
Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009 †


Abstract
:1. Introduction
2. Material and Methods
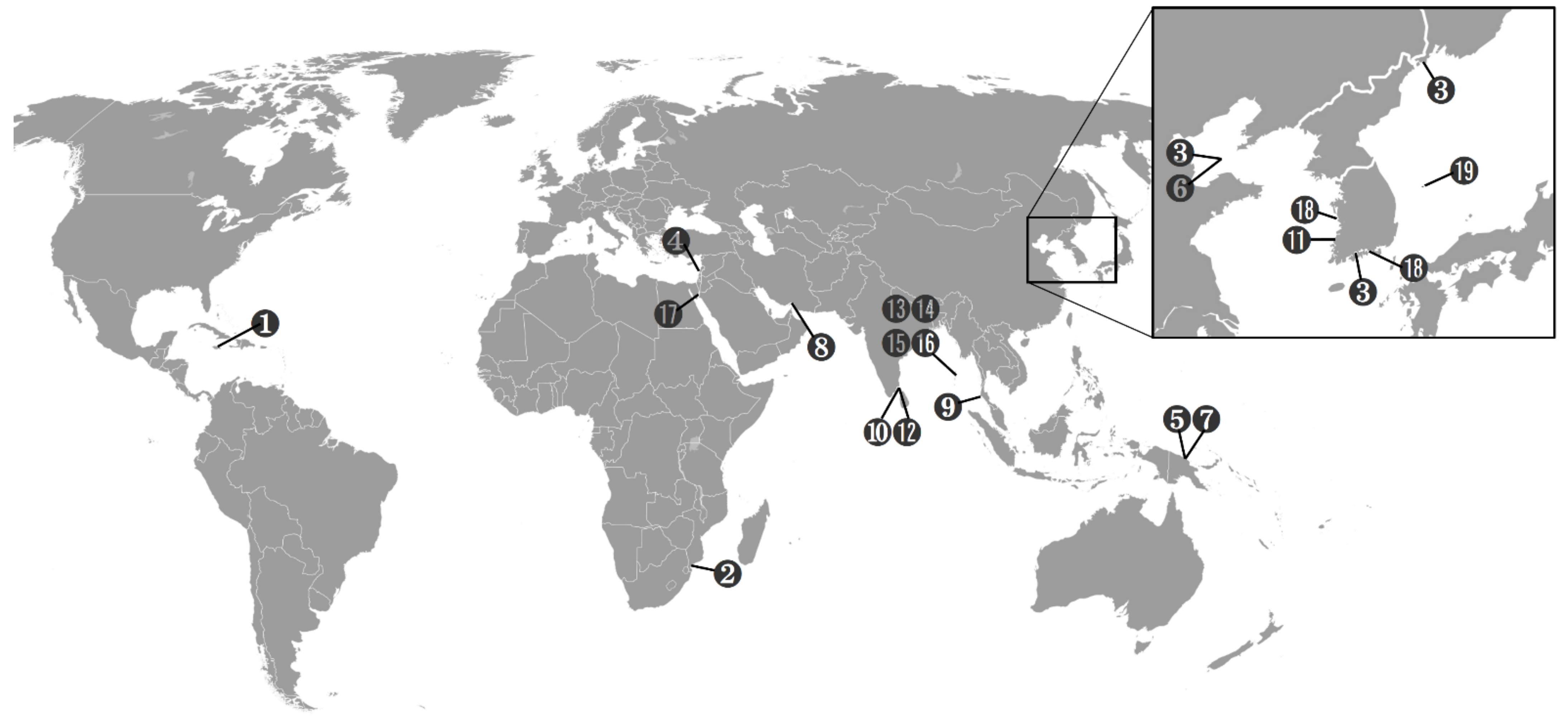
2.1. Sampling Locations and Methods
2.2. DNA Extraction and Sequencing
2.3. Morphological Characterization
3. Results
- Order Polyarthra Lang, 1944
- Family Canuellidae Lang, 1944
- Genus Scottolana Huys, 2009
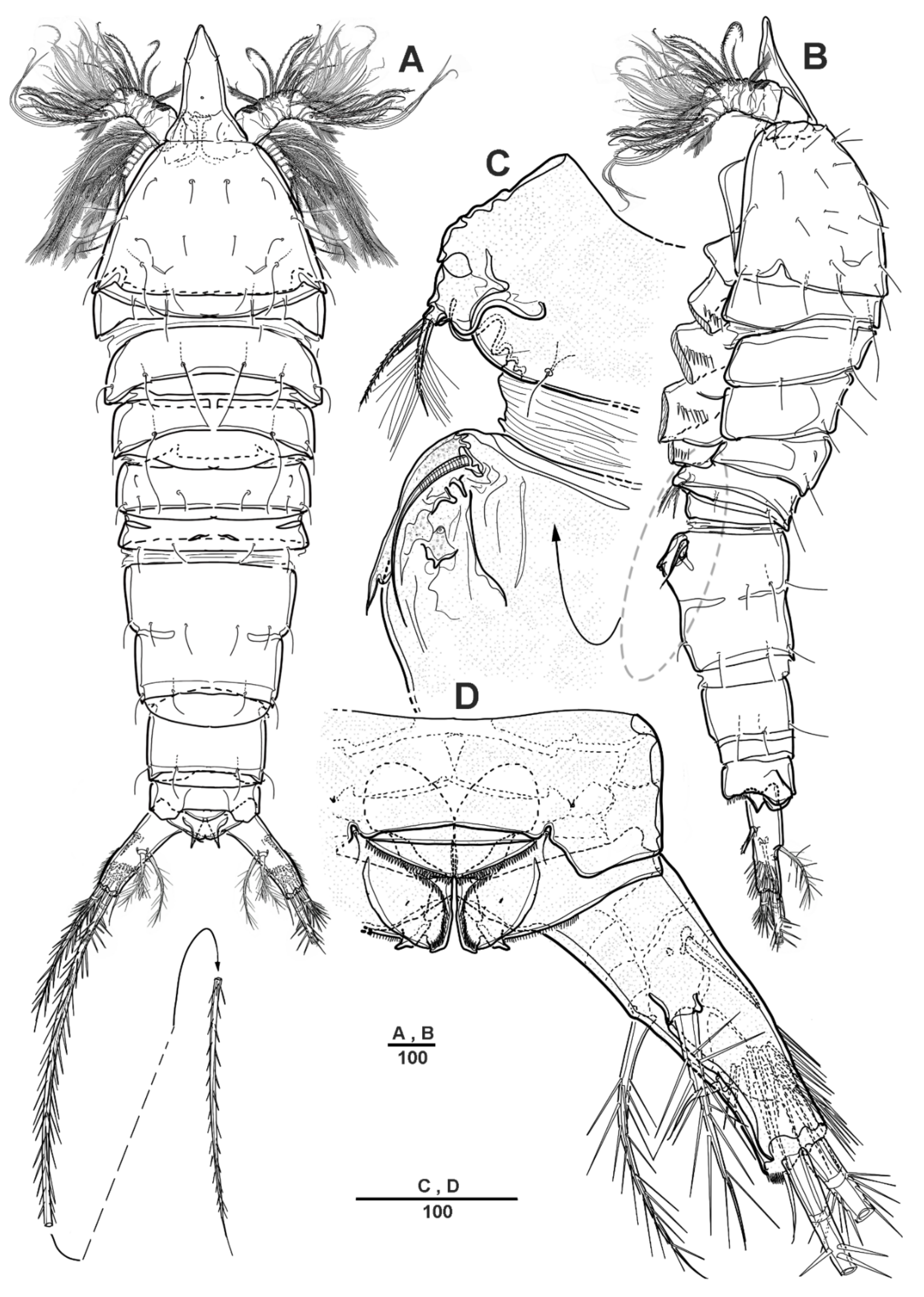
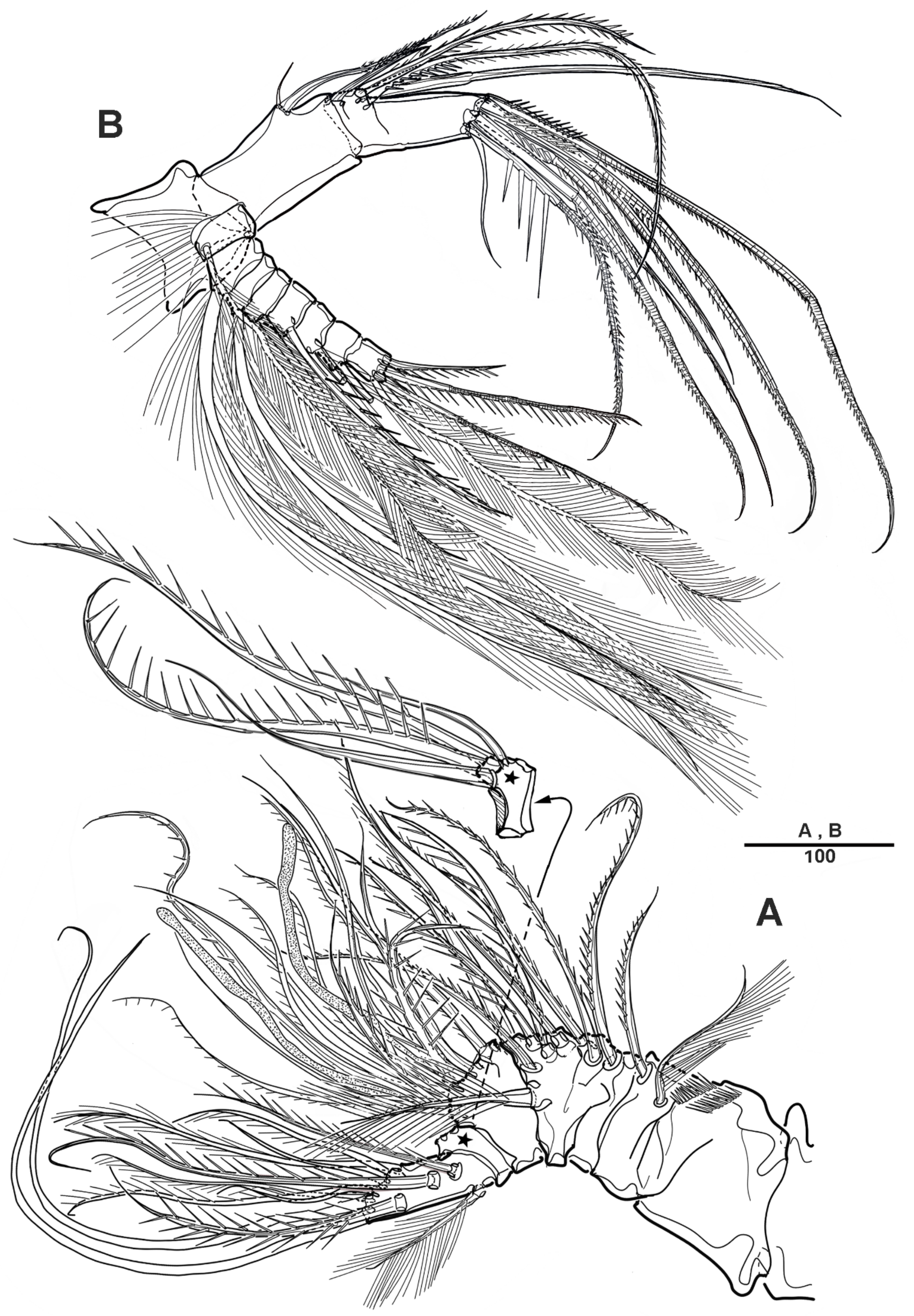
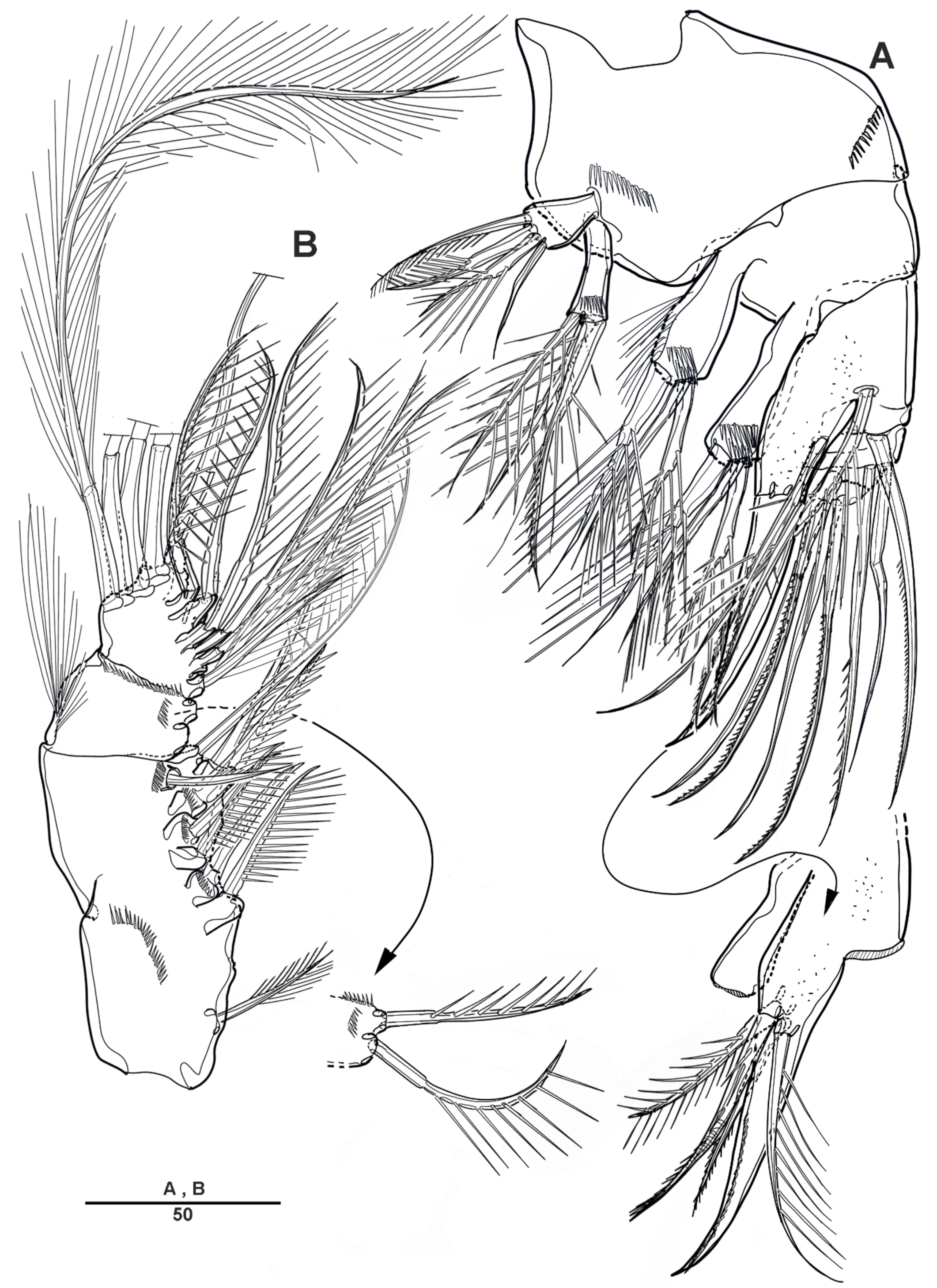
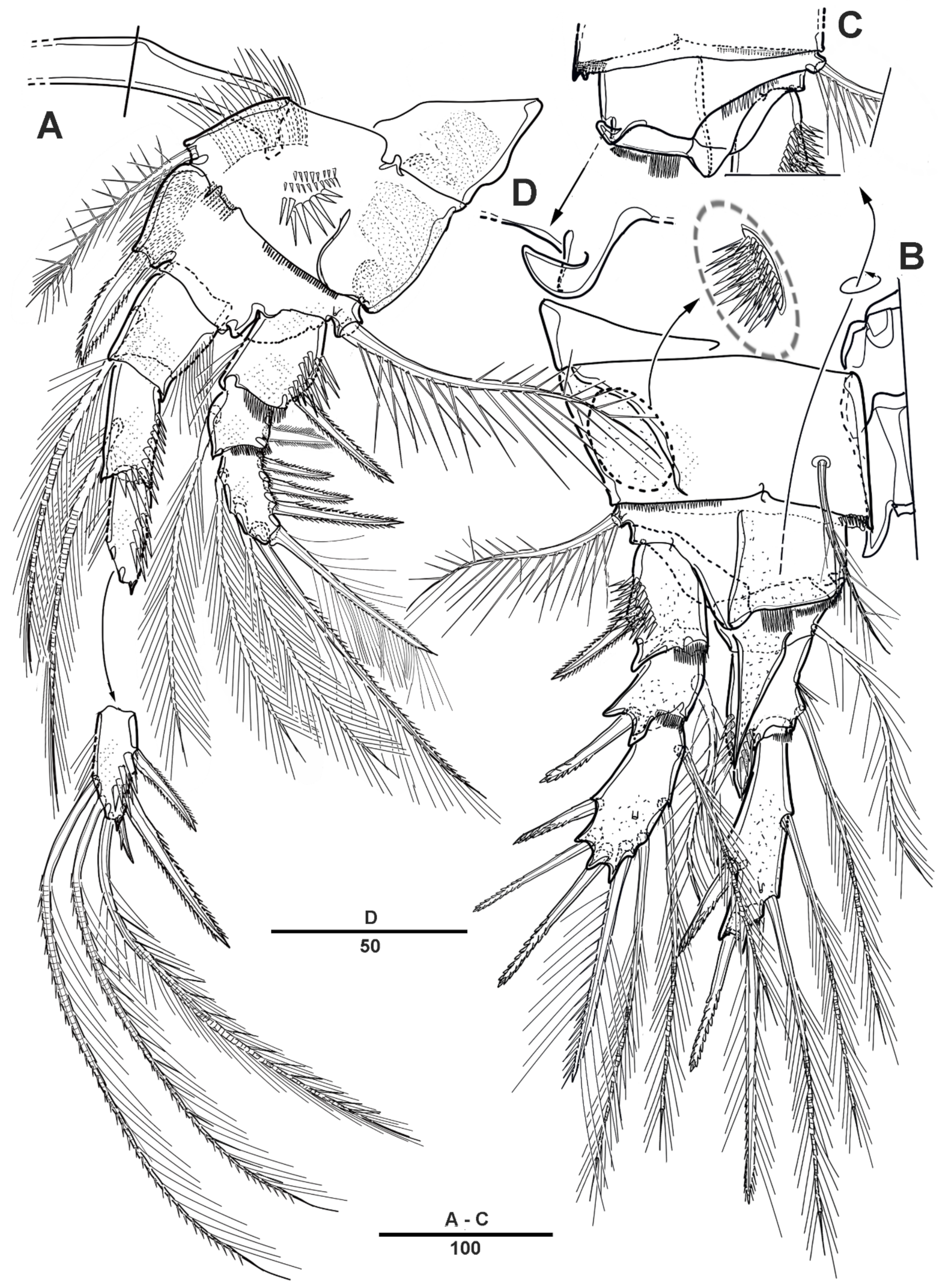
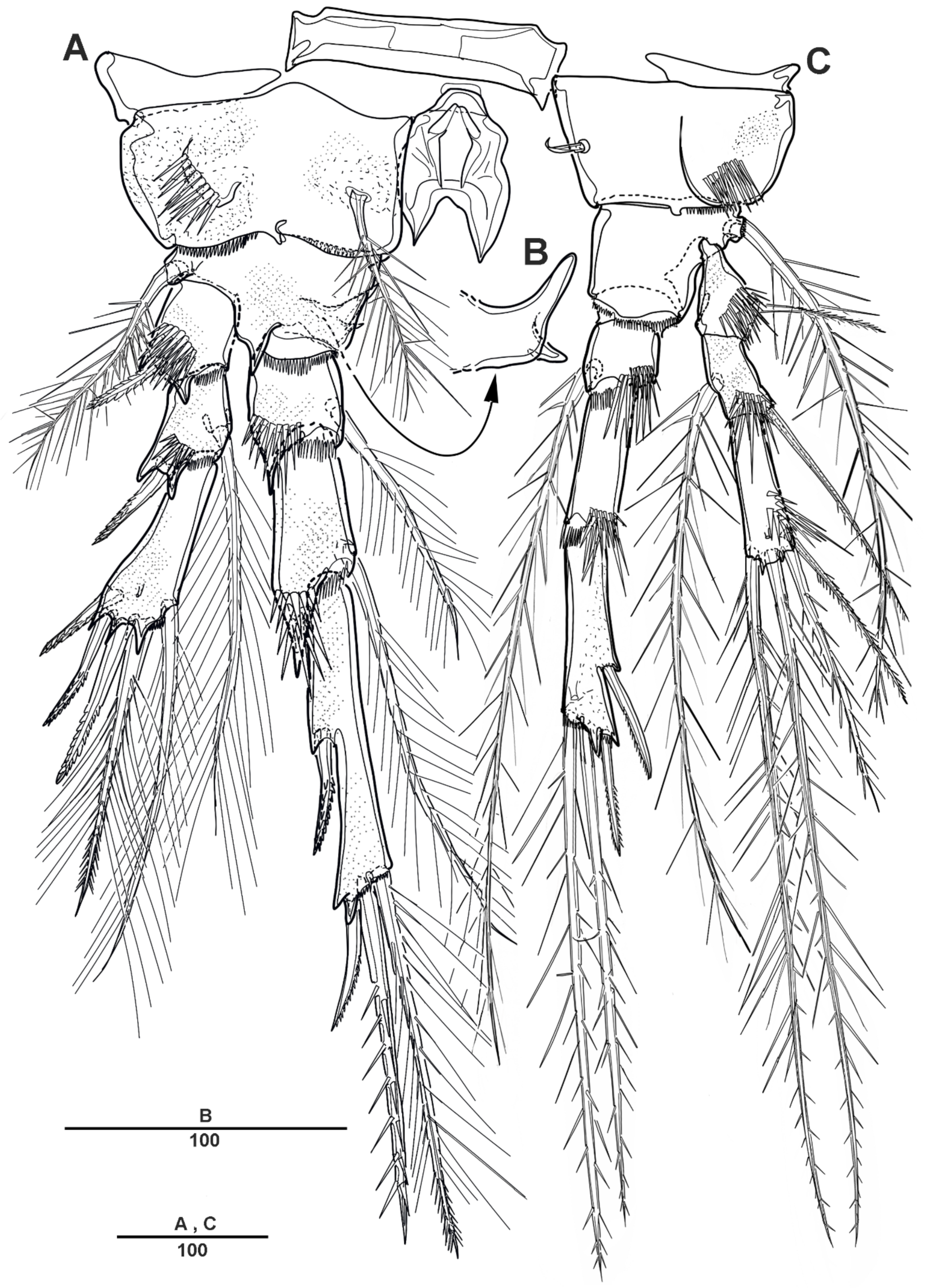
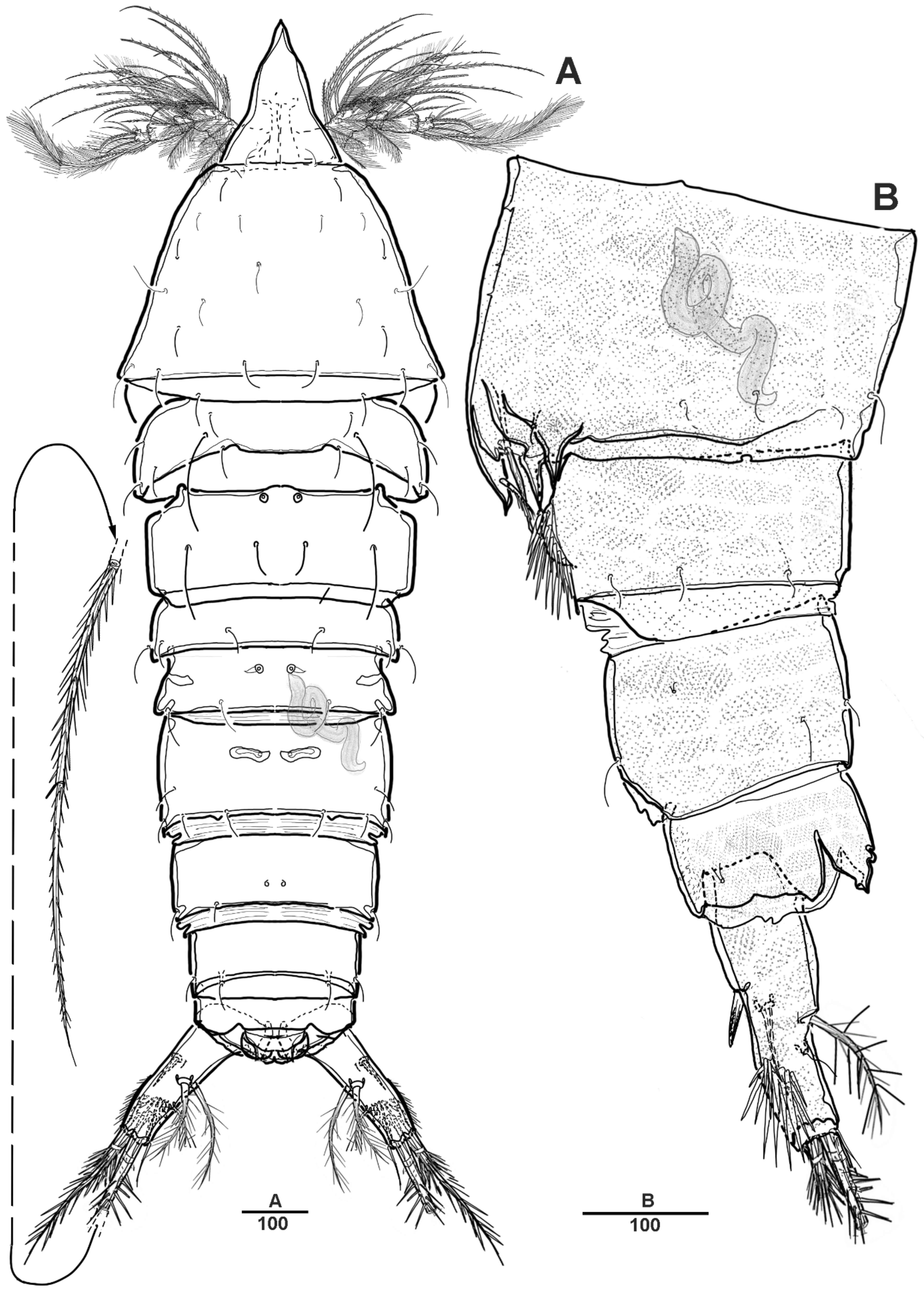
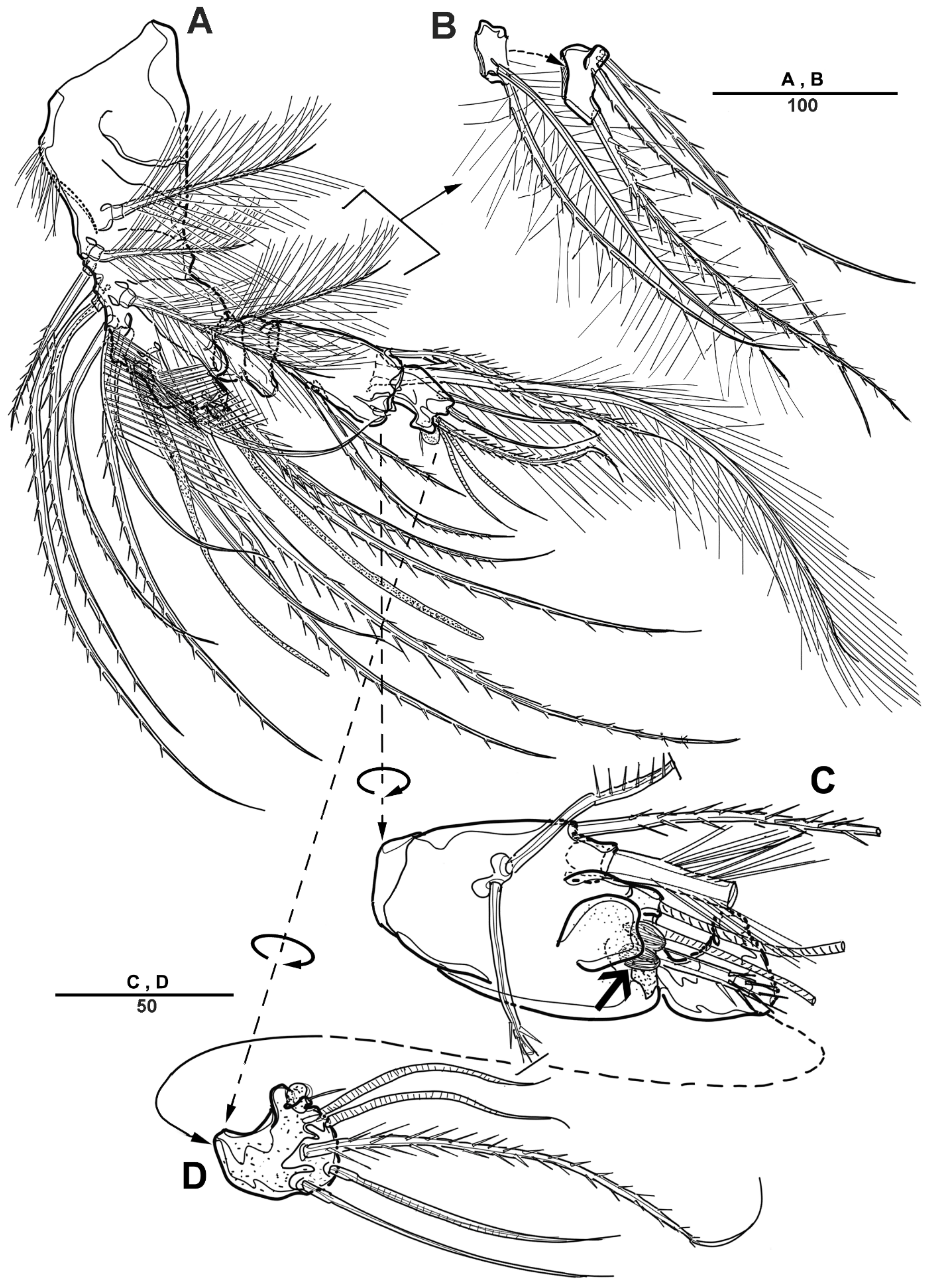
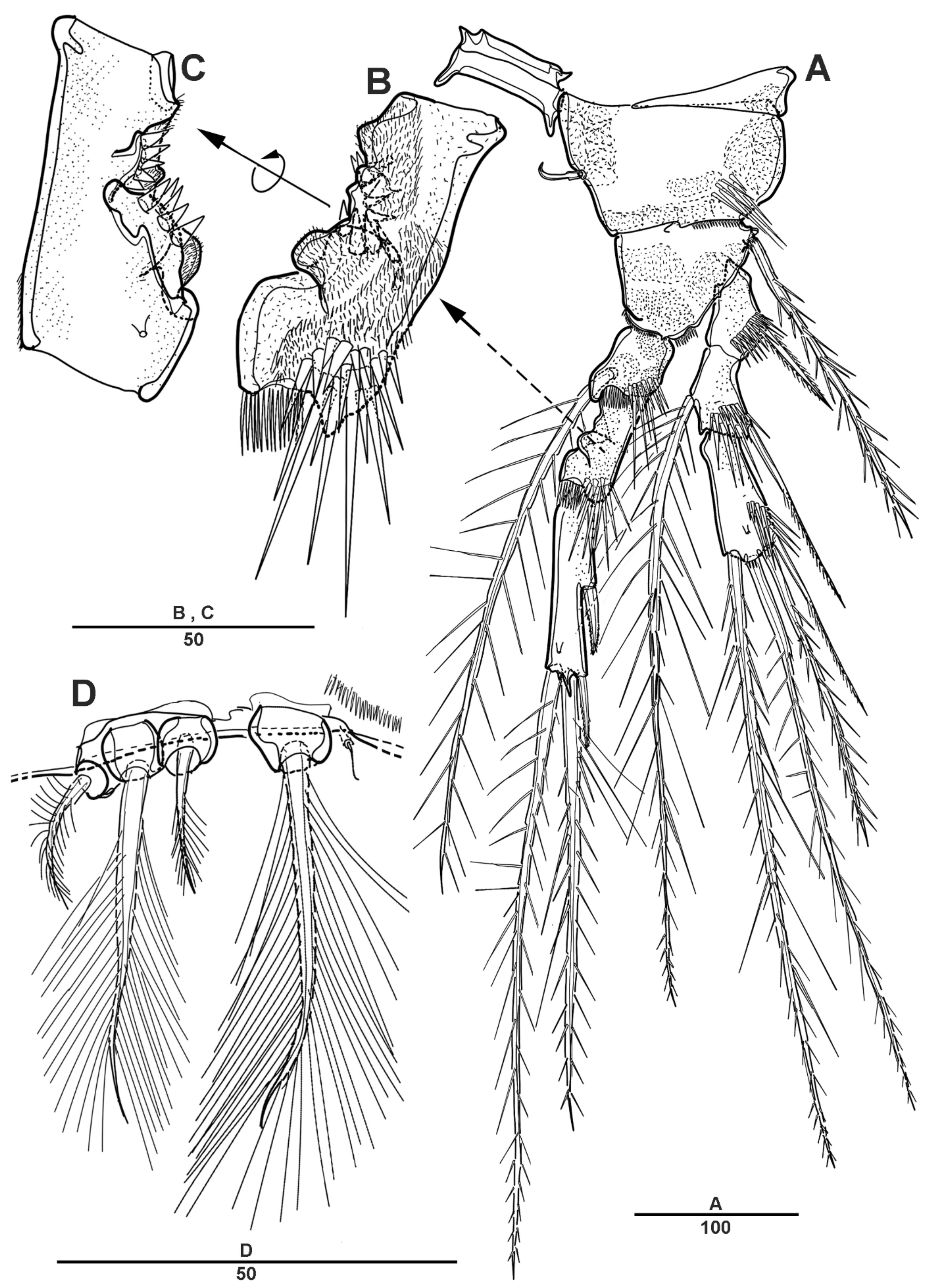
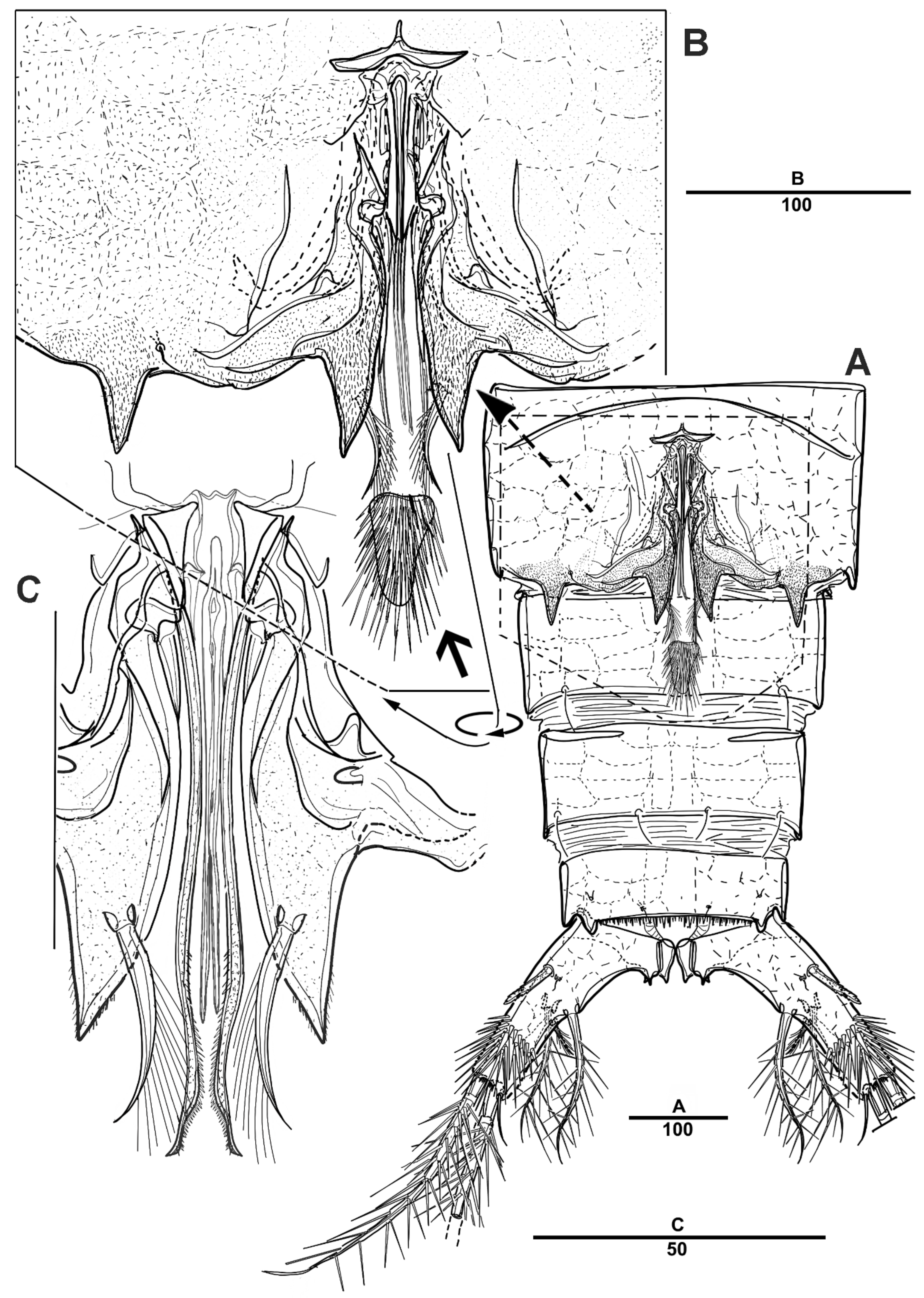
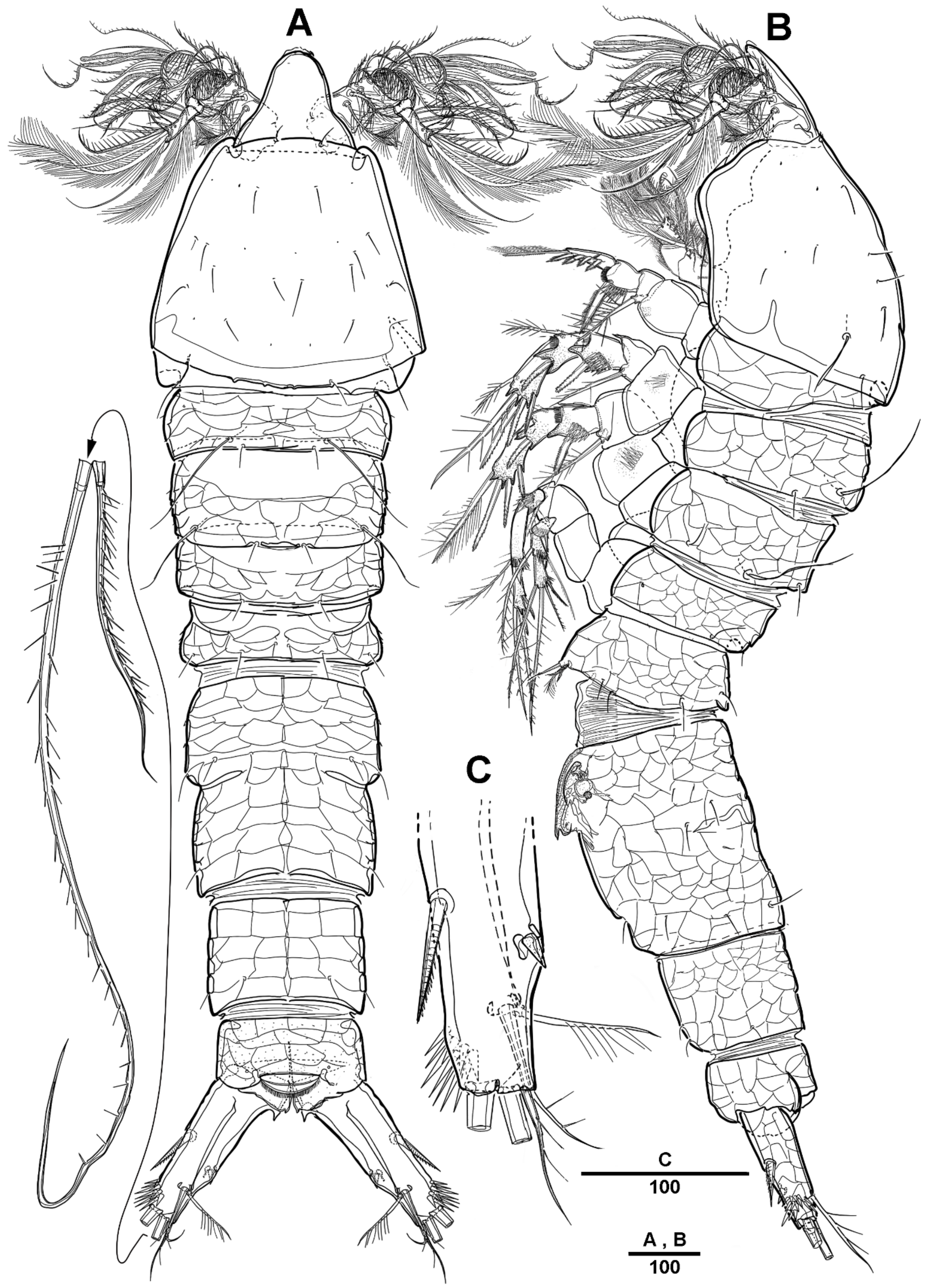
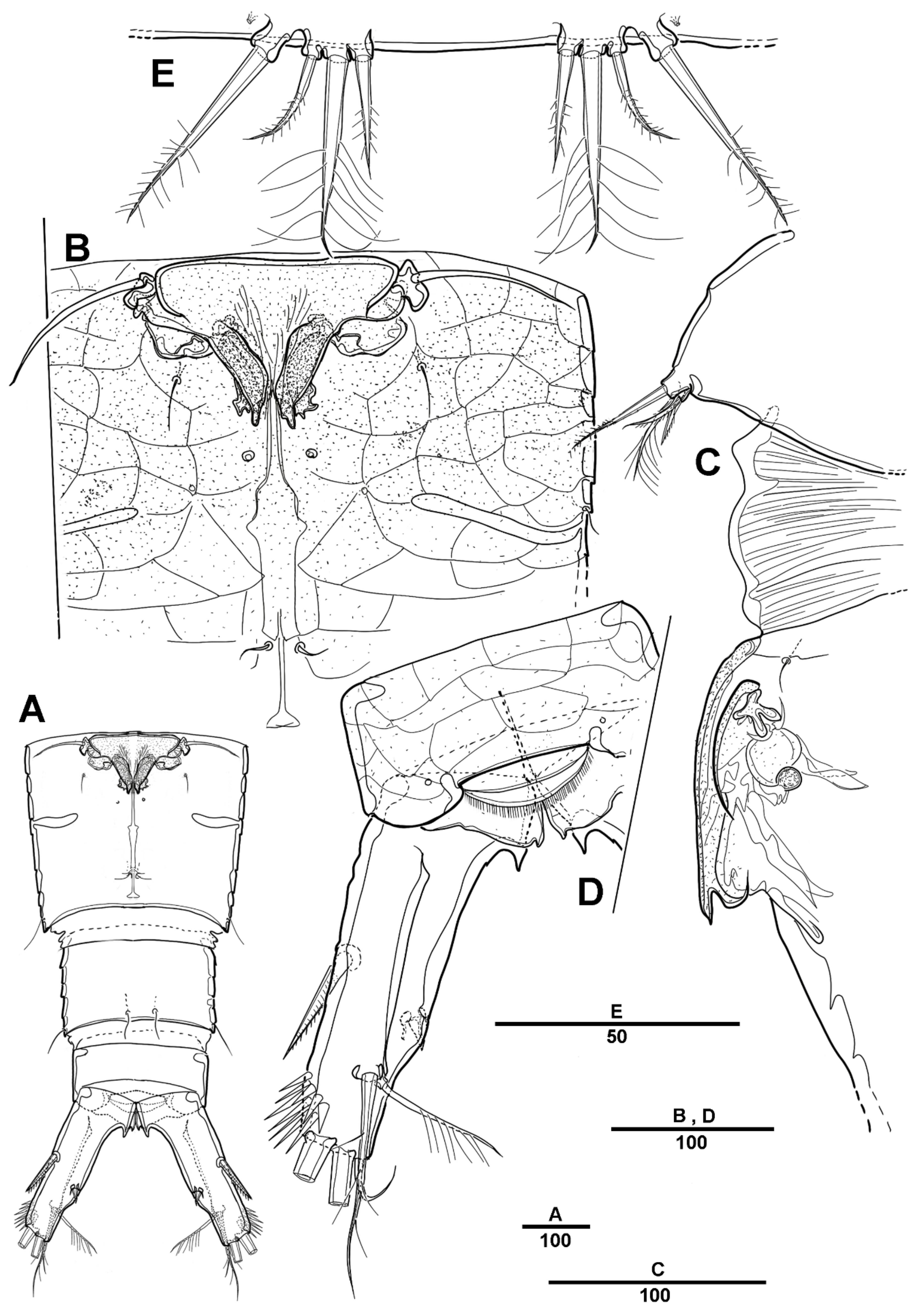
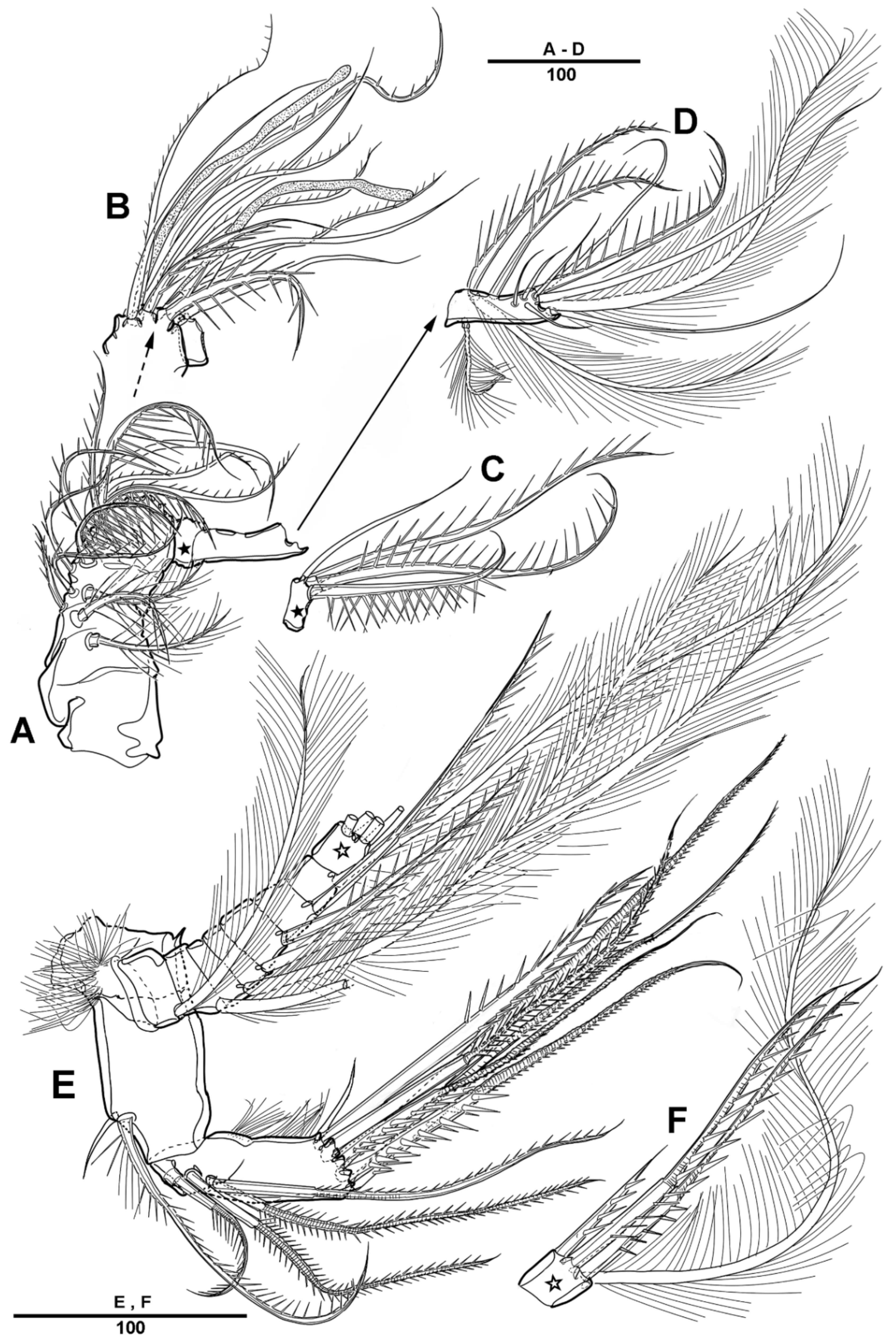
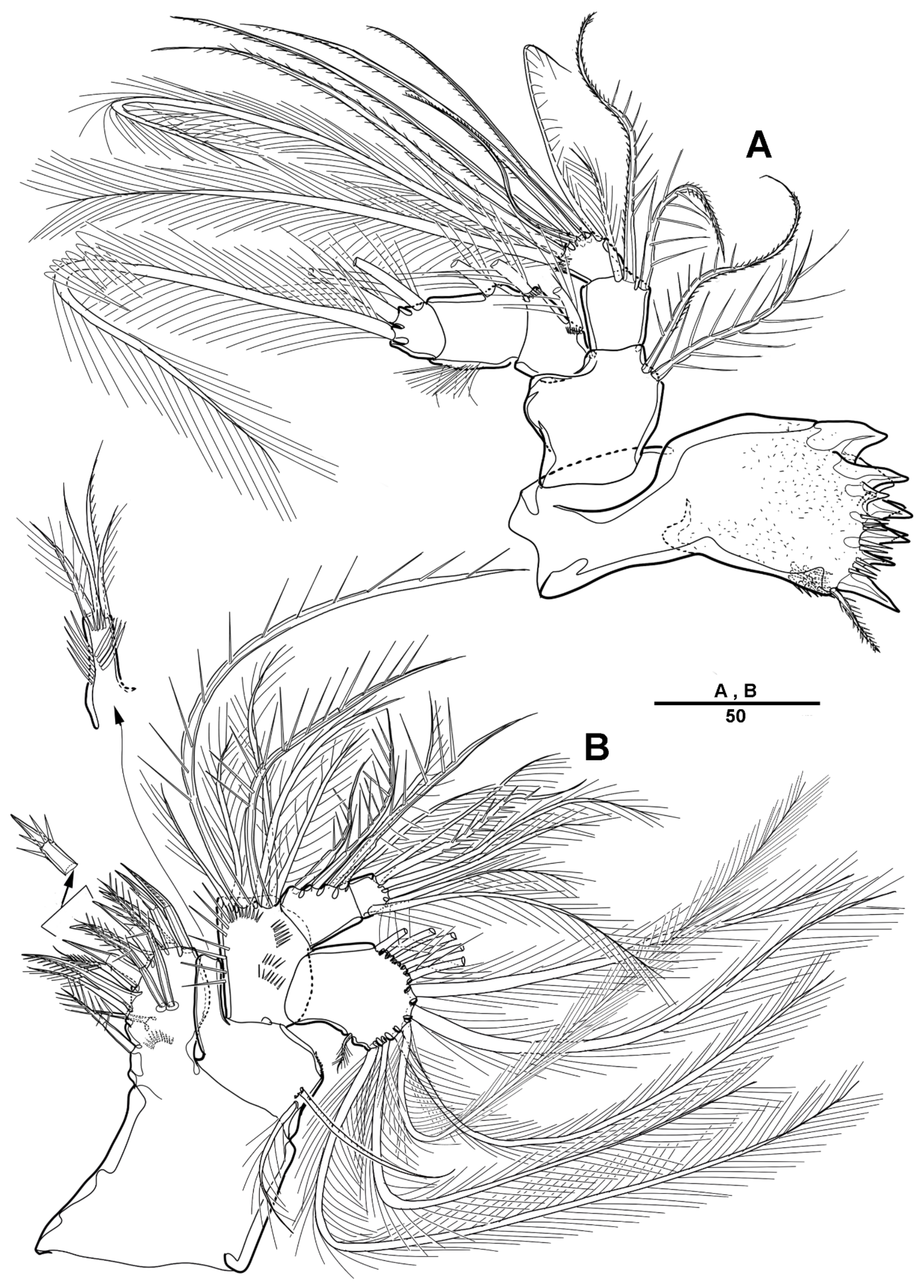
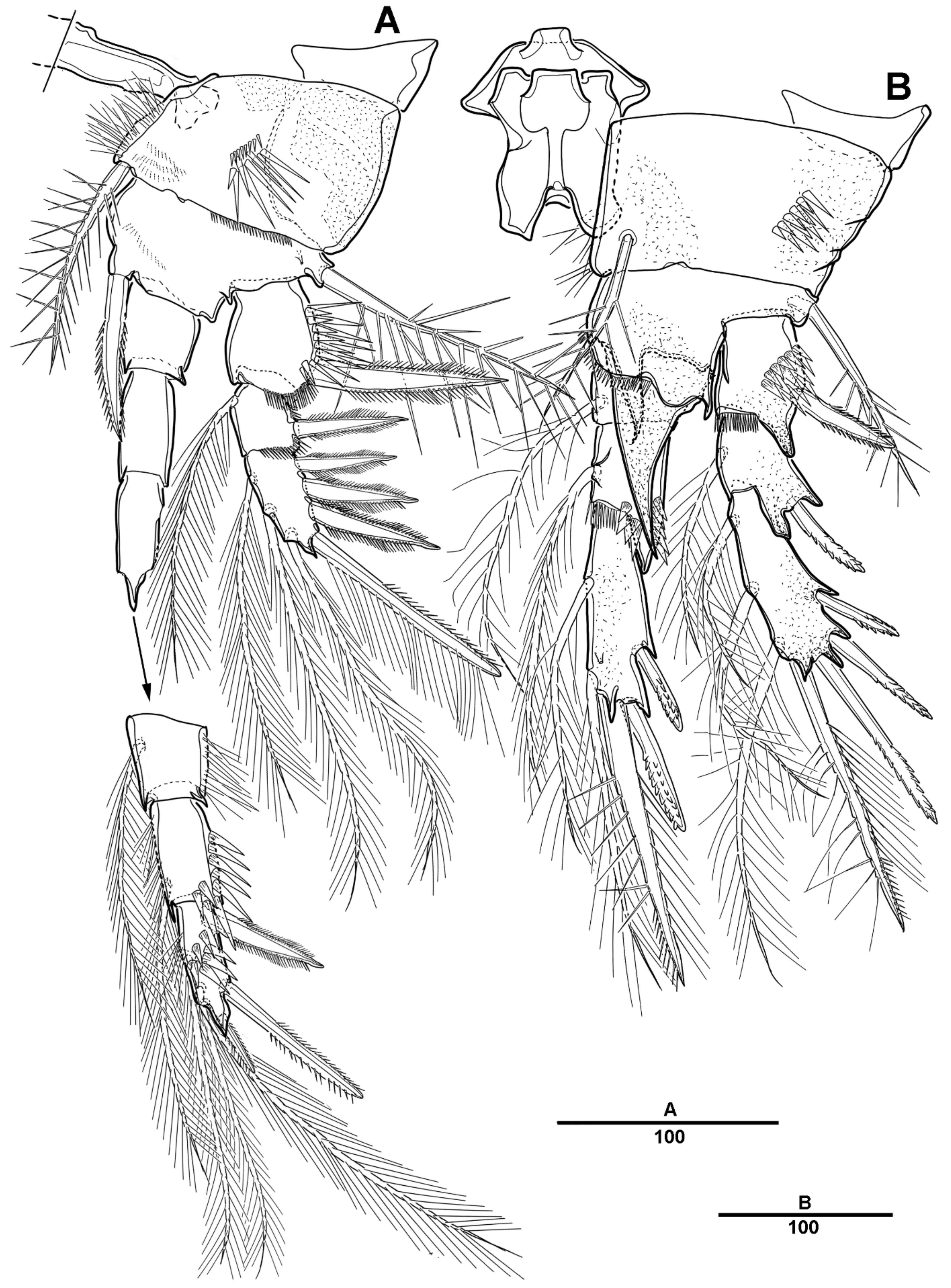
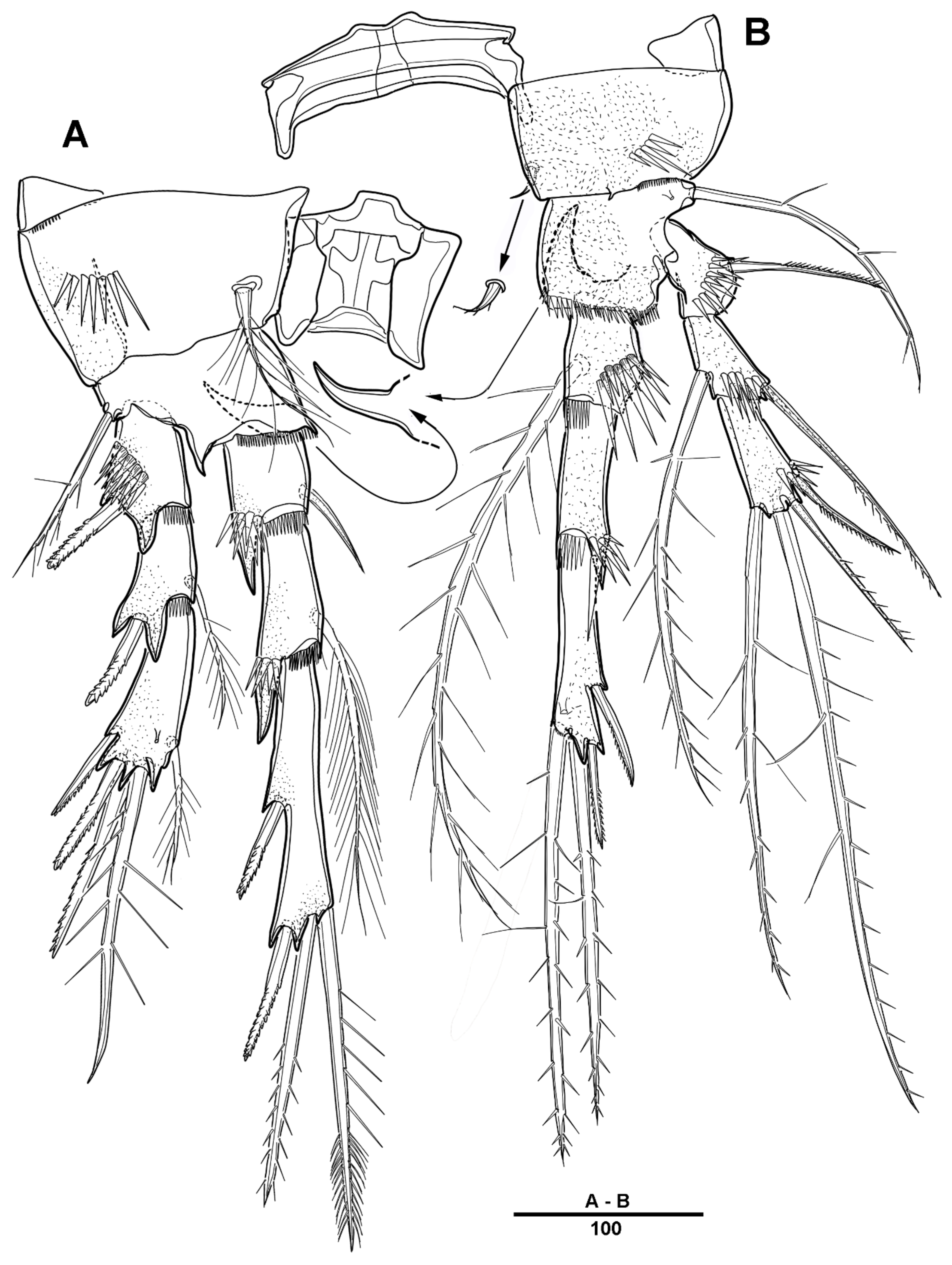
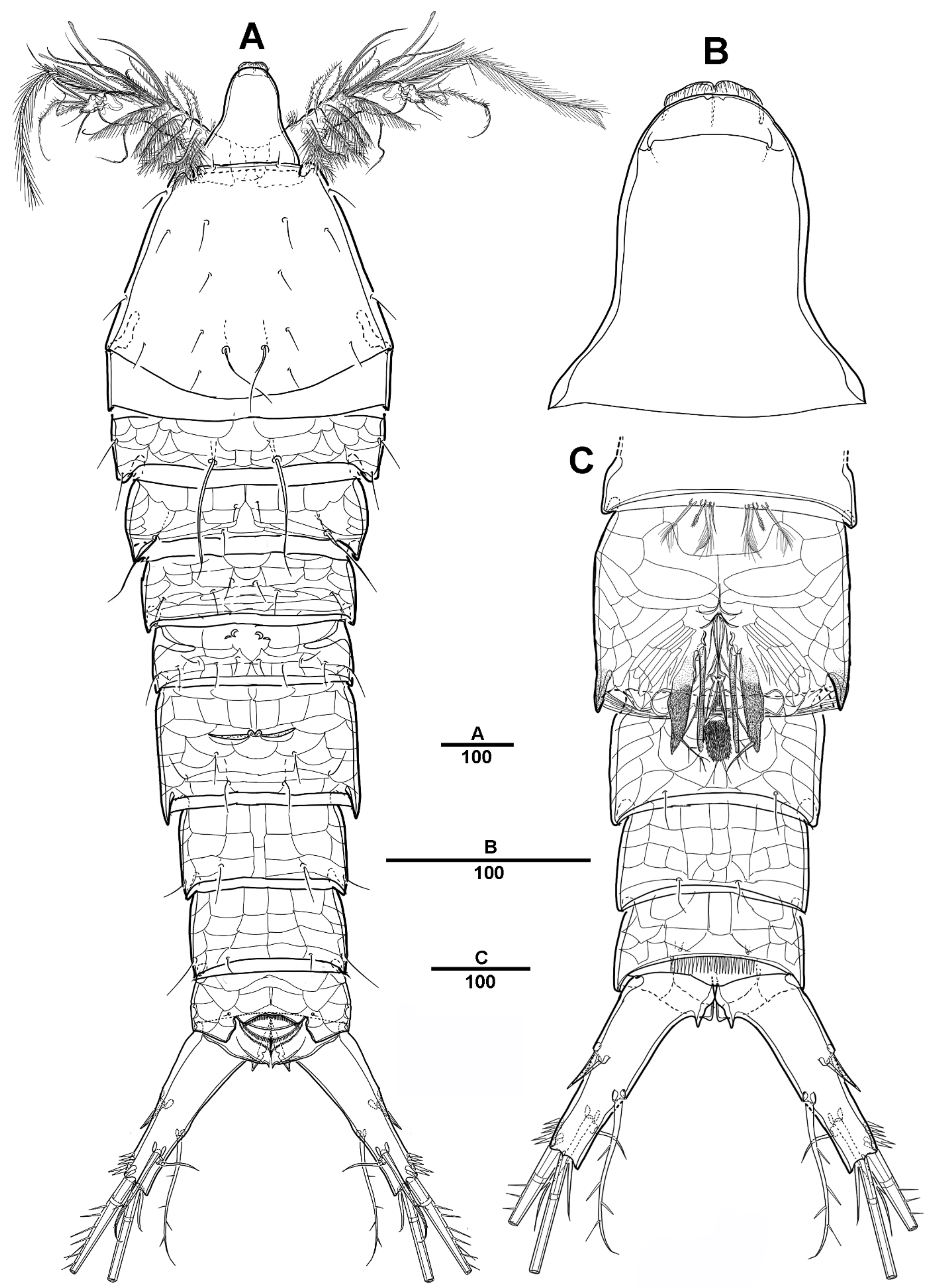
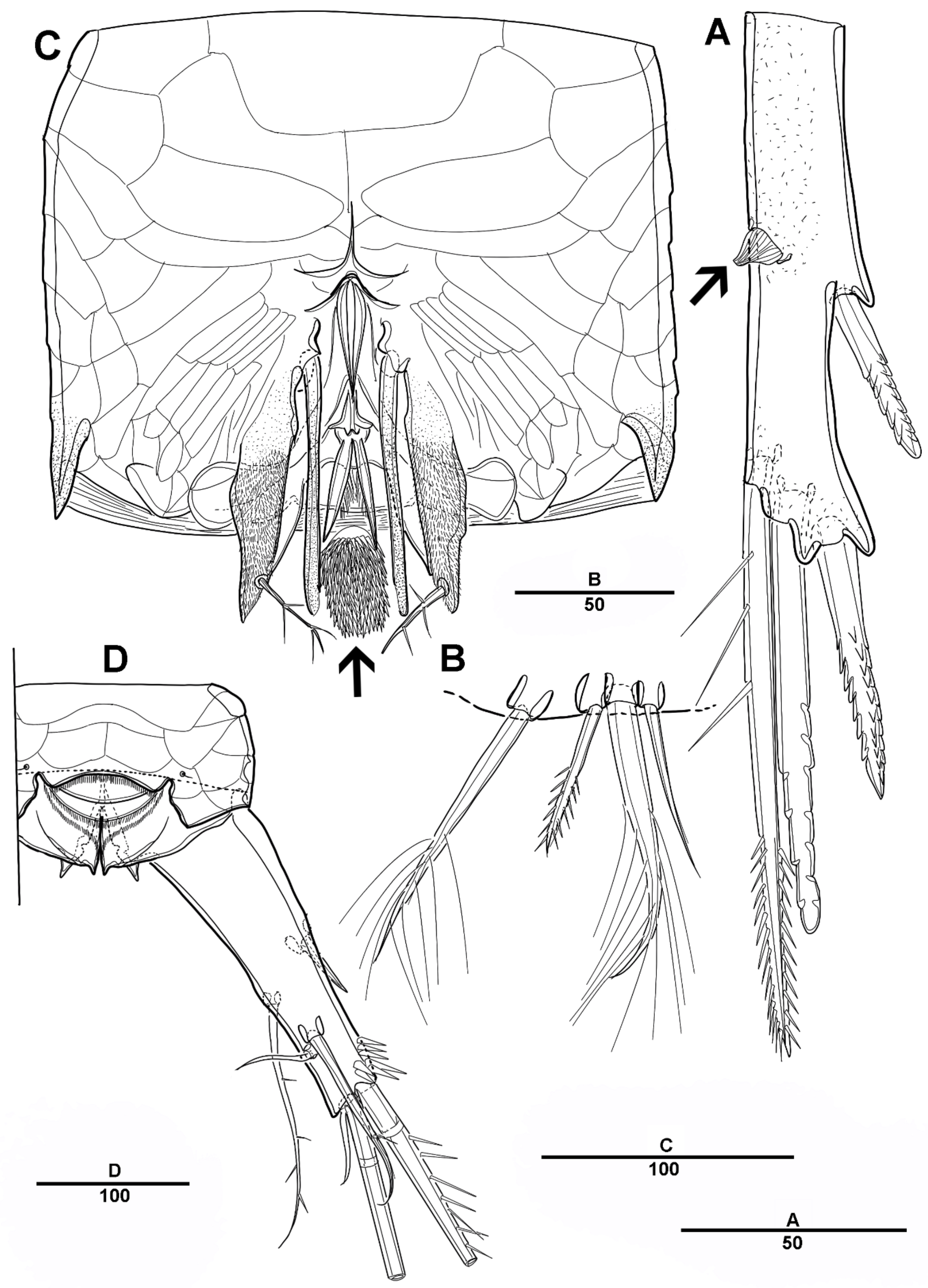
3.1. Scottolana Daecheonensis sp. nov.







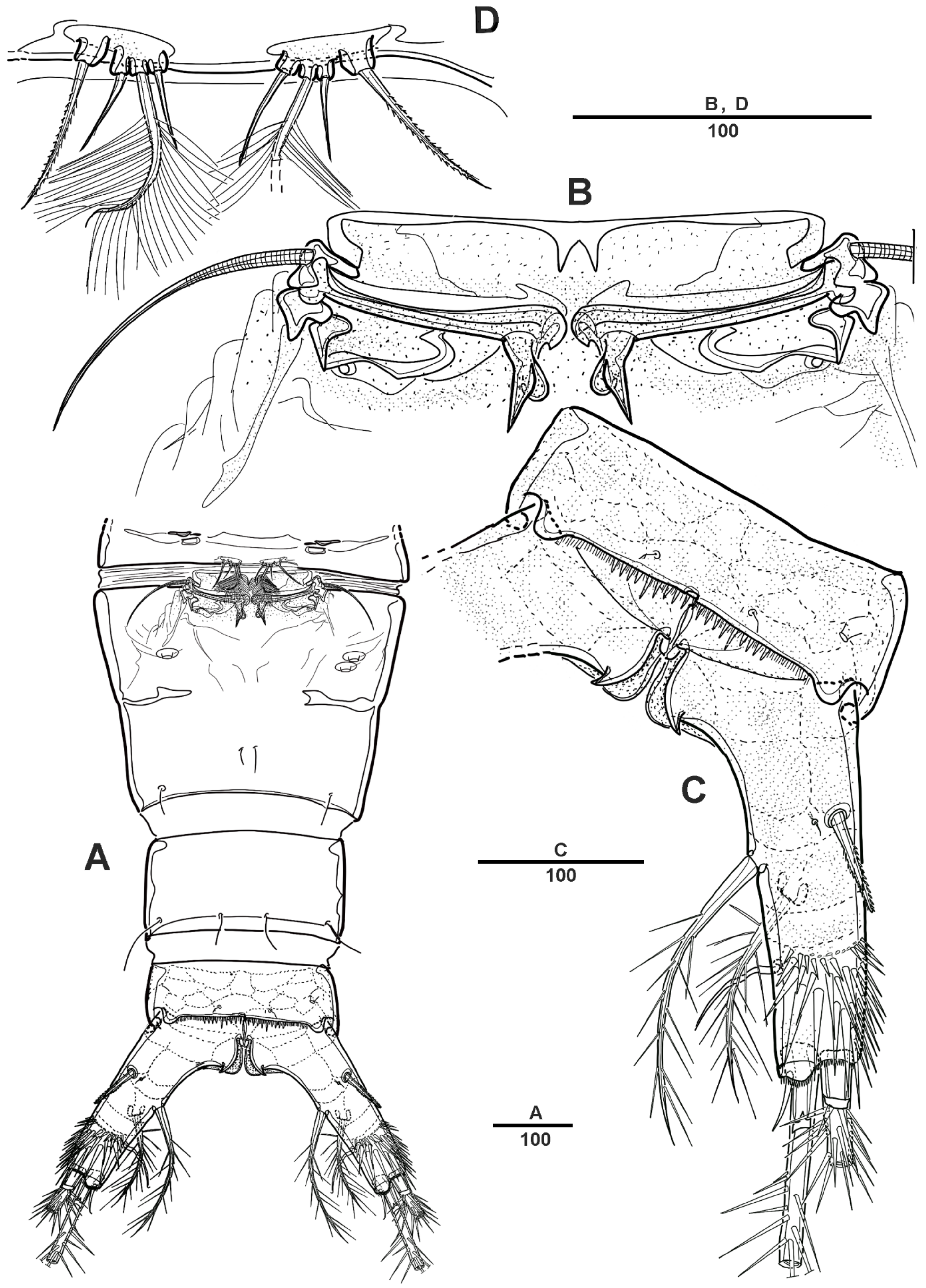
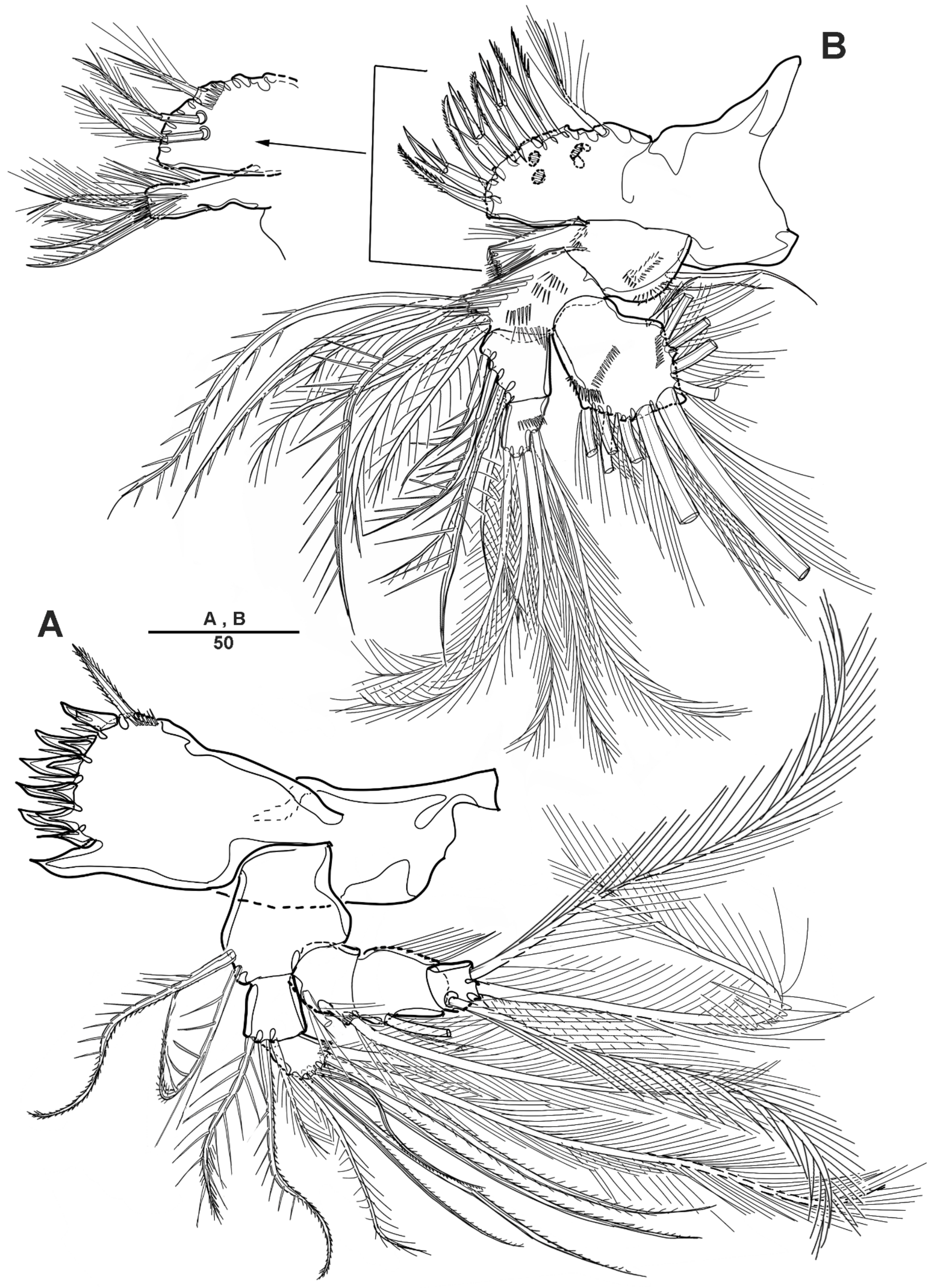
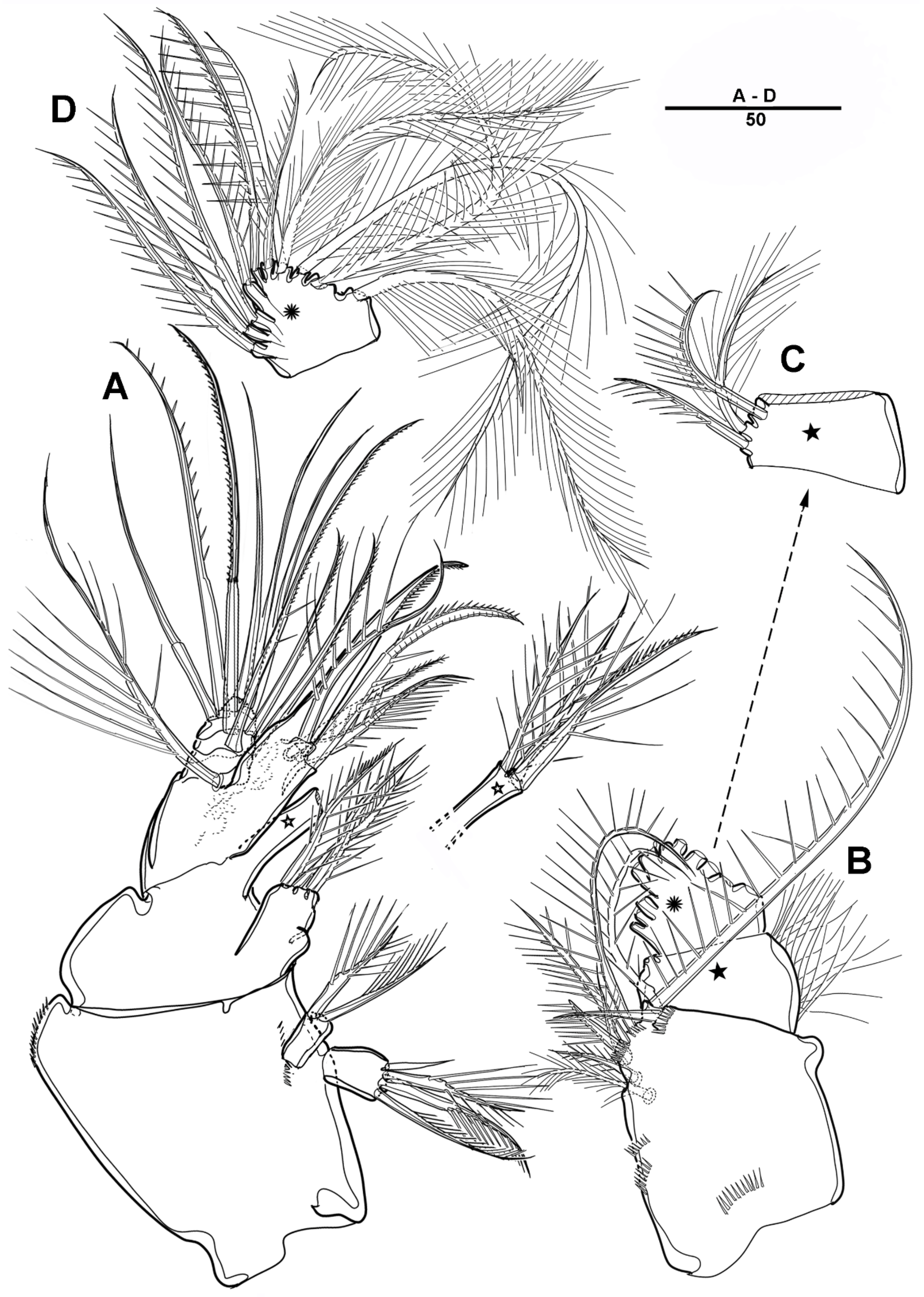
3.2. Scottolana Wonchoeli sp. nov.














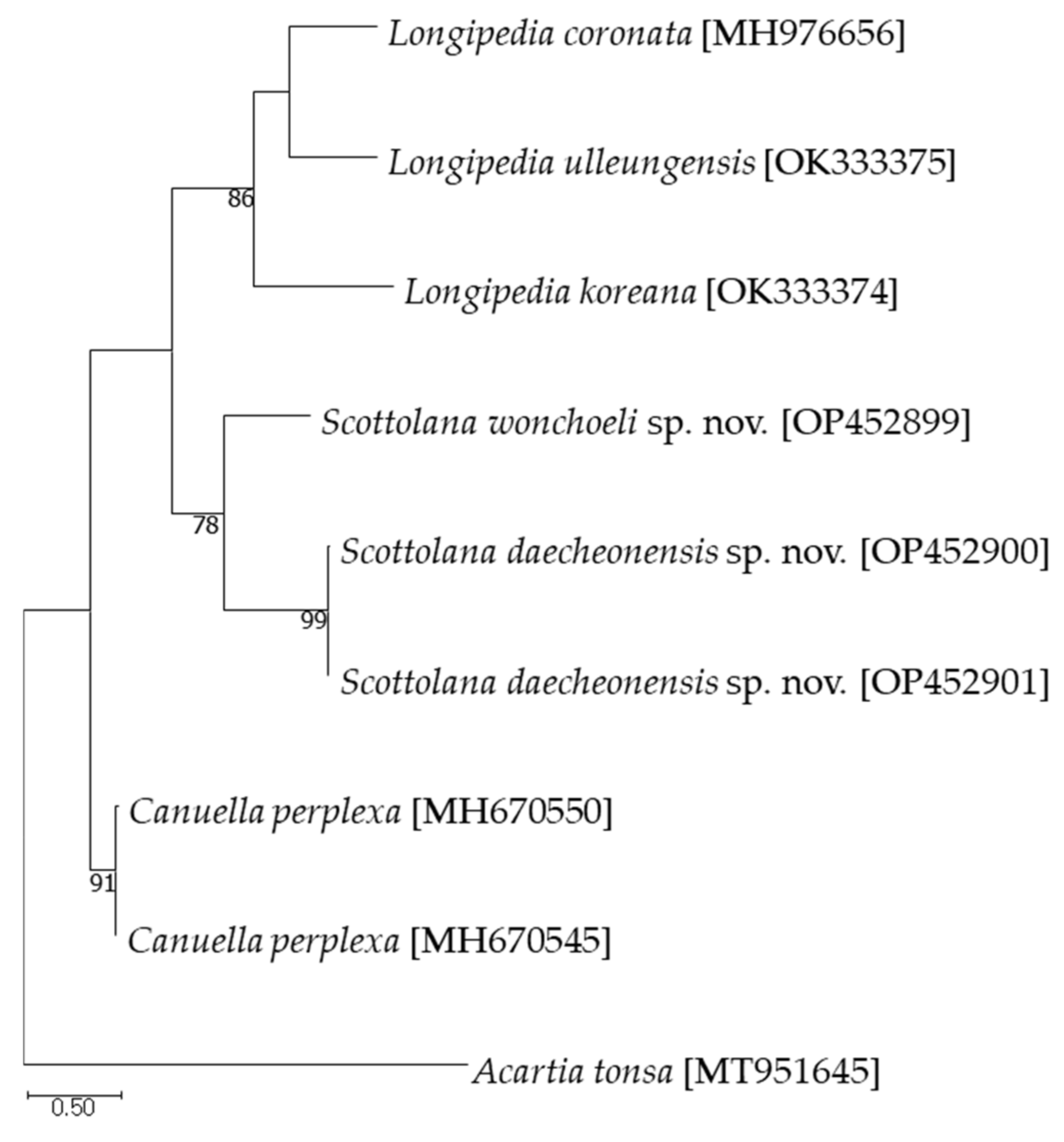
3.3. DNA Sequences in the Family Canuellidae
4. Discussion

| 1. | Urosome with two free abdominal somites in female and three in male; P4 modified in both sexes, including an outward relocation of exopod, and the elongation of several armature elements; P2–P3 basis with a curved spinous process; caudal ramus with a hook-like projection on the inner margin and a spinular patch around the outer distal corner ........................................................................................................ longipes-group ... 2 |
