
Investigating the Assemblages of Two Groups of Collembola (Strong Furca and Weak Furca) under Different Agricultural Management Systems, Northeastern China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
- MP-MC: Mould plowing among monoculture (corn)
- MP-CS: Mould plowing among corn–soybean rotation
- RT-MC: Ridge tillage among monoculture (corn)
- RT-CS: Ridge tillage among corn–soybean rotation
- NT-MC: No-tillage among monoculture (corn)
- NT-CS: No-tillage among corn–soybean rotation
2.3. Sampling Design
2.4. Statistical Analysis
3. Results
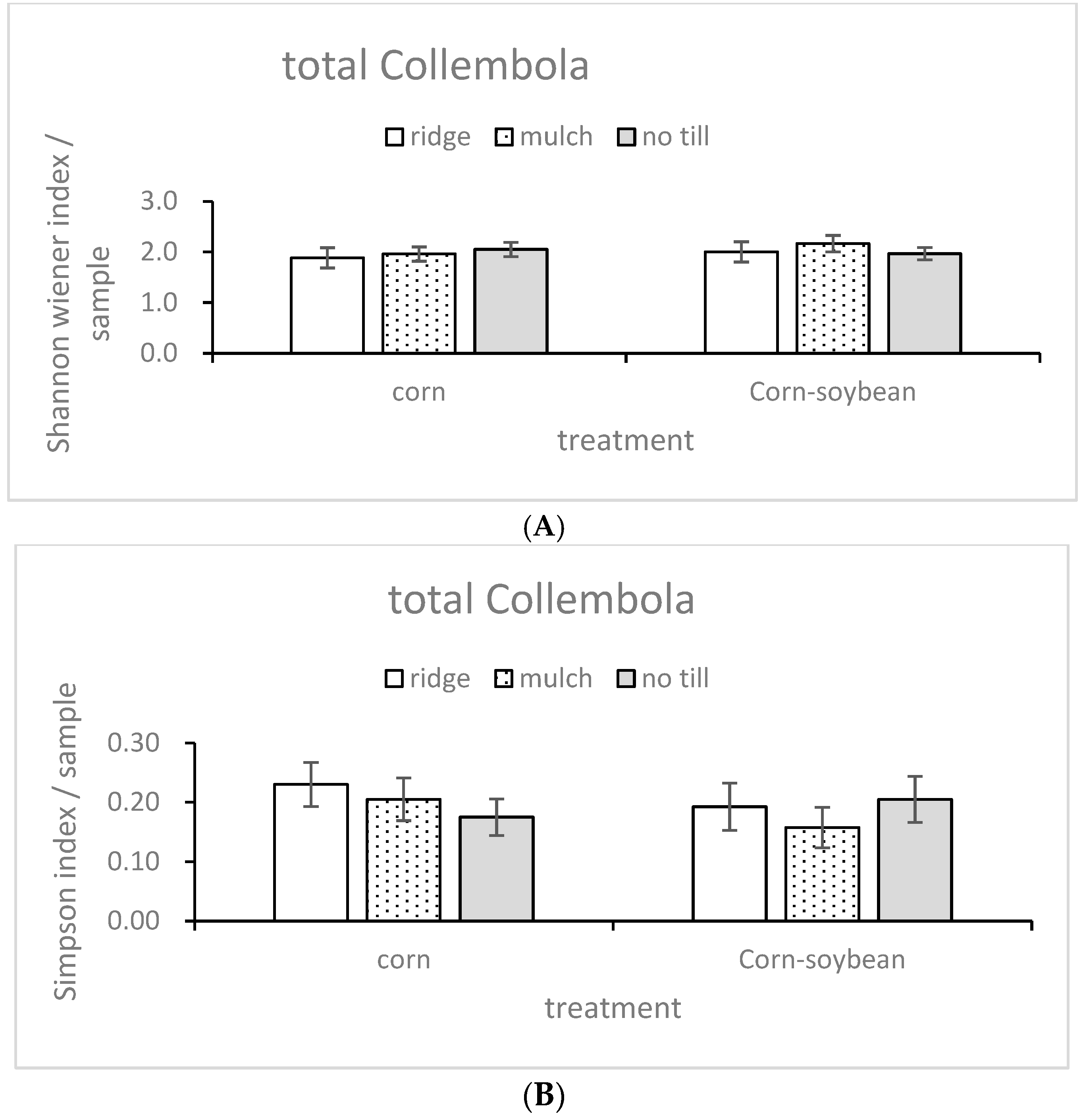
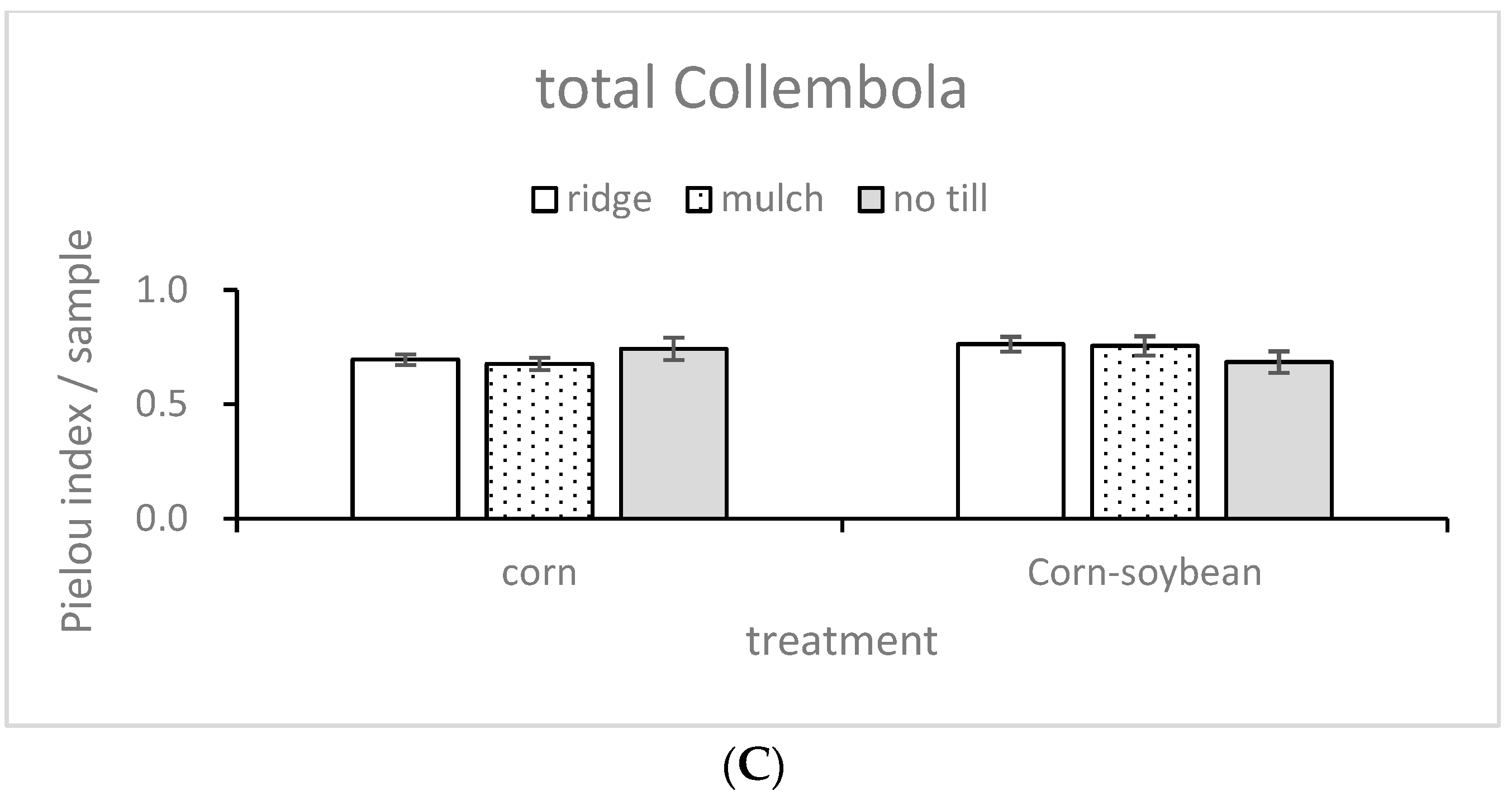
3.1. Community Structure Indices
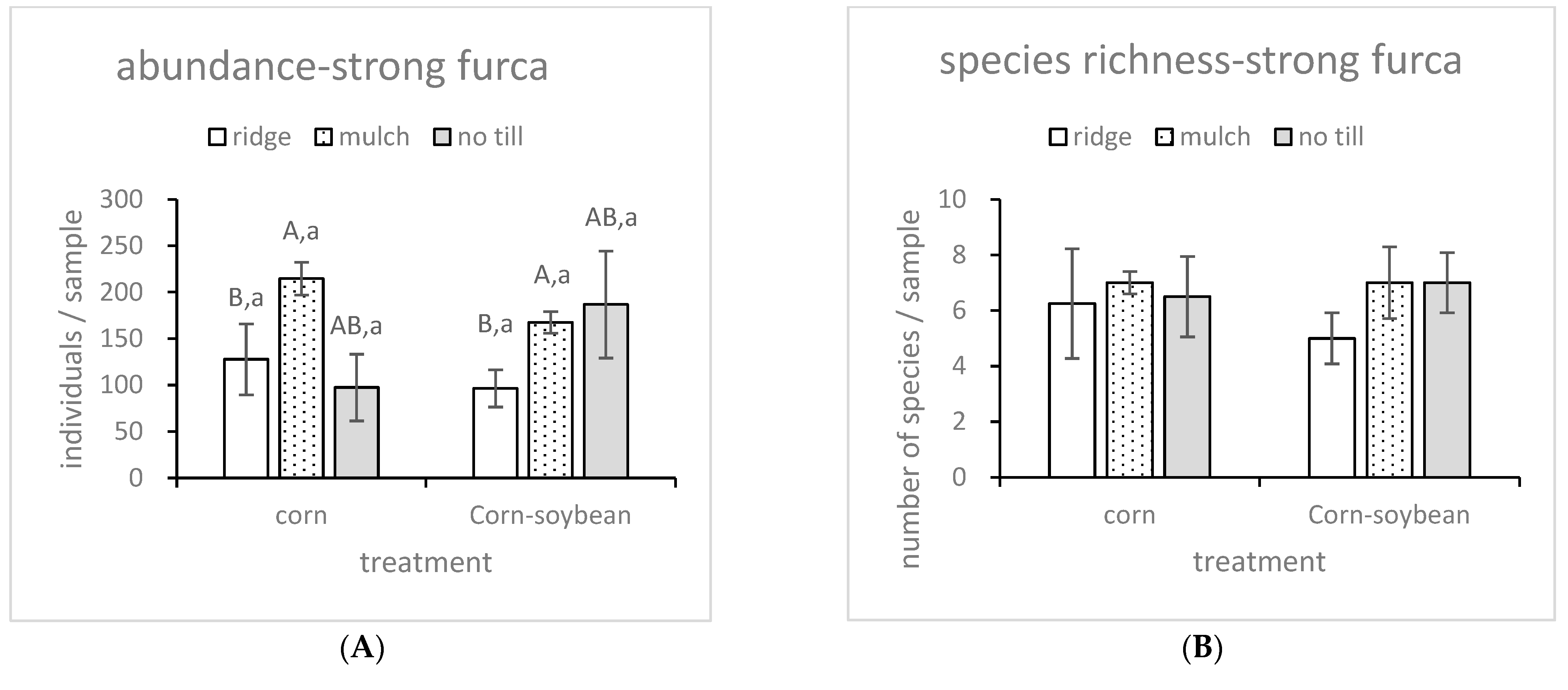
3.2. Different Responses of Species Richness and Abundances of Two Collembola Groups (Strong Furca and Weak Furca) to Different Tillage Types and Rotation Types
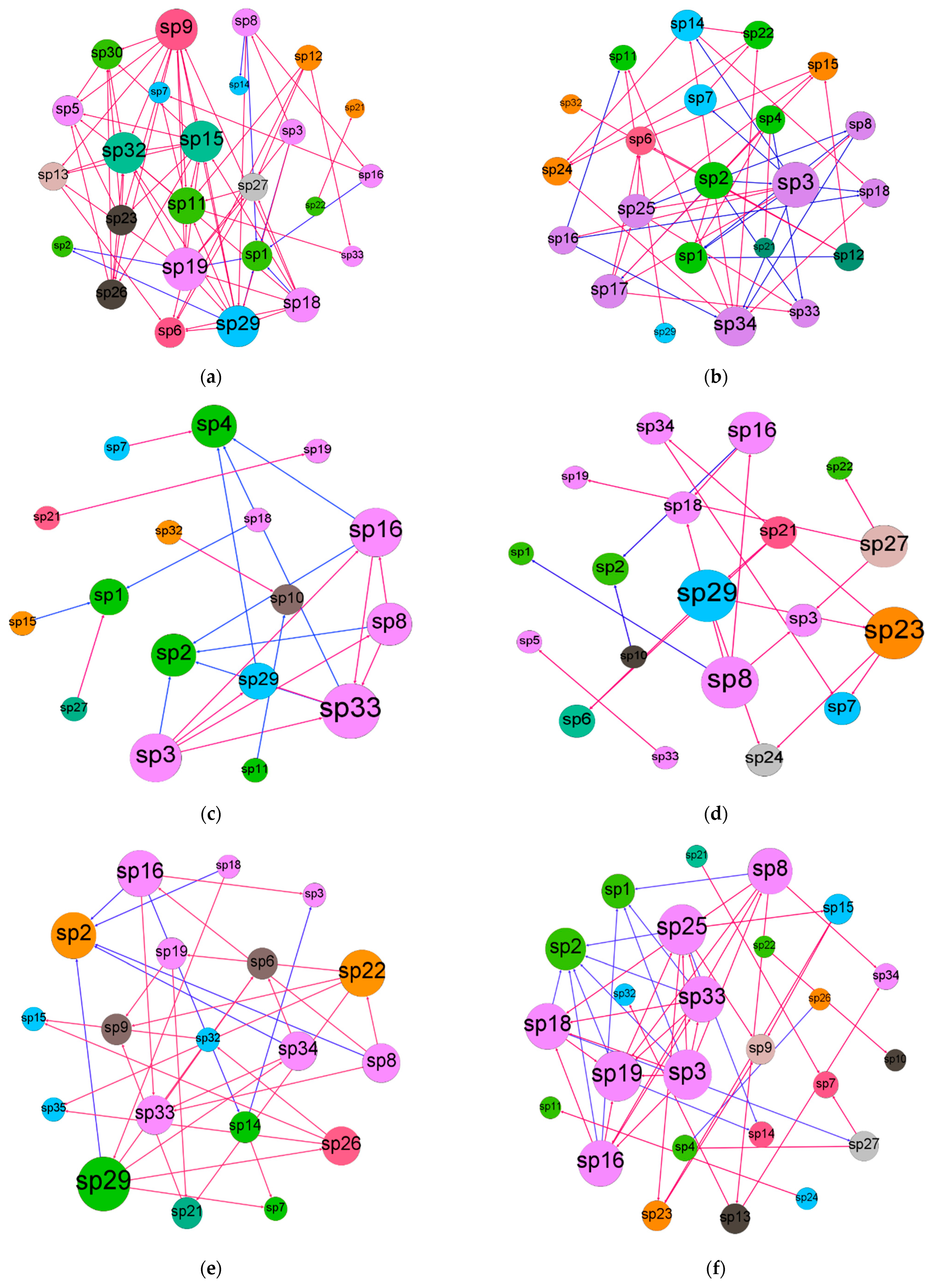
3.3. Co-Occurrence Pattern of Collembola Communities under Six Different Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fukami, T.; Wardle, D.A. Long-term ecological dynamics: Reciprocal insights from natural and anthropogenic gradients. Proc. R. Soc. B Biol. Sci. 2005, 272, 2105–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laliberté, E.; Kardol, P.; Didham, R.K.; Teste, F.P.; Turner, B.L.; Wardle, D.A. Soil fertility shapes belowground food webs across a regional climate gradient. Ecol. Lett. 2017, 20, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, A.; Mineau, P. The impact of agricultural practices on biodiversity. Agric. Ecosyst. Environ. 1995, 55, 201–212. [Google Scholar] [CrossRef]
- González de Molina, M.; Soto Fernández, D.; Guzmán Casado, G.; Infante-Amate, J.; Aguilera Fernández, E.; Vila Traver, J.; García Ruiz, R. The Social Metabolism of Spanish Agriculture, 1900–2008: The Mediterranean Way Towards Industrialization; Springer Nature: Berlin, Germany, 2020. [Google Scholar]
