

Deep-Sea Tardigrades from the North-Western Pacific, with Descriptions of Two New Species

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
3. Results
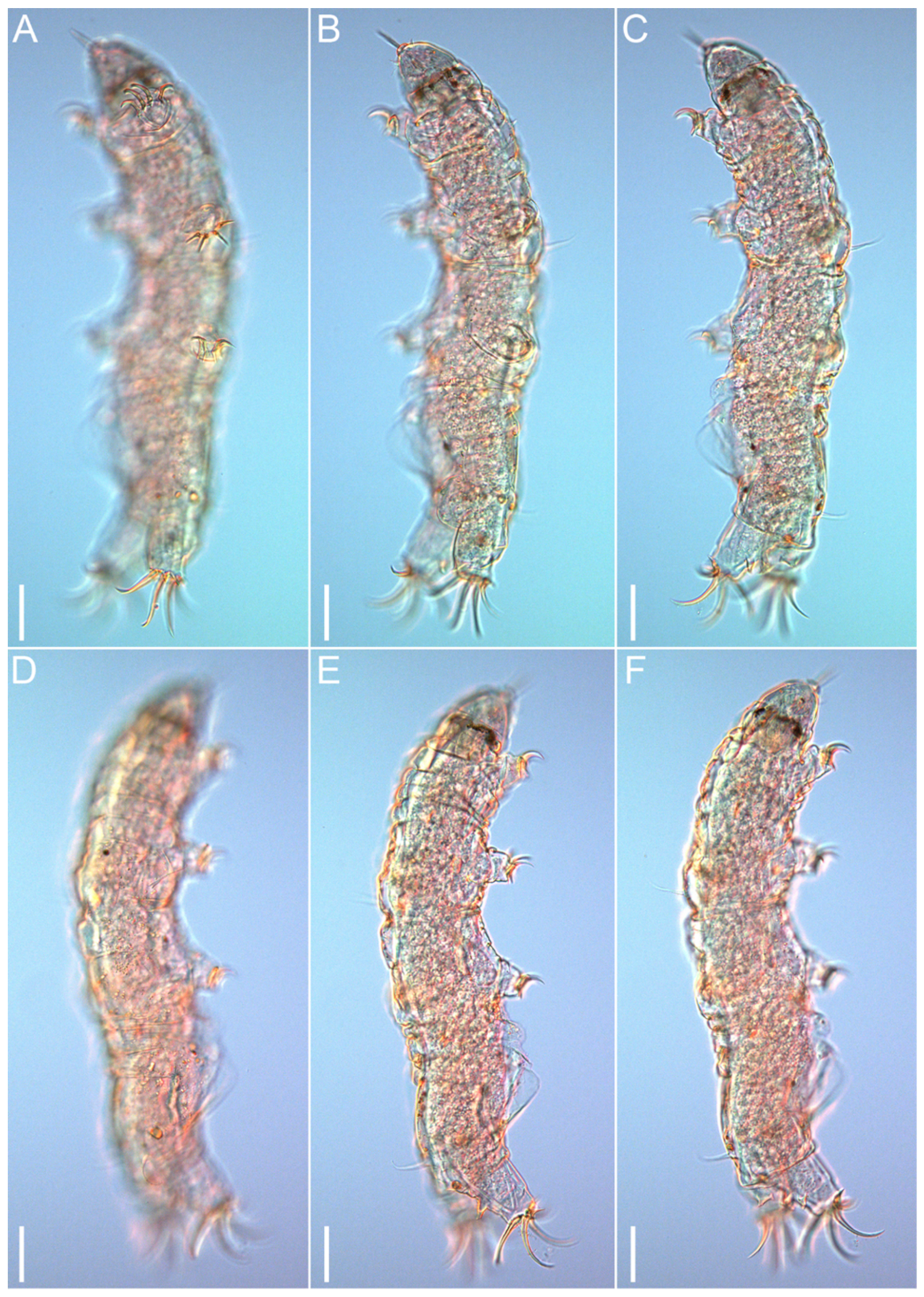
