



Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Fauna Samples
2.3. Environmental Variables
2.4. Data Analysis
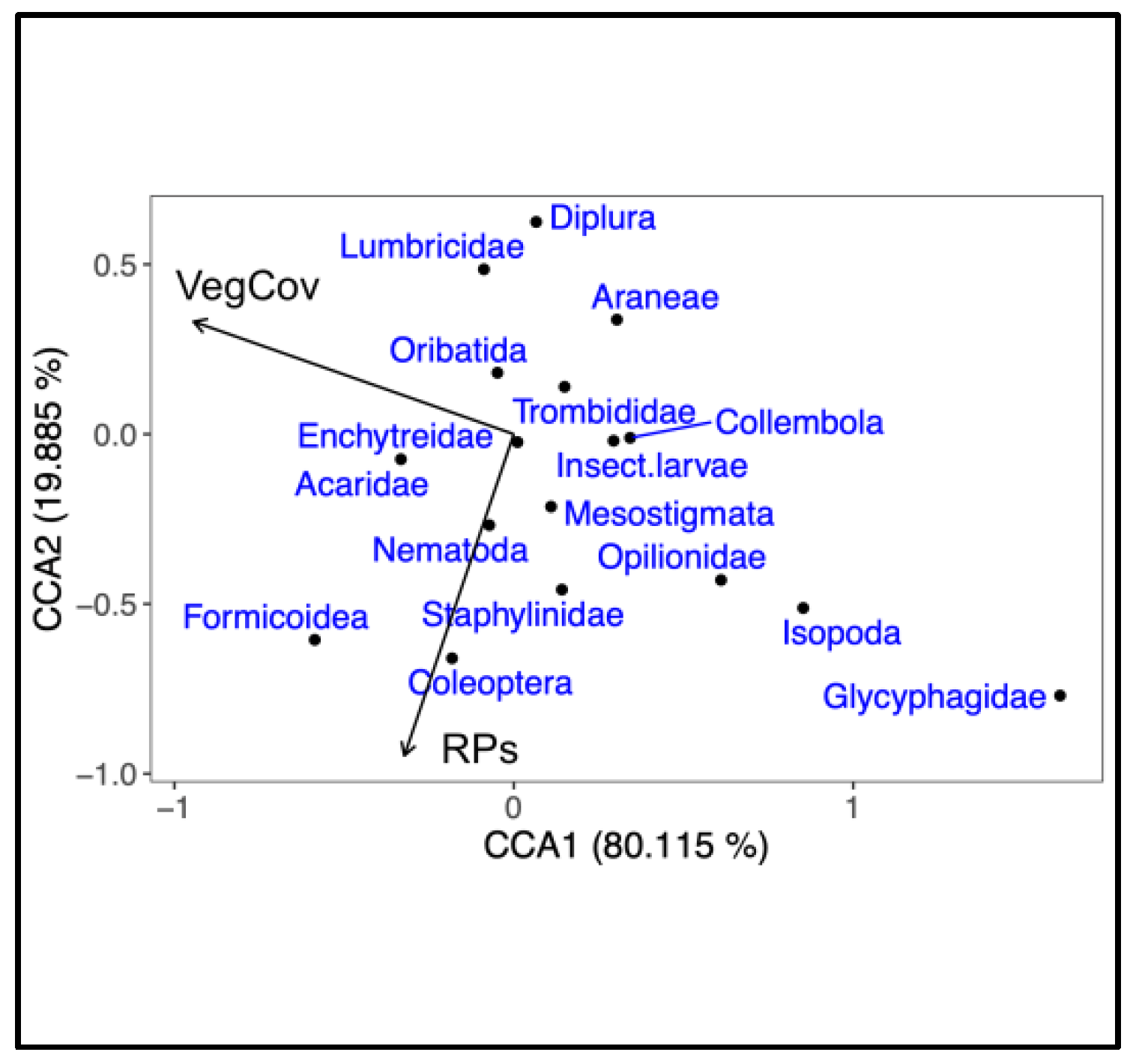
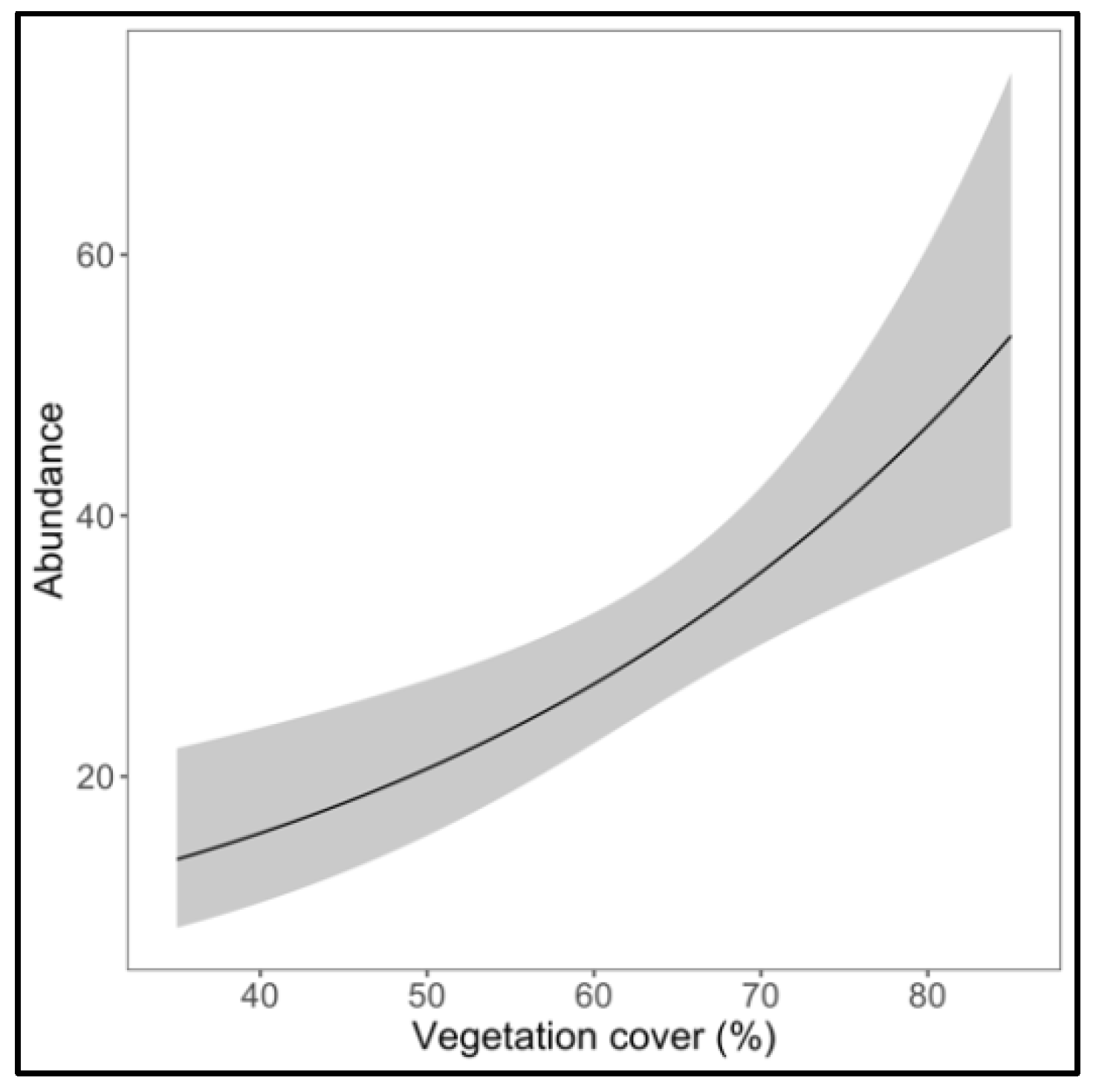
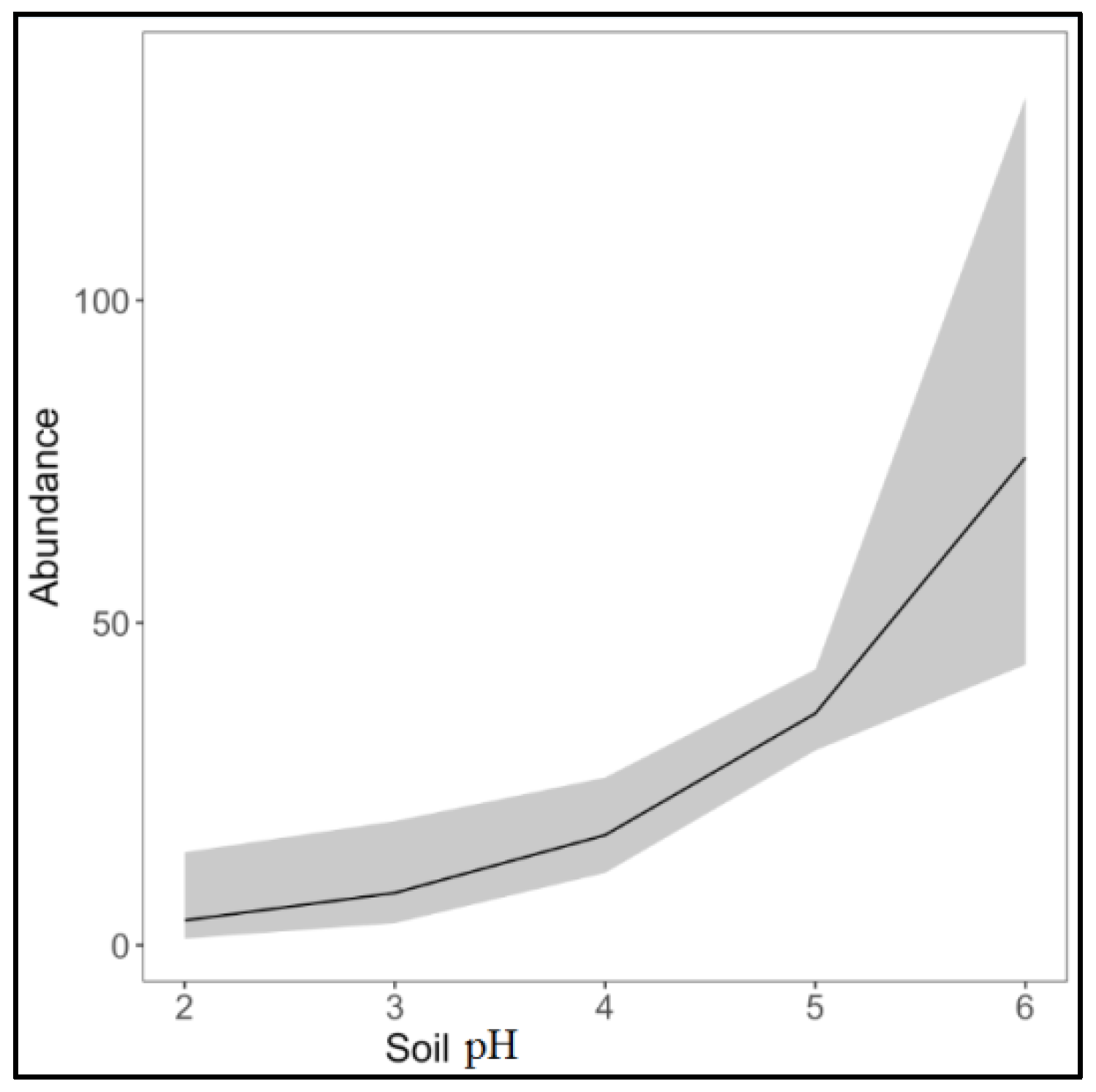
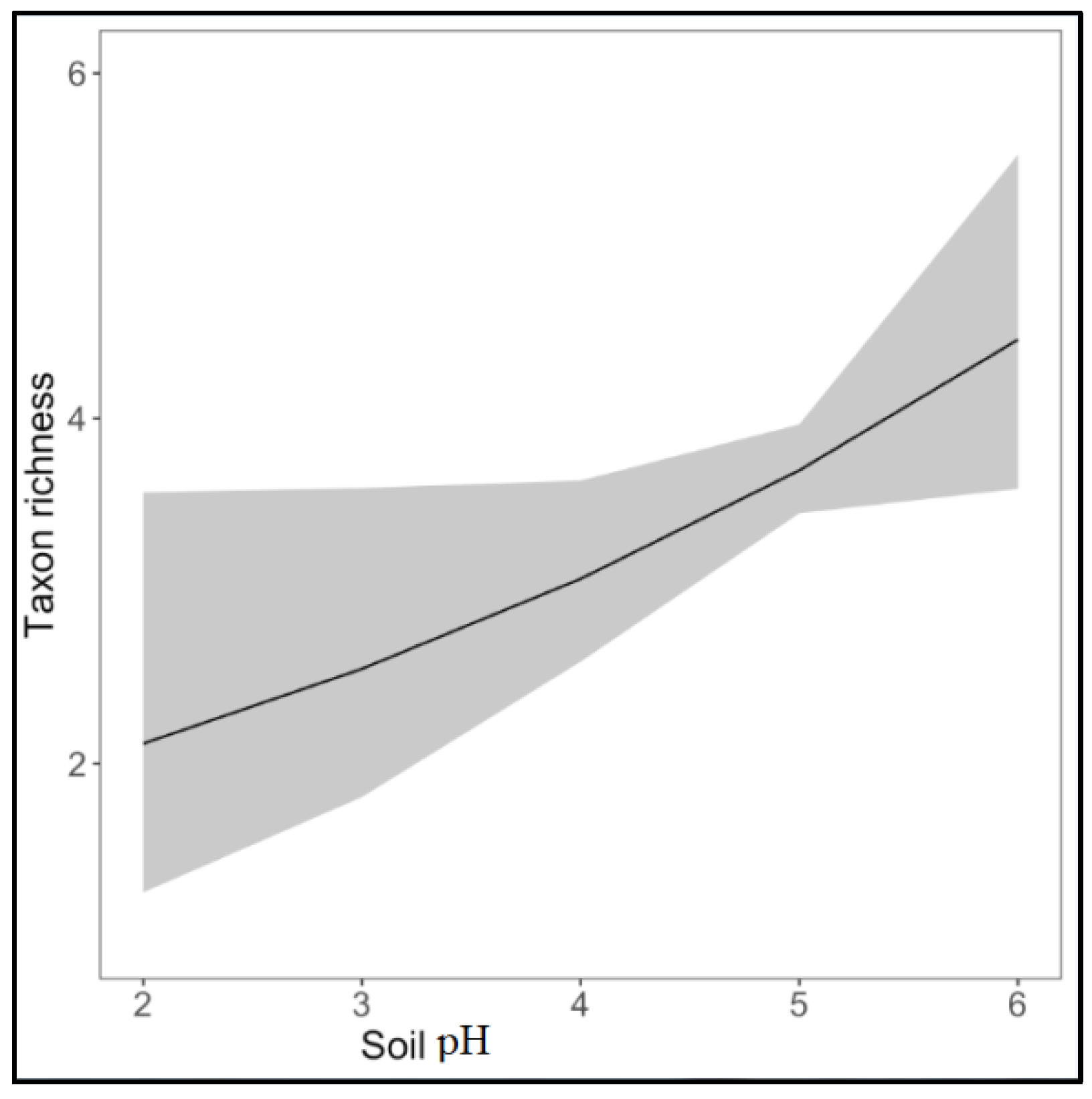
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Curry, J.P. The invertebrate fauna of grassland and its influence on productivity. The composition of the fauna. Grass Forage Sci. 1987, 42, 103–120. [Google Scholar] [CrossRef]
- Menta, C. Soil Fauna Diversity—Function, Soil Degradation, Biological Indices, Soil Restoration. In Biodiversity Conservation and Utilization in a Diverse World; Lameed, G.A., Ed.; IntechOpen: London, UK, 2012; pp. 59–94. [Google Scholar]
- Coleman, D.C.; Wall, D.H. Soil Fauna: Occurrence, Biodiversity, and Roles in Ecosystem Function, In Soil Microbiology, Ecology and Biochemistry, 4th ed.; Eldor, A.P., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 1–41. [Google Scholar]
- Rainio, J.; Niemelä, J. Ground beetles (Coleoptera: Carabidae) as bioindicators. Biodivers. Conserv. 2003, 12, 487–506. [Google Scholar] [CrossRef]
- Fischer, S. Earthworms (Lumbricidae) as bioindicators: The relationship between in-soil and in-tissue heavy metal content. Pol. J. Ecol. 2009, 57, 531–541. [Google Scholar]
- Lemanski, K.; Scheu, S. The influence of fertilizer addition, cutting frequency and herbicide application on soil organisms in grassland. Biol. Fertil. Soils 2014, 51, 197–205. [Google Scholar] [CrossRef]
- Pelosi, C.; Römbke, J. Enchytraeids as bioindicators of land use and management. Appl. Soil Ecol. 2018, 123, 775–779. [Google Scholar] [CrossRef]
- Meehan, M.L.; Song, Z.; Lumley, L.M.; Cobb, T.P.; Proctor, H. Soil mites as bioindicators of disturbance in the boreal forest in northern Alberta, Canada: Testing taxonomic sufficiency at multiple taxonomic levels. Ecol. Indic. 2019, 102, 349–365. [Google Scholar] [CrossRef]
- Crotty, F. Assessing soil health by measuring fauna. In Advances in Measuring Soil Health, 1st ed.; Otten, W., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 1–22. [Google Scholar]
- Van Straalen, N.M. Evaluation of bioindicator systems derived from soil arthropod communities. Appl. Soil Ecol. 1998, 9, 429–437. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Cook, R. Functional aspects of soil animal diversity in agricultural grasslands. Appl. Soil Ecol. 1998, 10, 263–276. [Google Scholar] [CrossRef]