- Postma-Blaauw, M.B.; de Goede, R.G.M.; Bloem, J.; Faber, J.H.; Brussaard, L. Soil biota community structure and abundance under agricultural intensification and extensification. Ecology 2010, 91, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, S.; Coudrain, V.; Hedde, M.; Brunet, N.; Mary, B.; Recous, S.; Chauvat, M. Effect of different crop management practices on soil Collembola assemblages: A 4-year follow-up. Appl. Soil Ecol. 2017, 119, 354–366. [Google Scholar] [CrossRef]
- Ghosh, A.; Misra, S.; Bhattacharyya, R.; Sarkar, A.; Singh, A.K.; Tyagi, V.C.; Kumar, R.V.; Meena, V.S. Agriculture, dairy and fishery farming practices and greenhouse gas emission footprint: A strategic appraisal for mitigation. Environ. Sci. Pollut. Res. 2020, 27, 10160–10184. [Google Scholar] [CrossRef]
- Cluzeau, D.; Guernion, M.; Chaussod, R.; Martin-Laurent, F.; Villenave, C.; Cortet, J.; Ruiz-Camacho, N.; Pernin, C.; Mateille, T.; Philippot, L. Integration of biodiversity in soil quality monitoring: Baselines for microbial and soil fauna parameters for different land-use types. Eur. J. Soil Biol. 2012, 49, 63–72. [Google Scholar] [CrossRef]
- Mahfouz, M.; Mohamed, M. Towards optimization of entomopathogenic nematodes for more service in the biological control. J. Nematol. 2019, 51, 77. [Google Scholar]
- Garnett, T.; Appleby, M.C.; Balmford, A.; Bateman, I.J.; Benton, T.G.; Bloomer, P.; Burlingame, B.; Dawkins, M.; Dolan, L.; Fraser, D. Sustainable intensification in agriculture: Premises and policies. Science 2013, 341, 33–34. [Google Scholar] [CrossRef]
- Ball, B.; Bingham, I.; Rees, R.; Watson, C.; Litterick, A. The role of crop rotations in determining soil structure and crop growth conditions. Can. J. Soil Sci. 2005, 85, 557–577. [Google Scholar] [CrossRef] [Green Version]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, P.R. Conservation agriculture: What is it and why is it important for future sustainable food production? J. Agric. Sci. Camb. 2007, 145, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. In Invertebrate Biodiversity as Bioindicators of Sustainable Landscapes; Elsevier: Amsterdam, The Netherlands, 1999; pp. 19–31. [Google Scholar]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.