| — | These characters not combined ............................................................................... 9 |
| 2. | Male P3 endopod-3 with modified tube-pore along inner margin ……........... 3 |
| — | Male P3 endopod-3 without modified tube-pore …............................................ 6 |
| 3. | Antennary exopod 9-segmented; female caudal ramus seta II long and plumose with a bulbiform base ...................................................................... S. geei |
| — | Antennary exopod 8-segmented; female caudal ramus seta II with normal base or very short ........................................................................................ 4 |
| 4. | Female antennule 6- or 7-segmented; maxillular endopod with ten setae in total ................................................................................................ S. longipes |
| — | Female antennule 3-segmented; maxillular endopod with eleven setae in total ............................................................................................................... 5 |
| 5. | Rostrum with acutely pointed apex; caudal ramus approximately twice as long as wide .................................................................................... S. jasani |
| — | Rostrum with rounded apex; caudal ramus at least 2.5 times as long as wide ....................................................................................... S. wonchoeli sp. nov. |
| 6. | Female caudal seta II very short ............................................................ S. dissimilis |
| — | Female caudal seta II with bulbiform base ................................................ S. huysi |
| — | Female caudal seta II with normal base ................................................................. 7 |
| 7. | Caudal ramus less than 1.3 times as long as wide; maxillulary endopod with ten setae in total; P4 exopod extends to about the end of endopod-3 .......................................................................................... S. uxoris |
| — | Caudal ramus about twice as long as wide; maxillulary endopod with eleven setae in total; P4 exopod extends to about halfway along endopod-3 .................................................................................................................. 8 |
| 8. | Rostrum without lateral sensilla; female antennule 3-segmented; mandibular exopod 4-segmented; caudal seta V modified to a bulbous process in female .......................................................................... S. gomezi |
| — | Rostrum with two lateral sensilla; female antennule 4-segmented; mandibular exopod 4-segmented; caudal seta V with normal base in female ............................................................................. S. daecheonensis sp. nov. |
| 9. | P2 endopod-3 with six setae; P4 exopod-3 with five setae ................... S. bulbosa |
| — | P2 endopod-3 with five setae; P4 exopod-3 with four setae ............................. 10 |
| 10. | P1 endopod-1 without inner seta ................................................................. S. scotti |
| — | P1 endopod-1 with an inner seta .......................................................................... 11 |
| 11. | P4 exopod extends only to about the end of endopod-2 .................. S. brevifurca |