- Tajik, S.; Ayoubi, S.; Shirani, H.; Zeraatpisheh, M. Digital mapping of soil invertebrates using environmental attributes in a deciduous forest ecosystem. Geoderma 2019, 353, 252–263. [Google Scholar] [CrossRef]
- Johnston, A.S.A.; Sibly, R.M. Multiple environmental controls explain global patterns in soil animal communities. Oecologia 2020, 192, 1047–1056. [Google Scholar] [CrossRef] [Green Version]
- Samways, M.J.; McGeoch, M.A.; New, T.R. Insect Conservation: A Handbook of Approaches and Methods; Oxford University Press: Oxford, UK, 2010; 458p. [Google Scholar]
- Báldi, A. Using higher taxa as surrogates of species richness: A study based on Coleoptera, Diptera, and Acari species in Central-Hungarian reserves. Basic Appl. Ecol. 2003, 4, 589–593. [Google Scholar] [CrossRef]
- Ekschmitt, K.; Stierhof, T.; Dauber, J.; Kreimes, K.; Wolters, V. On the quality of soil biodiversity indicators: Abiotic and biotic parameters as predictors of soil faunal richness at different spatial scales. Agric. Ecosyst. Environ. 2003, 98, 273–283. [Google Scholar] [CrossRef]
- Gerlach, J.; Samways, M.; Pryke, J. Terrestrial invertebrates as bioindicators: An overview of available taxonomic groups. J. Insect Conserv. 2013, 17, 831–850. [Google Scholar] [CrossRef]
- Manning, P.; Gossner, M.M.; Bossdorf, O.; Allan, E.; Zhang, Y.-Y.; Prati, D.; Blüthgen, N.; Boch, S.; Böhm, S.; Börschig, C.; et al. Grassland management intensification weakens the associations among the diversities of multiple plant and animal taxa. Ecology 2015, 96, 1492–1501. [Google Scholar] [CrossRef] [Green Version]
- Manu, M.; Bîrsan, C.C.; Mountford, O.; Lăcătuşu, A.R.; Onete, M. Preliminary study on soil fauna as a tool for monitoring of the “Springs Complex of Corbii Ciungi” protected area—Romania. Sci. Pap. Ser. D Anim. Sci. 2020, 63, 272–280. [Google Scholar]
- Manu, M.; Chiriac, L.S.; Onete, M. Structural characteristics of the soil invertebrate communities from two fragmented Natura 2000 sites from Romania. Sci. Pap. Ser. D Anim. Sci. 2021, 64, 529–535. [Google Scholar]
- Pryke, J.S.; Samways, M.J. Importance of using many taxa and having adequate controls for monitoring impacts of fire for arthropod conservation. J. Insect Conserv. 2012, 16, 177–185. [Google Scholar] [CrossRef]
- Nisa, R.U.; Tantray, A.Y.; Kaisar, N.; Allie, A.K.; Wania, S.M.; Alamri, S.A.; Alyemeni, M.N.; Wijaya, L.; Shah, A.A. Influence of ecological and edaphic factors on biodiversity of soil nematodes. Saudi J. Biol. Sci. 2021, 28, 3049–3059. [Google Scholar] [CrossRef]
- Warner, F. Soil fertility, pH, texture and nematodes. Diagn. Serv. 2009, 13, 21. [Google Scholar]