-F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Michels, K.; Sivakumar, M.; Allison, B. Wind erosion control using crop residue I. Effects on soil flux and soil properties. Field Crops Res. 1995, 40, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Scheunemann, N.; Maraun, M.; Scheu, S.; Butenschoen, O. The role of shoot residues vs. crop species for soil arthropod diversity and abundance of arable systems. Soil Biol. Biochem. 2015, 81, 81–88. [Google Scholar] [CrossRef]
- Brevik, E.; Cerdà, A.; Mataix-Solera, J.; Pereg, L.; Quinton, J.; Six, J.; Van Oost, K. The interdisciplinary nature of SOIL. Soil 2015, 1, 117–129. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.; Fu, G.; White, L.W. Complementary teaching approaches facilitating interdisciplinary soil science education. Can. J. Soil Sci. 2021, 102, 433–443. [Google Scholar] [CrossRef]
- Hou, D.; Bolan, N.S.; Tsang, D.C.; Kirkham, M.B.; O’Connor, D. Sustainable soil use and management: An interdisciplinary and systematic approach. Sci. Total Environ. 2020, 729, 138961. [Google Scholar] [CrossRef]
- Wahl, J.; Theron, P.; Maboeta, M. Soil mesofauna as bioindicators to assess environmental disturbance at a platinum mine in South Africa. Ecotoxicol. Environ. Saf. 2012, 86, 250–260. [Google Scholar] [CrossRef]
- Endlweber, K.; Scheu, S. Interactions between mycorrhizal fungi and Collembola: Effects on root structure of competing plant species. Biol. Fertil. Soils 2007, 43, 741–749. [Google Scholar] [CrossRef]
- Menta, C. Soil fauna diversity-function, soil degradation, biological indices, soil restoration. Biodivers. Conserv. Util. A Divers. World 2012. [Google Scholar] [CrossRef]
- Rzeszowski, K.; Zadrożny, P.; Nicia, P. The effect of soil nutrient gradients on Collembola communities inhabiting typical urban green spaces. Pedobiologia 2017, 64, 15–24. [Google Scholar] [CrossRef]