| — | P4 exopod extends to about halfway along endopod-3 .................................... 12 |
| — | P4 exopod extends to about the end of endopod-3 ............................................ 13 |
| 12. | Caudal seta II long and smooth ............................................................. S. inopinata |
| — | Caudal seta II very thick ....................................................................... S. antillensis |
| 13. | Caudal ramus about as long as wide ......................................................... S. glabra |
| — | Caudal ramus about 1.5 times as long as wide ...................................... S. rostrata |
| — | Caudal ramus about twice as long as wide ......................................................... 14 |
| 14. | Antennary exopod 8-segmented ............................................................ S. bulbifera |
| — | Antennary exopod 7-segmented .......................................................................... 15 |
| 15. | Caudal setae all slender and filiform ......................................................... S. oleosa |
| — | Caudal ramus with two bulbiform setae ........................................... S. tumidiseta |
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lang, K. Monographie der Harpacticiden (Vorläufige Mitteilung); Almqvist & Wiksells Boktryckeri Ab: Uppsala, Sweden, 1944; pp. 1–39. [Google Scholar]
- Dahms, H.U. Usefulness of postembryonic characters for phylogenetic reconstruction in Harpacticoida (Crustacea, Copepoda). Bull. Plankt. Soc. Jpn. Spec. 1991, 1991, 87–104. [Google Scholar]
- Dahms, H.U. Exclusion of the Polyarthra from Harpacticoida and its reallocation as an underived branch of the Copepoda (Arthropoda, Crustacea). Invertebr. Zool. 2004, 1, 29–51. [Google Scholar] [CrossRef]