- Šalamún, P.; Renčo, M.; Kucanová, E.; Brázová, T.; Papajová, I.; Miklisová, D.; Hanzelová, V. Nematodes as bioindicators of soil degradation due to heavy metals. Ecotoxicology 2012, 21, 2319–2330. [Google Scholar] [CrossRef]
- Van der Wal, A.; Geerts, R.H.E.M.; Korevaar, H.; Schouten, A.J.; Jagers op Akkerhuis, G.A.J.M.; Rutgers, M.; Mulder, C. Dissimilar response of plant and soil biota communities to long-term nutrient addition in grasslands. Biol. Fertil. Soils 2009, 45, 663–667. [Google Scholar] [CrossRef]
- Briones, M.J.I.; Ineson, P.; Heinemeyer, A. Predicting potential impacts of climate change on the geographical distribution of enchytraeids: A meta-analysis approach. Glob. Chang. Biol. 2007, 13, 2252–2269. [Google Scholar] [CrossRef]
- Rőmbke, J.; Jänsch, S.; Didden, W.A.M. The use of earthworms in ecological soil classification and assessment concepts. Ecotoxicol. Environ. Saf. 2005, 62, 249–265. [Google Scholar] [CrossRef] [PubMed]
- Nahmani, J.; Hodson, M.E.; Devin, S.; Vijver, M.G. Uptake kinetics of metals by the earthworm Eisenia fetida exposed to field-contaminated soils. Environ. Pollut. 2009, 157, 2622–2628. [Google Scholar] [CrossRef] [PubMed]
- Fründ, H.C.; Graefe, U.; Tischer, S. Earthworms as Bioindicators of Soil Quality. In Biology of Earthworms, Soil Biology; Karaca, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 24, pp. 261–278. [Google Scholar]
- Frampton, G.W. Spatial variation in non-target effects of the insecticides chlorpyrifos, cypermethrin and pirimicarb on Collembola in winter wheat. Pesticide Sci. 1999, 55, 875–886. [Google Scholar] [CrossRef]
- Cole, L.; Bradford, M.A.; Shaw, P.J.A.; Bardgett, R.D. The abundance, richness and functional role of soil meso- and macrofauna in temperate grassland—A case study. Appl. Soil Ecol. 2006, 33, 186–198. [Google Scholar] [CrossRef]
- García-Segura, D.; Castillo-Murrieta, I.M.; Martínez-Rabelo, F.; Gomez-Anaya, A.; Rodríguez-Campos, J.; Hernández-Castellanos, B.; Contreras-Ramos, S.M.; Barois, I. Macrofauna and mesofauna from soil contaminated by oil extraction. Geoderma 2018, 332, 180–189. [Google Scholar] [CrossRef]
- Manu, M.; Honciuc, V.; Neagoe, A.; Băncilă, R.I.; Iordache, V.; Onete, M. Soil mite communities (Acari: Mesostigmata, Oribatida) as bioindicators for environmental conditions from polluted soils. Sci. Rep. 2019, 9, 20250. [Google Scholar] [CrossRef] [Green Version]
- Pearce, J.L.; Venier, L.A. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecol. Indic. 2006, 6, 780–793. [Google Scholar] [CrossRef]