- Kitching, R.L.; Dahlsjö, C.A.; Eggleton, P. Invertebrates and the complexity of tropical ecosystems. Biotropica 2020, 52, 207–214. [Google Scholar] [CrossRef]
- Coulibaly, S.F.; Winck, B.R.; Akpa-Vinceslas, M.; Mignot, L.; Legras, M.; Forey, E.; Chauvat, M.J.A.S.E. Functional assemblages of Collembola determine soil microbial communities and associated functions. Front. Environ. Sci. 2019, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Winck, B.R.; Chauvat, M.; Coulibaly, S.F.; Santonja, M.; Saccol de Sá, E.L.; Forey, E. Functional collembolan assemblages induce different plant responses in Lolium perenne. Plant Soil 2020, 452, 347–358. [Google Scholar] [CrossRef]
- Frampton, G.K. The potential of Collembola as indicators of pesticide usage: Evidence and methods from the UK arable ecosystem. Pedobiologia 1997, 41, 179–184. [Google Scholar]
- Kopeszki, H. An active bioindication method for the diagnosis of soil properties using Collembola. Pedobiologia 1997, 41, 159–166. [Google Scholar]
- Van Straalen, N.M.; Verhoef, H.A. The development of a bioindicator system for soil acidity based on arthropod pH preferences. J. Appl. Ecol. 1997, 34, 217–232. [Google Scholar] [CrossRef]
- Andersen, A. Plant protection in spring cereal production with reduced tillage. II. Pests and beneficial insects. Crop Prot. 1999, 18, 651–657. [Google Scholar] [CrossRef]
- Bockus, W.; Shroyer, J. The impact of reduced tillage on soilborne plant pathogens. Annu. Rev. Phytopathol. 1998, 36, 485–500. [Google Scholar] [CrossRef]
- Vanhée, B.; Devigne, C. Differences in collembola species assemblages (Arthropoda) between spoil tips and surrounding environments are dependent on vegetation development. Sci. Rep. 2018, 8, 18067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, J.; Pratt, J.R. A history of biological monitoring using benthic macroinvertebrates. Freshw. Biomonitoring Benthic Macroinvertebrates 1993, 10, 27. [Google Scholar]