- Khodami, S.; McArthur, J.V.; Blanco-Bercial, L.; Arbizu, P.M. RETRACTED ARTICLE: Molecular Phylogeny and Revision of Copepod Orders (Crustacea: Copepoda). Sci. Rep. 2017, 7, 9164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhailov, K.V.; Ivanenko, V.N. Low support values and lack of reproducibility of molecular phylogenetic analysis of Copepoda orders. bioRxiv 2019. [Google Scholar] [CrossRef]
- Walter, T.C.; Boxshall, G. World of Copepods database. Canuellidae Lang. 1944. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=115141 (accessed on 23 September 2022).
- Chislenko, L.L. Novye massovye formy garpaktitsid (Copepoda, Harpacticoida) iz zaliva Pos’eta Yaponskogo morya. [New common forms of harpacticids (Copepoda, Harpacticoida) from Possjet Bay of the Sea of Japan.]. Issled. Fauny Morei 1971, 8, 151–181. (In Russian) [Google Scholar]
- Fiers, F. New Canuellidae from the northern coast of Papua New Guinea (Copepoda: Harpacticoida). Bull. Inst. Royal Sci. Nat. Belg. Biologie. 1982, 54, 1–32. [Google Scholar]
- Mu, F.H.; Huys, R. Canuellidae (Copepoda, Harpacticoida) from the Bohai Sea, China. J. Nat. Hist. 2004, 38, 1–36. [Google Scholar] [CrossRef]
- Song, S.J.; Kangtia, P.; Khim, J.S.; Chullasorn, S. Two new Asian species of the genus Scottolana Huys, 2009 (Copepoda: Canuelloida: Canuellidae). J. Nat. Hist. 2018, 52, 377–403. [Google Scholar] [CrossRef]
- Thompson, I.C.; Scott, A. Report on the Copepoda collected by Professor Herdman, at Ceylon, in 1902. Rep. Govt. Ceylon Pearl Oyster Fish. Gulf Manaar 1903, 1 (Suppl. S7), 227–307. [Google Scholar]
- Sewell, R.B.S. Copepoda Harpacticoida. Sci. Rep. John Murray Exped. 1940, 7, 117–382. [Google Scholar]