- Manu, M.; Băncilă, R.I.; Mountford, O.J.; Maruşca, T.; Blaj, V.A.; Onete, M. Soil Mite (Acari: Mesostigmata) Communities and Their Relationship with Some Environmental Variables in Experimental Grasslands from Bucegi Mountains in Romania. Insects 2022, 13, 285. [Google Scholar] [CrossRef]
- Covaciu-Marcov, S.-D.; Ferenţi, S.; Urák, I.; Sas-Kovács, É.-H.; Cicort-Lucaciu, A.-Ş.; Sas-Kovács, I. After the Last Train Passes: Data on the Fauna from Abandoned Railway Tunnels in Romania. Ann. Zool. Fenn. 2017, 54, 335–346. [Google Scholar] [CrossRef]
- Fiera, C.; Ulrich, W.; Popescu, D.; Bunea, C.I.; Manu, M.; Nae, I.; Stan, M.; Markó, B.; Urák, I.; Giurgincă, A.; et al. Effects of vineyard inter-row management on the diversity and abundance of plants and surface-dwelling invertebrates in Central Romania. J. Insect Conserv. 2020, 24, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Blaj, V.A.; Maruşca, T.; Andreoiu, A.C.; Mocanu, V. The long-term effect on cow’s milk quality of methods for improving subalpine grasslands. In Grassland Resources for Extensive Farming Systems in Marginal Lands: Major Drivers and Future Scenarios, Grassland Science of Europe; Porqueddu, C., Franca, A., Lombardi, G., Molle, G., Peratoner, G., Hopkins, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2017; Volume 22, pp. 109–111. [Google Scholar]
- Stănilă, A.L.; Dumitru, M. Soil zones in Romania and pedogenetic processes. Agric. Agric. Sci. Procedia 2016, 10, 135–139. [Google Scholar] [CrossRef] [Green Version]
- Balogh, J. The Oribatid Genera of the World; Akademiai Kiado: Budapest, Hungary, 1972; 188p. [Google Scholar]
- Gîdei, P.; Popescu, I.E. Îndrumător Pentru Cunoașterea Coleopterelor; Editura Pim: Iași, Romania, 2009; pp. 5–419. [Google Scholar]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; 807p. [Google Scholar]
- Ceuca, T. Diplopoda. In Determinatorul Ilustrat Al Florei Si Faunei Romaniei—Mediul Terestru; Godeanu, S.P., Ed.; Vasile Goldiş University Press: Arad, România, 2010; Volume 3, pp. 290–300. [Google Scholar]
- Platnick, N.I.; World Spider Catalog. World Spider Catalog. Version 19.5. Natural History Museum Bern. 2018. Available online: http://wsc.nmbe.ch (accessed on 5 February 2022).
- Brussaard, L.; Behan-Pelletier, V.M.; Bignell, D.E.; Brown, V.K.; Didde, W.; Folgaria, P.; Fragoso, C.; Freckman, D.W.; Gupta, V.V.S.R.; Hattori, T.; et al. Biodiversity and ecosystem functioning in soil. Ambio. J. Hum. Environ. 1997, 26, 563–570. [Google Scholar]