- Dickens, C.; Graham, P. Biomonitoring for effective management of wastewater discharges and the health of the river environment. Aquat. Ecosyst. Health Manag. 1998, 1, 199–217. [Google Scholar] [CrossRef]
- Carlisle, D.M.; Meador, M.R.; Moulton II, S.R.; Ruhl, P.M. Estimation and application of indicator values for common macroinvertebrate genera and families of the United States. Ecol. Indic. 2007, 7, 22–33. [Google Scholar] [CrossRef]
- Sandler, R.V.; Falco, L.B.; Di Ciocco, C.A.; Castro-Huerta, R.; Coviella, C.E. The Degree of Change of Collembolan Community Structure Related to Anthropic Soil Disturbance; PeerJ PrePrints: London, UK, 2014; pp. 2167–9843. [Google Scholar]
- da Fonseca, J.C.; Sarkar, S. Soil microarthropods in two different managed ecological systems (Tripura, India). Appl. Soil Ecol. 1998, 9, 105–107. [Google Scholar] [CrossRef]
- Cortet, J.; Ronce, D.; Poinsot-Balaguer, N.; Beaufreton, C.; Chabert, A.; Viaux, P.; de Fonseca, J.P.C. Impacts of different agricultural practices on the biodiversity of microarthropod communities in arable crop systems. Eur. J. Soil Biol. 2002, 38, 239–244. [Google Scholar] [CrossRef]
- Dittmer, S.; Schrader, S. Longterm effects of soil compaction and tillage on Collembola and straw decomposition in arable soil. Pedobiologia 2000, 44, 527–538. [Google Scholar] [CrossRef]
- Winter, J.; Voroney, R.; Ainsworth, D. Soil microarthropods in long-term no-tillage and conventional tillage corn production. Can. J. Soil Sci. 1990, 70, 641–653. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H. Effects of non-inverting deep tillage vs. conventional ploughing on collembolan populations in an organic wheat field. Eur. J. Soil Biol. 2002, 38, 177–180. [Google Scholar] [CrossRef]
- Gupta, V.; Roper, M.; Thompson, J.; Pratley, J.; Kirkegaard, J. Harnessing the benefits of soil biology in conservation agriculture. In Australian Agriculture in 2020: From Conservation to Automation; Pratley, J., Kirkegaard, J., Eds.; Agronomy Australia and Charles Sturt University: Wagga Wagga, NSW, Australia; pp. 237–253.