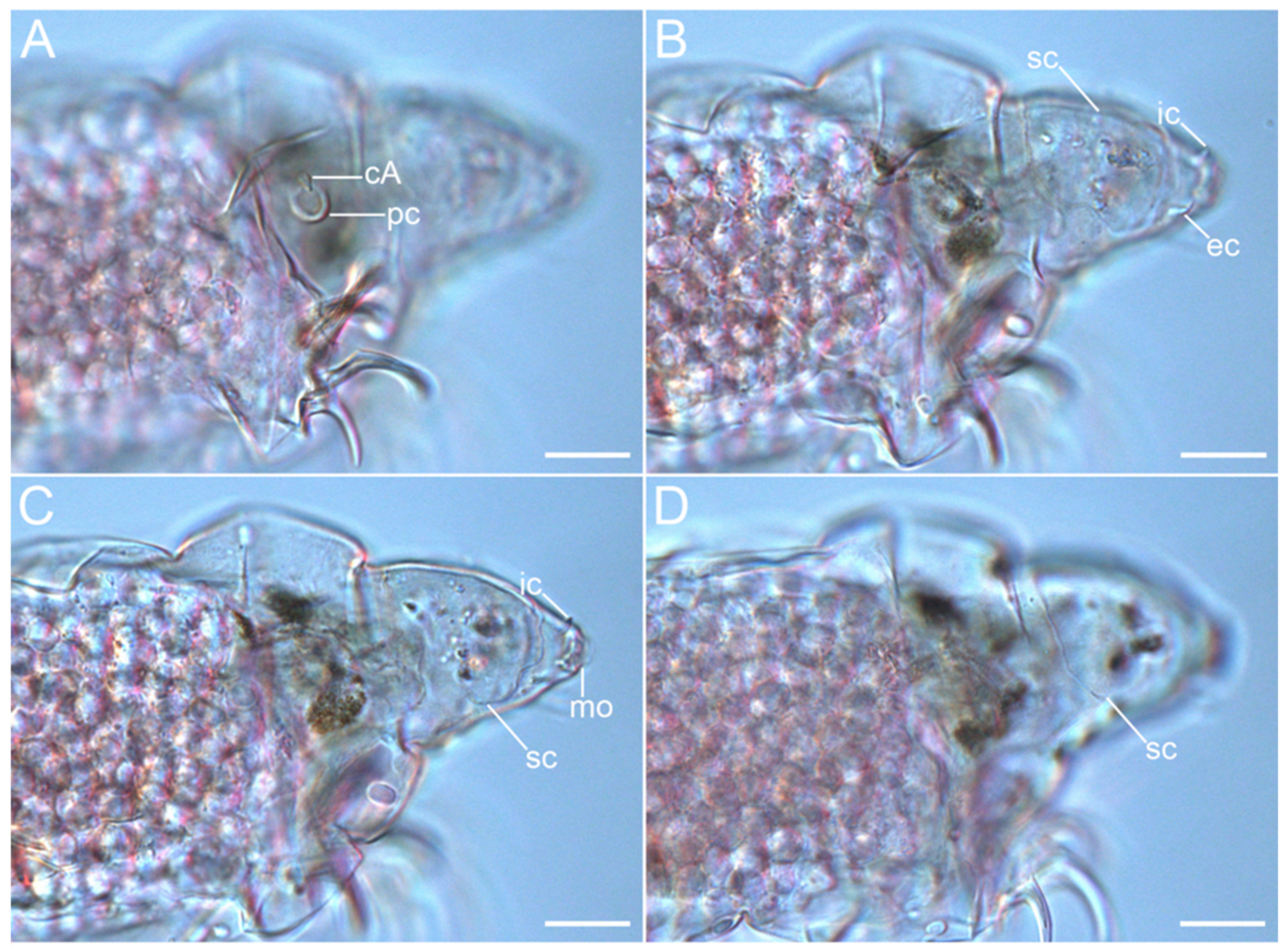
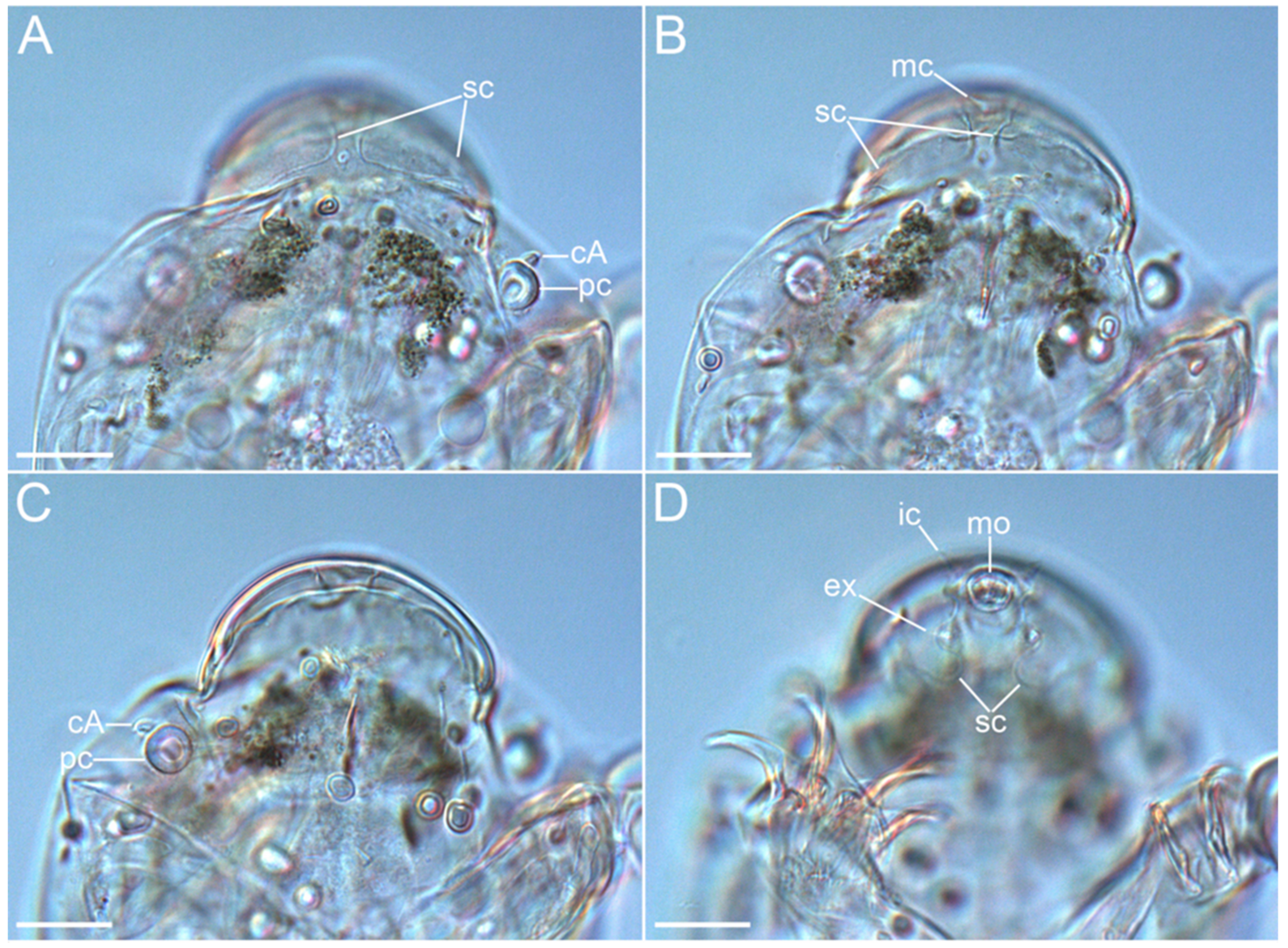