- Chahartaghi, M.; Langel, R.; Scheu, S.; Ruess, L. Feeding guilds in Collembola based on nitrogen stable isotope ratios. Soil Biol. Biochem. 2005, 37, 1718–1725. [Google Scholar] [CrossRef]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the Spatial Component of Ecological Variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L. Numerical Ecology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 1998; 1006p. [Google Scholar]
- Ter Braak, J.F.C. Canonical Correspondence Analysis: A new eigenvector technique for Multivariate Direct Gradient Analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, L.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Community Ecology Package. R Package Version 2.5-2. October 2018. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 12 May 2022).
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Interference. A Practical Information—Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; 515p. [Google Scholar]
- Johnson, J.B.; Omland, K.S. Model selection in ecology and evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.0-3. Available online: http://CRAN.R-project.org/package=AICcmodavg (accessed on 10 May 2022).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 15 May 2022).
- Birkhofer, K.; Dietrich, C.; John, K.; Schorpp, Q.; Zaitsev, A.S.; Wolters, V. Regional Conditions and Land-Use Alter the Potential Contribution of Soil Arthropods to Ecosystem Services in Grasslands. Front. Ecol. Evol. 2016, 3, 150. [Google Scholar] [CrossRef] [Green Version]
- Briones, M.J.I.; Ostle, N.J.; McNamara, N.P.; Poskitt, J. Functional shifts of grassland soil communities in response to soil warming. Soil Biol. Biochem. 2009, 41, 315–322. [Google Scholar] [CrossRef]
- García, R.R.; Ocharan, F.J.; García, U.; Osoro, K.; Celaya, R. Arthropod fauna on grassland–heathland associations under different grazing managements with domestic ruminants. Comptes Rendus Biol. 2010, 333, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Seeber, J.; Seeber, G.U.H.; Kössler, W.; Langel, R.; Scheu, S.; Meyer, E. Abundance and trophic structure of macro-decomposers on alpine pastureland (Central Alps, Tyrol): Effects of abandonment of pasturing. Pedobiologia 2005, 49, 221–228. [Google Scholar] [CrossRef]
- Vandegehuchte, M.L.; Raschein, U.; Schütz, M.; Gwiazdowicz, D.J.; Risch, A.C. Indirect Short- and Long-Term Effects of Aboveground Invertebrate and Vertebrate Herbivores on Soil Microarthropod Communities. PLoS ONE 2015, 10, e0118679. [Google Scholar] [CrossRef] [PubMed]
- Menta, C.; Leoni, A.; Gardi, C.; Conti, F.D. Are grasslands important habitats for soil microarthropod conservation? Biodivers. Conserv. 2011, 20, 1073–1087. [Google Scholar] [CrossRef]