- Wardle, D.A. Impacts of disturbance on detritus food webs in agro-ecosystems of contrasting tillage and weed management practices. In Advances in Ecological Research; Elsevier: Amsterdam, The Netherlands, 1995; Volume 26, pp. 105–185. [Google Scholar]
- Axelsen, J.A.; Kristensen, K.T. Collembola and mites in plots fertilised with different types of green manure. Pedobiologia 2000, 44, 556–566. [Google Scholar] [CrossRef]
- Kladivko, E.J. Tillage systems and soil ecology. Soil Tillage Res. 2001, 61, 61–76. [Google Scholar] [CrossRef]
- Ngosong, C.; Raupp, J.; Scheu, S.; Ruess, L. Low importance for a fungal based food web in arable soils under mineral and organic fertilization indicated by Collembola grazers. Soil Biol. Biochem. 2009, 41, 2308–2317. [Google Scholar] [CrossRef]
- Cole, L.; Buckland, S.; Bardgett, R.D. Influence of disturbance and nitrogen addition on plant and soil animal diversity in grassland. Soil Biol. Biochem. 2008, 40, 505–514. [Google Scholar] [CrossRef]
- Holland, J.M. The environmental consequences of adopting conservation tillage in Europe: Reviewing the evidence. Agric. Ecosyst. Environ. 2004, 103, 1–25. [Google Scholar] [CrossRef]
- Tsiafouli, M.A.; Thébault, E.; Sgardelis, S.P.; De Ruiter, P.C.; Van Der Putten, W.H.; Birkhofer, K.; Hemerik, L.; De Vries, F.T.; Bardgett, R.D.; Brady, M.V. Intensive agriculture reduces soil biodiversity across Europe. Glob. Chang. Biol. 2015, 21, 973–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Capelle, C.; Schrader, S.; Brunotte, J. Tillage-induced changes in the functional diversity of soil biota–A review with a focus on German data. Eur. J. Soil Biol. 2012, 50, 165–181. [Google Scholar] [CrossRef]
- Moos, J.H.; Schrader, S.; Paulsen, H.M. Minor changes in collembolan communities under different organic crop rotations and tillage regimes. Landbauforsch. J. Sustain. Org. Agric. Syst. 2020, 70, 113–128. [Google Scholar]
- Akkerhuis, G.J.O.; De Ley, F.; Zwetsloot, H.; Ponge, J.-F.; Brussaard, L. Soil microarthropods (Acari and Collembola) in two crop rotations on a heavy marine clay soil. Rev. D’ecologie Et De Biol. Du Sol 1988, 25, 175–202. [Google Scholar]
- Rebek, E.; Hogg, D.; Young, D. Effect of four cropping systems on the abundance and diversity of epedaphic springtails (Hexapoda: Parainsecta: Collembola) in southern Wisconsin. Environ. Entomol. 2002, 31, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Gruss, I.; Twardowski, J. The assemblages of soil-dwelling springtails (Collembola) in winter rye under long-term monoculture and crop rotation. Zemdirb. Agric. 2016, 103, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Lawton, J.H. Community Ecology in a Changing World; Oldendorf/Luhe (Germany) Ecology Inst.: Oldendorf/Luhe, Germany, 2000. [Google Scholar]
- Baillie, I. Soil Survey Staff 1999, Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, Agricultural Handbook 436, Natural Resources Conservation Service, USDA, Washington DC, USA; Wiley Online Library: Hoboken, NJ, USA, 2001; p. 869. [Google Scholar]
- Thomson, L.; Neville, P.; Hoffmann, A. Effective trapping methods for assessing invertebrates in vineyards. Aust. J. Exp. Agric. 2004, 44, 947–953. [Google Scholar] [CrossRef]
- Buchholz, J.; Querner, P.; Paredes, D.; Bauer, T.; Strauss, P.; Guernion, M.; Scimia, J.; Cluzeau, D.; Burel, F.; Kratschmer, S. Soil biota in vineyards are more influenced by plants and soil quality than by tillage intensity or the surrounding landscape. Sci. Rep. 2017, 7, 17445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melbourne, B.A. Bias in the effect of habitat structure on pitfall traps: An experimental evaluation. Aust. J. Ecol. 1999, 24, 228–239. [Google Scholar] [CrossRef] [Green Version]
- Fiera, C.; Ulrich, W.; Popescu, D.; Buchholz, J.; Querner, P.; Bunea, C.-I.; Strauss, P.; Bauer, T.; Kratschmer, S.; Winter, S. Tillage intensity and herbicide application influence surface-active springtail (Collembola) communities in Romanian vineyards. Agric. Ecosyst. Environ. 2020, 300, 107006. [Google Scholar] [CrossRef]
- Betsch, J.-M. Éléments pour une monographie des collemboles symphyplèones (hexapodes, aptérygotes). Éditions Du Muséum 1980, 116, 1–227. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola: Symphypleona. Abh. Und Ber. Des Nat. Gorlitz. 1999, 71, 1–318. [Google Scholar]
- Potapov, M. Synopses on Palaearctic Collembola: Isotomidae. Abh. Und Ber. Des Nat. Gorlitz. 2001, 73, 1–603. [Google Scholar]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: http://www.collembola.org (accessed on 11 May 2021).