- Wells, J.B. VII. The Littoral Copepoda (Crustacea) of Inhaca Island, Mozambique. Earth Environ. Sci. Trans. R. Soc. Edinb. 1967, 67, 189–358. [Google Scholar] [CrossRef]
- Wells, J.B.J.; Rao, C.G. Littoral Harpacticoida (Crustacea: Copepoda) from Andaman and Nicobar Islands. Mem. Zool. Surv. India 1987, 16, 1–385. [Google Scholar]
- Fiers, F. Harpacticoid copepods from the West Indian Islands: Canuellidae and Longipediidae (Copepoda, Harpacticoida). Bijdr. Tot Dierkd. 1984, 54, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Por, F.D. A study of the levantine and pontic harpacticoida (Crustacea, Copepoda). Zool. Verh. 1964, 64, 1–128. [Google Scholar]
- Por, F.D. A note on two new species of Canuellidae (Copepoda, Harpacticoida) from the Red Sea. Crustaceana 1983, 44, 187–197. [Google Scholar] [CrossRef]
- Nazari, F.; Mirshamsi, O.; Sari, A.; Aliabadian, M.; Martinez Arbizu, P. Three new Canuellidae (Copepoda: Canuelloida) from Iran. Zootaxa 2018, 4446, 401–441. [Google Scholar] [CrossRef]
- Park, E.O.; Lee, W. New record of Scottolana bulbifera (Copepoda: Harpacticoida: Canuellidae) from Korea. Bull. NIBR 2011, 2, 66–75. [Google Scholar]
- Burgess, R. An improved protocol for separating meiofauna from sediments using colloidal silica sols. Mar. Ecol. Prog. Ser. 2001, 214, 161–165. [Google Scholar] [CrossRef]
- Lim, B.J.; Bang, H.W.; Moon, H.; Back, J. Integrative description of Diosaccus koreanus sp. nov. (Hexanauplia, Harpacticoida, Miraciidae) and integrative information on further Korean species. ZooKeys 2020, 927, 1–35. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape split by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991; pp. 1–468. [Google Scholar]