- Gongalsky, K.B. Soil macrofauna: Study problems and perspectives. Soil Biol. Biochem. 2021, 159, 108281. [Google Scholar] [CrossRef]
- Spehn, E.M.; Joshi, J.; Schmid, B.; Alphei, J.; Körner, C. Plant diversity effects on soil heterotrophic activity in experimental grassland ecosystems. Plant Soil 2000, 224, 217–230. [Google Scholar] [CrossRef]
- Schon, N.L.; Mackay, A.D.; Minor, M.A. Effects of dairy cow treading pressures and food resources on invertebrates in two contrasting and co-occurring soils. Soil Res. 2011, 49, 703–714. [Google Scholar] [CrossRef]
- Skuodiene, R.; Tomchuk, D.; Kinderiene, I. The influence of natural and anthropogenic conditions on the earthworm population in different grassland ecosystems. Agriculture 2019, 106, 19–226. [Google Scholar] [CrossRef] [Green Version]
- Laška, V.; Kopecký, O.; Růžička, V.; Mikula, J.; Véle, A.; Šarapatka, B.; Tuf, I.H. Vertical distribution of spiders in soil. J. Arachnol. 2011, 39, 393–398. [Google Scholar] [CrossRef]
- Pompeo, P.N.; Filho, L.C.I.O.; Klauberg Filho, O.; Mafra, Á.L.; Baretta, D. Coleoptera Diversity and Soil Properties in Land Use Systems. Floresta Ambiente 2020, 27, e20180068. [Google Scholar] [CrossRef]
- Qodri, A.; Raffiudin, R.; Noerdjito, W.A. Diversity and Abundance of Carabidae and Staphylinidae (Insecta: Coleoptera) in Four Montane Habitat Types on Mt. Bawakaraeng, South Sulawesi. Hayati J. Biosci. 2016, 23, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Dixie, B.; White, H.; Hassall, M. Effects of microclimate on behavioural and life history traits of terrestrial isopods: Implications for responses to climate change. Zookeys 2015, 515, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Benevenute, P.A.N.; Morais, E.G.; de Souza, A.A.; Vasques, I.C.F.; Cardoso, D.P.; Sales, F.R.; Severiano, E.C.; Homem, B.C.G.; Casagrande, D.R.; Silva, B.M. Penetration resistance: An effective indicator for monitoring soil compaction in pastures. Ecol. Indic. 2020, 117, 106647. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Wani, A.B.; Dhanjal, D.S.; Romero, R.; Singh, J. Glyphosate uptake, translocation, resistance emergence in crops, analytical monitoring, toxicity and degradation: A review. Environ. Chem. Lett. 2020, 18, 663–702. [Google Scholar] [CrossRef]
- Duke, S.O. Glyphosate: Environmental fate and impact. Weed Sci. 2020, 68, 201–207. [Google Scholar] [CrossRef]