- Vandewalle, M.; De Bello, F.; Berg, M.P.; Bolger, T.; Doledec, S.; Dubs, F.; Feld, C.K.; Harrington, R.; Harrison, P.A.; Lavorel, S. Functional traits as indicators of biodiversity response to land use changes across ecosystems and organisms. Biodivers. Conserv. 2010, 19, 2921–2947. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.L.; Bouffaud, M.-L.; Buée, M.; Dimmers, W. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [Green Version]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Newman, M.E. The structure and function of complex networks. SIAM Rev. 2003, 45, 167–256. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [Green Version]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open Source Software for Exploring and Manipulating Networks. In Proceedings of the Third International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Erdos, P.; Rényi, A. On the evolution of random graphs. Publ. Math. Inst. Hung. Acad. Sci 1960, 5, 17–60. [Google Scholar]
- Degrune, F.; Theodorakopoulos, N.; Colinet, G.; Hiel, M.-P.; Bodson, B.; Taminiau, B.; Daube, G.; Vandenbol, M.; Hartmann, M. Temporal dynamics of soil microbial communities below the seedbed under two contrasting tillage regimes. Front. Microbiol. 2017, 8, 1127. [Google Scholar] [CrossRef]
- Pausch, J.; Hofmann, S.; Scharroba, A.; Kuzyakov, Y.; Ruess, L. Fluxes of root-derived carbon into the nematode micro-food web of an arable soil. Food Webs 2016, 9, 32–38. [Google Scholar] [CrossRef]
- Spedding, T.; Hamel, C.; Mehuys, G.; Madramootoo, C. Soil microbial dynamics in maize-growing soil under different tillage and residue management systems. Soil Biol. Biochem. 2004, 36, 499–512. [Google Scholar] [CrossRef]
- Zhang, B.; He, H.; Ding, X.; Zhang, X.; Zhang, X.; Yang, X.; Filley, T.R. Soil microbial community dynamics over a maize (Zea mays L.) growing season under conventional-and no-tillage practices in a rainfed agroecosystem. Soil Tillage Res. 2012, 124, 153–160. [Google Scholar] [CrossRef]
- Six, J.; Ogle, S.M.; Jay Breidt, F.; Conant, R.T.; Mosier, A.R.; Paustian, K. The potential to mitigate global warming with no-tillage management is only realized when practised in the long term. Glob. Chang. Biol. 2004, 10, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Chen, X.; Jia, S.; Liang, A.; Zhang, X.; Yang, X.; Wei, S.; Sun, B.; Huang, D.; Zhou, G. The potential mechanism of long-term conservation tillage effects on maize yield in the black soil of Northeast China. Soil Tillage Res. 2015, 154, 84–90. [Google Scholar] [CrossRef]
- Cole, L.J.; McCracken, D.I.; Foster, G.N.; Aitken, M.N. Using Collembola to assess the risks of applying metal-rich sewage sludge to agricultural land in western Scotland. Agric. Ecosyst. Environ. 2001, 83, 177–189. [Google Scholar] [CrossRef]
- Loring, S.; Snider, R.J.; Robertson, L.S. The effects of three tillage practices on Collembola and Acarina populations. Pedobiologia 1981, 22, 172–184. [Google Scholar]
- Brennan, A.; Fortune, T.; Bolger, T. Collembola abundances and assemblage structures in conventionally tilled and conservation tillage arable systems. Pedobiologia 2006, 50, 135–145. [Google Scholar] [CrossRef]
- Govaerts, B.; Mezzalama, M.; Sayre, K.D.; Crossa, J.; Nicol, J.M.; Deckers, J. Long-term consequences of tillage, residue management, and crop rotation on maize/wheat root rot and nematode populations in subtropical highlands. Appl. Soil Ecol. 2006, 32, 305–315. [Google Scholar] [CrossRef]
- Sabatini, M.A.; Rebecchi, L.; Cappi, C.; Bertolani, R.; Fratello, B. Long-term effects of three different continuous tillage practices on Collembola populations. Pedobiologia 1997, 41, 185–193. [Google Scholar]
- MooRE, J.C.; Snider, R.J.; Robertson, L.S. Effects of different management practices on Collembola and Acarina in corn production systems. I: The effects of no-tillage and atrazine. Pedobiologia 1984, 26, 143–152. [Google Scholar]
- de Oliveira Filho, L.C.I.; Zeppelini, D.; Sousa, J.P.; Baretta, D.; Klauberg-Filho, O. Collembola community structure under different land management in subtropical Brazil. Ann. Appl. Biol. 2020, 177, 294–307. [Google Scholar] [CrossRef]
- Baretta, D.; Ferreira, C.S.; Sousa, J.P.; Cardoso, E.J.B.N. Colêmbolos (Hexapoda: Collembola) como bioindicadores de qualidade do solo em áreas com Araucaria angustifolia. Rev. Bras. De Ciência Do Solo 2008, 32, 2693–2699. [Google Scholar] [CrossRef]
- Pflug, A.; Wolters, V. Collembola communities along a European transect. Eur. J. Soil Biol. 2002, 38, 301–304. [Google Scholar] [CrossRef]
- Rożen, A.; Sobczyk, Ł.; Liszka, K.; Weiner, J. Soil faunal activity as measured by the bait-lamina test in monocultures of 14 tree species in the Siemianice common-garden experiment, Poland. Appl. Soil Ecol. 2010, 45, 160–167. [Google Scholar] [CrossRef]
- Schröder, P. Mesofauna. In Perspectives for Agroecosystem Management; Elsevier: Amsterdam, The Netherlands, 2008; pp. 293–306. [Google Scholar]