- Por, F.D. Level bottom Harpacticoida (Crustacea, Copepoda) from Elat (Red Sea), part I. Isr. J. Zool. 1967, 16, 101–165. [Google Scholar]
- Huys, R. Unresolved cases of type fixation, synonymy and homonymy in harpacticoid copepod nomenclature (Crustacea: Copepoda). Zootaxa 2009, 2183, 1–99. [Google Scholar] [CrossRef]
- Huys, R. Harpacticoid copepods—Their symbiotic associations and biogenic substrata: A review. Zootaxa 2016, 4174, 448–729. [Google Scholar] [CrossRef]
- Wells, J.B. An annotated checklist and keys to the species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.Y.; Jang, K.H.; Choi, E.H.; Ryu, S.H.; Kim, S.K.; Lee, J.H.; Lim, Y.J.; Lee, J.; Jun, J.; Kwak, M.; et al. DNA Barcoding of Metazoan Zooplankton Copepods from South Korea. PLoS ONE 2016, 11, e0157307. [Google Scholar] [CrossRef]
- Yeom, J.; Park, N.; Jeong, R.; Lee, W. Integrative description of cryptic Tigriopus species from Korea using MALDI-TOF MS and DNA barcoding. Front. Mar. Sci. 2021, 8, 648197. [Google Scholar] [CrossRef]
- Bang, H.W.; Moon, H.; Back, J. Two New Species of the Genus Longipedia Claus, 1863 (Copepoda: Harpacticoida: Longipediidae) from Korea, with an Update and a Key to Species. Diversity 2021, 13, 590. [Google Scholar] [CrossRef]
- Wu, S.; Xiong, J.; Yu, Y. Taxonomic resolutions based on 18S rRNA genes: A case study of subclass Copepoda. PLoS ONE 2015, 10, e0131498. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Date | Locality | Methods/ Sediment/Depth | Specimen Nos. |
|---|---|---|---|---|
| Genus Scottolana Huys, 2009 | ||||
| S. daecheonensis sp. nov. | 13 May 2021 | 36°13′18″ N 126° 31′32″ E | Light trap/ Muddy/2 m | MABIK CR00252814, MABIK CR00252811 |