- Zaller, J.G.; Weber, M.; Maderthaner, M.; Gruber, E.; Takács, E.; Mörtl, M.; Klátyik, S.; Győri, J.; Römbke, J.; Leisch, F.; et al. Effects of glyphosate-based herbicides and their active ingredients on earthworms, water infiltration and glyphosate leaching are influenced by soil properties. Environ. Sci. Eur. 2021, 33, 51. [Google Scholar] [CrossRef]
- Santos, M.J.G.; Morgado, R.; Ferreira, N.G.C.; Soares, A.M.V.M.; Loureiro, S. Evaluation of the joint effect of glyphosate and dimethoate using a small-scale terrestrial ecosystem. Ecotoxicol. Environ. Saf. 2011, 74, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Hagner, M.; Mikola, J.; Saloniemi, I.; Saikkonen, K.; Helander, M. Effects of a glyphosate-based herbicide on soil animal trophic groups and associated ecosystem functioning in a northern agricultural field. Sci. Rep. 2019, 9, 8540. [Google Scholar] [CrossRef] [Green Version]
- Lemic, D.; Pajač Živković, I.; Posarić, M.; Bažok, R. Influence of Pre-Sowing Operations on Soil-Dwelling Fauna in Soybean Cultivation. Agriculture 2021, 11, 474. [Google Scholar] [CrossRef]
- Chiriac, L.S.; Murariu, D. Plant–soil fauna interaction—Bioindicators of soil properties in agroecosystems. Sci. Pap. Ser. A Agron. 2021, 64, 39–49. [Google Scholar]
- Ponge, J.F. Food resources and diets of soil animals in a small area of Scots pine litter. Geoderma 1991, 49, 33–62. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, A.C.C.; Oliveira, O.C.C.; Silva, R.A.; Albuquerque, P.M.C.; Rebêlo, J.M.M.; Viana, J.H.; Siqueira, G.M. Abundance and diversity of beetles (Insecta: Coleoptera) in land use and management systems. Rev. Bras. Cienc. Solo. 2020, 44, e0190183. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | CG (N = 50) | A (N = 50) | B (N = 50) | C (N = 50) | D (N = 50) | Total (N = 250) | p Value |
|---|---|---|---|---|---|---|---|
| VegCovr | 70.360 (5.992) | 75.960 (8.748) | 60.540 (10.514) | 57.280 (8.593) | 66.160 (8.665) | 66.060 (10.871) | <0.001 |
| 60.000–80.000 | 50.000–85.000 | 35.000–70.000 | 50.000–72.000 | 50.000–80.000 | 35.000–85.000 | ||
| RPs | 1.352 (0.163) | 1.309 (0.139) | 1.394 (0.149) | 1.327 (0.119) | 1.340 (0.173) | 1.344 (0.151) | 0.06 |
| 1.034–1.861 | 0.965–1.723 | 1.034–1.723 | 1.034–1.585 | 1.034–1.723 | 0.965–1.861 | ||
| Ts | 15.842 (2.124) | 16.746 (1.918) | 16.868 (2.917) | 16.980 (1.624) | 14.656 (1.745) | 16.218 (2.276) | <0.001 |
| 12.400–19.600 | 14.300–21.700 | 13.200–26.500 | 14.000–22.000 | 14.000–22.000 | 11.800–26.500 | ||
| Rhs | 65.260 (8.741) | 68.976 (6.820) | 63.964 (7.683) | 61.490 (5.902) | 63.230 (5.578) | 64.584 (7.424) | <0.001 |