- Culik, M.P.; Martins, D.d.S.; Ventura, J.A. Collembola (Arthropoda: Hexapoda) communities in the soil of papaya orchards managed with conventional and integrated production in Espírito Santo, Brazil. Biota Neotrop. 2006, 6, No. 3. [Google Scholar] [CrossRef] [Green Version]
- Nakamoto, T.; Yamagishi, J.; Miura, F. Effect of reduced tillage on weeds and soil organisms in winter wheat and summer maize cropping on Humic Andosols in Central Japan. Soil Tillage Res. 2006, 85, 94–106. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Milcu, A.; Sabais, A.C.; Bessler, H.; Brenner, J.; Engels, C.; Klarner, B.; Maraun, M.; Partsch, S.; Roscher, C. Plant diversity surpasses plant functional groups and plant productivity as driver of soil biota in the long term. PLoS ONE 2011, 6, e16055. [Google Scholar] [CrossRef]
- Milcu, A.; Partsch, S.; Langel, R.; Scheu, S. The response of decomposers (earthworms, springtails and microorganisms) to variations in species and functional group diversity of plants. Oikos 2006, 112, 513–524. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; WEIBULL, A.C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Jeanneret, P.; Schüpbach, B.; Luka, H. Quantifying the impact of landscape and habitat features on biodiversity in cultivated landscapes. Agric. Ecosyst. Environ. 2003, 98, 311–320. [Google Scholar] [CrossRef]
- Heiniger, C.; Barot, S.; Ponge, J.-F.; Salmon, S.; Meriguet, J.; Carmignac, D.; Suillerot, M.; Dubs, F. Collembolan preferences for soil and microclimate in forest and pasture communities. Soil Biol. Biochem. 2015, 86, 181–192. [Google Scholar] [CrossRef]
- Querner, P.; Bruckner, A.; Drapela, T.; Moser, D.; Zaller, J.; Frank, T. Landscape and site effects on Collembola diversity and abundance in winter oilseed rape fields in eastern Austria. Agric. Ecosyst. Environ. 2013, 164, 145–154. [Google Scholar] [CrossRef]
- Pfingstmann, A.; Paredes, D.; Buchholz, J.; Querner, P.; Bauer, T.; Strauss, P.; Kratschmer, S.; Winter, S.; Zaller, J. Contrasting effects of tillage and landscape structure on spiders and springtails in vineyards. Sustainability 2019, 11, 2095. [Google Scholar] [CrossRef] [Green Version]
- Joimel, S.; Schwartz, C.; Hedde, M.; Kiyota, S.; Krogh, P.H.; Nahmani, J.; Pérès, G.; Vergnes, A.; Cortet, J. Urban and industrial land uses have a higher soil biological quality than expected from physicochemical quality. Sci. Total Environ. 2017, 584, 614–621. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Abundance | Richness | ||||||
|---|---|---|---|---|---|---|---|---|
| Weak Furca | Strong Furca | Weak Furca | Strong Furca | |||||
| F | P | F | p | F | p | F | p | |
| Til | 0.024 | 0.976 | 3.891 | 0.039 | 0.324 | 0.727 | 0.656 | 0.531 |
| Mag | 4.198 | 0.055 | 0.164 | 0.69 | 1.362 | 0.258 | 0.057 | 0.813 |
| Til × Mag | 1.062 | 0.366 | 2.565 | 0.105 | 0.676 | 0.521 | 0.248 | 0.783 |
| Factors | Shanon | Simpson | Pielou | |||
|---|---|---|---|---|---|---|
| F | p | F | P | F | P | |
| Til | 0.262 | 0.773 | 0.364 | 0.7 | 0.09 | 0.914 |
| Mag | 0.355 | 0.558 | 0.385 | 0.543 | 0.887 | 0.359 |
| Til × Mag | 0.402 | 0.675 | 0.679 | 0.52 | 1.968 | 0.169 |
| Treatments | Nodes Number | Edges Number | Modularity (MD) | Clustering Coefficient (cc) | Average Path Length (APL) | Network Diameter (NA) | Graph Density (GD) | Average Degree (AD) |
|---|---|---|---|---|---|---|---|---|
| MT-MC-SR | 17 | 22 | 0.459 | 0.556 | 1.5 | 3 | 0.162 | 1.294 |
| MT-CS-SR | 19 | 20 | 0.609 | 0.551 | 2.418 | 6 | 0.117 | 1.053 |
| RT-MC-SR | 25 | 72 | 0.341 | 0.629 | 2.091 | 4 | 0.24 | 2.88 |
| RT-CS-SR | 22 | 49 | 0.397 | 0.54 | 2.519 | 6 | 0.212 | 2.227 |
| NT-MC-SR | 19 | 32 | 0.482 | 0.549 | 2.62 | 5 | 0.187 | 1.684 |
| NT-CS-SR | 25 | 46 | 0.424 | 0.622 | 2.802 | 6 | 0.153 | 1.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habib, T.; Liu, S.; Chang, L.; Wu, Y.; Hao, C.; Wu, D. Investigating the Assemblages of Two Groups of Collembola (Strong Furca and Weak Furca) under Different Agricultural Management Systems, Northeastern China. Diversity 2022, 14, 994. https://doi.org/10.3390/d14110994
Habib T, Liu S, Chang L, Wu Y, Hao C, Wu D. Investigating the Assemblages of Two Groups of Collembola (Strong Furca and Weak Furca) under Different Agricultural Management Systems, Northeastern China. Diversity. 2022; 14(11):994. https://doi.org/10.3390/d14110994
Chicago/Turabian StyleHabib, Tayyiba, Shuchen Liu, Liang Chang, Yunga Wu, Cao Hao, and Donghui Wu. 2022. "Investigating the Assemblages of Two Groups of Collembola (Strong Furca and Weak Furca) under Different Agricultural Management Systems, Northeastern China" Diversity 14, no. 11: 994. https://doi.org/10.3390/d14110994
APA StyleHabib, T., Liu, S., Chang, L., Wu, Y., Hao, C., & Wu, D. (2022). Investigating the Assemblages of Two Groups of Collembola (Strong Furca and Weak Furca) under Different Agricultural Management Systems, Northeastern China. Diversity, 14(11), 994. https://doi.org/10.3390/d14110994