishes from Japan. Biochem. Syst. Ecol. 2011, 39, 31–42. [Google Scholar] [CrossRef]
- Zhang, J. Species identification of marine fishes in China with DNA barcoding. Evid. Based. Complement. Altern. Med. 2011, 2011, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Liao, J.; Bai, Q.; Chen, C.; Guo, Y.; Liu, C.; Wang, Z. COI barcode assisted species diversity study on mangrove fish in Leizhou Peninsula. Oceanol. Limnol. Sin. 2016, 47, 663–672. [Google Scholar]
- Ward, R.D.; Holmes, B.H. An analysis of nucleotide and amino acid variability in the barcode region of cytochrome c oxidase I (cox1) in fishes. Mol. Ecol. Notes. 2007, 7, 899–907. [Google Scholar] [CrossRef]
- Inoue, J.G.; Miya, M.; Tsukamoto, K.; Nishida, M. Mitogenomic evidence for the monophyly of elopomorph fishes (Teleostei) and the evolutionary origin of the leptocephalus larva. Mol. Phylogenet. Evol. 2004, 32, 274–286. [Google Scholar] [CrossRef]
- Thu, P.T.; Huang, W.C.; Chou, T.K.; Van Quan, N.; Van Chien, P.; Li, F.; Shao, K.T.; Liao, T.Y. DNA barcoding of coastal ray-finned fishes in Vietnam. PLoS ONE 2019, 14, e0222631. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.; Shao, K.T.; Lin, H.Y.; Chiu, Y.C.; Lee, M.Y.; Liu, S.H.; Lin, P.L. DNA barcodes of the native ray-finned fishes in Taiwan. Mol. Ecol. Resour. 2017, 17, 796–805. [Google Scholar] [CrossRef]
- Steinke, D.; Connell, A.D.; Hebert, P.D. Linking adults and immatures of South African marine fishes. Genome 2016, 59, 959–967. [Google Scholar] [CrossRef] [Green Version]
- April, J.; Mayden, R.L.; Hanner, R.H.; Bernatchez, L. Genetic calibration of species diversity among North America’s freshwater fishes. Proc. Natl. Acad. Sci. USA 2011, 108, 10602–10607. [Google Scholar] [CrossRef] [Green Version]
- Valdez-Moreno, M.; Vásquez-Yeomans, L.; Elías-Gutiérrez, M.; Ivanova, N.V.; Hebert, P.D. Using DNA barcodes to connect adults and early life stages of marine fishes from the Yucatan Peninsula, Mexico: Potential in fisheries management. Mar. Freshw. Res. 2010, 61, 655–671. [Google Scholar] [CrossRef]
- Ribeiro, A.O.; Caires, R.A.; Mariguela, T.C.; Pereira, L.H.G.; Hanner, R.; Oliveira, C. DNA barcodes identify marine fishes of São Paulo State, Brazil. Mol. Ecol. Res. 2012, 12, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Guimarães-Costa, A.J.; Machado, F.S.; Oliveira, R.R.; Silva-Costa, V.; Andrade, M.C.; Giarrizzo, T.; Saint-Paul, U.; Sampaio, I.; Schneider, H. Fish diversity of the largest deltaic formation in the Americas-a description of the fish fauna of the Parnaíba Delta using DNA Barcoding. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weigt, L.A.; Baldwin, C.C.; Driskell, A.; Smith, D.G.; Ormos, A.; Reyier, E.A. Using DNA barcoding to assess Caribbean reef fish biodiversity: Expanding taxonomic and geographic coverage. PLoS ONE 2012, 7, e41059. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Number of Vertebrae | Genetic Distance Cytb | Genetic Distance COI | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Authors | [7] | [8] | [3] | [5] | [3] | [5] | [4] | [8] | [2] | [4] | [2] | |
| Species | ||||||||||||
| E. smithi | − | − | 73–80 | − | 2.3–2.9 | 0.9–2.2 | 1–12.1 | 1.9–2.9 | 1.8–3.3 | − | 2.3–2.6 | |
| E. saurus | 78–79 | 73–80 | 79–87 | − | 2.3–2.9 | 1.7–2.2 | 1.3–12.3 | 1.9–3.2 | 1.8–3.2 | 1.4–2.1 | 1.8–2.6 | |
| E. affinis | 79 | 73–80 | − | − | − | − | 12.0–12.3 | 2.8–3.2 | 3.2–3.3 | 2–2.3 | 1.8–2.3 | |
| E. machnata | 63–64 | 63–64 | 63–64 | 63–64 | − | − | − | − | − | 0.3–2 | − | |
| E. hawaiensis | 68–69 | 68–70 | 68–70 | 65–67 | − | 0.9–2.2 | 1–12.0 | − | − | 0.3–2.3 | − | |
| E. senegalensis | 69 | 63–70 | 67 | − | − | − | − | − | − | − | − | |
| E. lacerta | 74 | 63–70 | − | − | − | − | − | − | − | − | − | |
| Species | 1 | 2 | 3 | 4 | 5 | 6 |
|---|---|---|---|---|---|---|
| 1- E. affinis | 0 | |||||
| 2- E. hawaiensis + E. machnata | 2.11 | 0.25 | ||||
| 3- E. saurus | 1.83 | 2.47 | 0.33 | |||
| 4- E. smithi | 2.32 | 1.29 | 2.78 | 0.69 | ||
| 5- M. atlanticus | 18.74 | 19.41 | 19.31 | 19.35 | 0 | |
| 6- M. cyprinoides | 21.54 | 22.42 | 21.85 | 22.55 | 15.38 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Sousa, R.P.C.; Bessa-Brito, C.D.; Guimarães-Costa, A.; Evangelista-Gomes, G.; Sampaio, I.; de Oliveira, E.H.C.; Vallinoto, M. Exploring the Diversity of Elopidae (Teleostei; Elopiformes) Using DNA Barcoding Analysis. Diversity 2022, 14, 1008. https://doi.org/10.3390/d14111008
de Sousa RPC, Bessa-Brito CD, Guimarães-Costa A, Evangelista-Gomes G, Sampaio I, de Oliveira EHC, Vallinoto M. Exploring the Diversity of Elopidae (Teleostei; Elopiformes) Using DNA Barcoding Analysis. Diversity. 2022; 14(11):1008. https://doi.org/10.3390/d14111008
Chicago/Turabian Stylede Sousa, Rodrigo Petry Corrêa, Carla Denise Bessa-Brito, Auryceia Guimarães-Costa, Grazielle Evangelista-Gomes, Iracilda Sampaio, Edivaldo Herculano Corrêa de Oliveira, and Marcelo Vallinoto. 2022. "Exploring the Diversity of Elopidae (Teleostei; Elopiformes) Using DNA Barcoding Analysis" Diversity 14, no. 11: 1008. https://doi.org/10.3390/d14111008
APA Stylede Sousa, R. P. C., Bessa-Brito, C. D., Guimarães-Costa, A., Evangelista-Gomes, G., Sampaio, I., de Oliveira, E. H. C., & Vallinoto, M. (2022). Exploring the Diversity of Elopidae (Teleostei; Elopiformes) Using DNA Barcoding Analysis. Diversity, 14(11), 1008. https://doi.org/10.3390/d14111008