| 47.900–82.400 | 52.000–83.500 | 43.400–80.500 | 49.800–76.900 | 49.800–75.000 | 43.400–83.500 | ||
| pH | 4.654 (0.139) | 4.980 (0.266) | 4.553 (0.219) | 5.053 (0.421) | 4.975 (0.300) | 4.843 (0.346) | <0.001 |
| 4.480–4.990 | 4.520–5.260 | 4.220–5.180 | 4.670–6.200 | 4.420–5.450 | 4.220–6.200 |
| Taxon | Short Name | CG | A | B | C | D | Total Nr. Ab. |
|---|---|---|---|---|---|---|---|
| Nematoda | Nematoda | 8 | 14 | 22 | |||
| Enchytraeidae | Enchytrd | 37 | 54 | 13 | 37 | 21 | 162 |
| Lumbricidae | Lumbricd | 22 | 105 | 19 | 22 | 11 | 179 |
| Collembola | Collembl | 1176 | 889 | 124 | 1180 | 1207 | 4576 |
| Diplura | Diplura | 5 | 8 | 3 | 16 | ||
| Mesostigmata | Messtgmt | 273 | 204 | 52 | 273 | 584 | 1386 |
| Acaridae | Acaridae | 756 | 2250 | 357 | 756 | 679 | 4798 |
| Glycyphagidae | Glycyphg | 8 | 8 | ||||
| Oribatida | Oribatid | 693 | 1110 | 447 | 693 | 716 | 3659 |
| Trombidiidae | Trombidd | 3 | 3 | 6 | |||
| Opiliones | Opiliond | 1 | 1 | 2 | |||
| Araneae | Araneae | 3 | 3 | ||||
| Isopoda | Isopoda | 45 | 45 | ||||
| Formicoidae | Formicod | 3 | 3 | 6 | |||
| Coleoptera | Coleoptr | 1 | 1 | ||||
| Staphylinidae | Stphlnd | 4 | 5 | 4 | 22 | 35 | |
| Insect larva | Inselarv | 9 | 8 | 6 | 9 | 17 | 49 |
| Total no. taxa | 10 | 10 | 11 | 10 | 13 | 17 | |
| Total nr. ab. | 2974 | 4638 | 1033 | 2978 | 3330 | 14,953 | |
| Shannon_H | 1.401 | 1.321 | 1.371 | 1.4 | 1.547 | ||
| Equitability_J | 0.608 | 0.573 | 0.571 | 0.608 | 0.603 |
| Grassland | Taxon | IndVal | p |
|---|---|---|---|
| A | Lumbricidae | 32.1 | <0.001 |
| Acaridae | 27.0 | <0.01 | |
| Oribatida | 22.0 | <0.01 | |
| B | Araneae | 18.6 | <0.05 |
| D | Staphylinidae | 35.1 | <0.001 |
| Mesostigmata | 27.2 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manu, M.; Băncilă, R.I.; Mountford, O.J.; Onete, M. Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania. Diversity 2022, 14, 1031. https://doi.org/10.3390/d14121031
Manu M, Băncilă RI, Mountford OJ, Onete M. Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania. Diversity. 2022; 14(12):1031. https://doi.org/10.3390/d14121031
Chicago/Turabian StyleManu, Minodora, Raluca I. Băncilă, Owen J. Mountford, and Marilena Onete. 2022. "Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania" Diversity 14, no. 12: 1031. https://doi.org/10.3390/d14121031
APA StyleManu, M., Băncilă, R. I., Mountford, O. J., & Onete, M. (2022). Soil Invertebrate Communities as Indicator of Ecological Conservation Status of Some Fertilised Grasslands from Romania. Diversity, 14(12), 1031. https://doi.org/10.3390/d14121031