| 05 June 2019 | 36°18′00″ N 126°31′02″ E | Hand net/ Rock pool/0 m | MABIK CR00252815, MABIK CR00252812 | |
| 27 March 2018 | 34°54′28″ N 128°9′ 31″ E | Hand net/ Sand beach/0 m | MABIK CR00252816, MABIK CR00252813 | |
| S. wonchoeli sp. nov. | 26 July 2017 | 37°29′13″ N 130°55′20″ E | SCUBA/ Sandy/5 m | MABIK CR00252817, MABIK CR00252818 |
| Gene | Primer | Primer Sequence | PCR Condition |
|---|---|---|---|
| COI | MaxilloF MaxilloR | CWAAYCATAAAGAYATTG ACTTCAGGRTGNCCAAAR | 94 °C, 300 s; 40 cycles × (94 °C, 60 s; 46 °C, 120 s; 72 °C, 180 s; 72 °C, 600 s) |
| 18S rRNA | 18S F1 18S R9 18S F2 18S F3 18S F4 18S R6 18S R7 18S R8 | TACCTGGTTGATCCTGCCAG GATCCTTCCGCAGGTTCACCTAC CCTGAGAAACGGCTRCCACAT GYGRTCAGATACCRCCSTAGTT GGTCTGTGATGCCCTYAGATGT TYTCTCRKGCTBCCTCTCC GYYARAACTAGGGCGGTATCTG ACATCTRAGGGCATCACAGACC | 94 °C, 300 s; 40 cycle × (94 °C, 30 s; 50 °C, 30 s; 72 °C, 60 s); 72 °C, 420 s Internal primers, 18S F2–F4 and 18S R6–8, were used for primer walking to sequence over 1700 bps. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bang, H.W.; Moon, H.; Back, J. Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009. Diversity 2022, 14, 967. https://doi.org/10.3390/d14110967
Bang HW, Moon H, Back J. Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009. Diversity. 2022; 14(11):967. https://doi.org/10.3390/d14110967
Chicago/Turabian StyleBang, Hyun Woo, Heejin Moon, and Jinwook Back. 2022. "Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009" Diversity 14, no. 11: 967. https://doi.org/10.3390/d14110967
APA StyleBang, H. W., Moon, H., & Back, J. (2022). Two New Species of the Family Canuellidae Lang, 1944 (Copepoda: Polyarthra), from Korea, with a Key to Species of the Genus Scottolana Huys, 2009. Diversity, 14(11), 967. https://doi.org/10.3390/d14110967