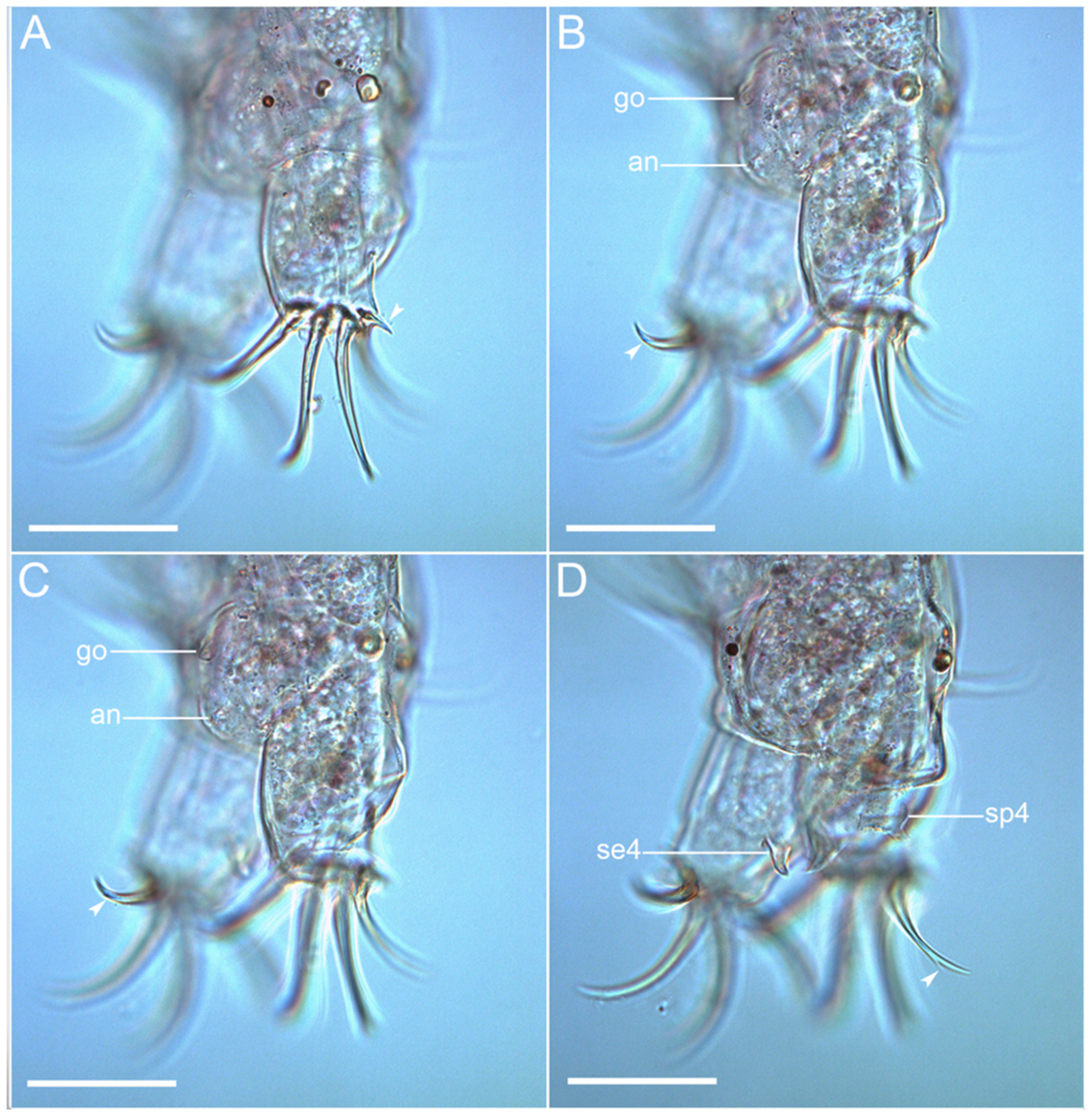
3.1. Taxonomy










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
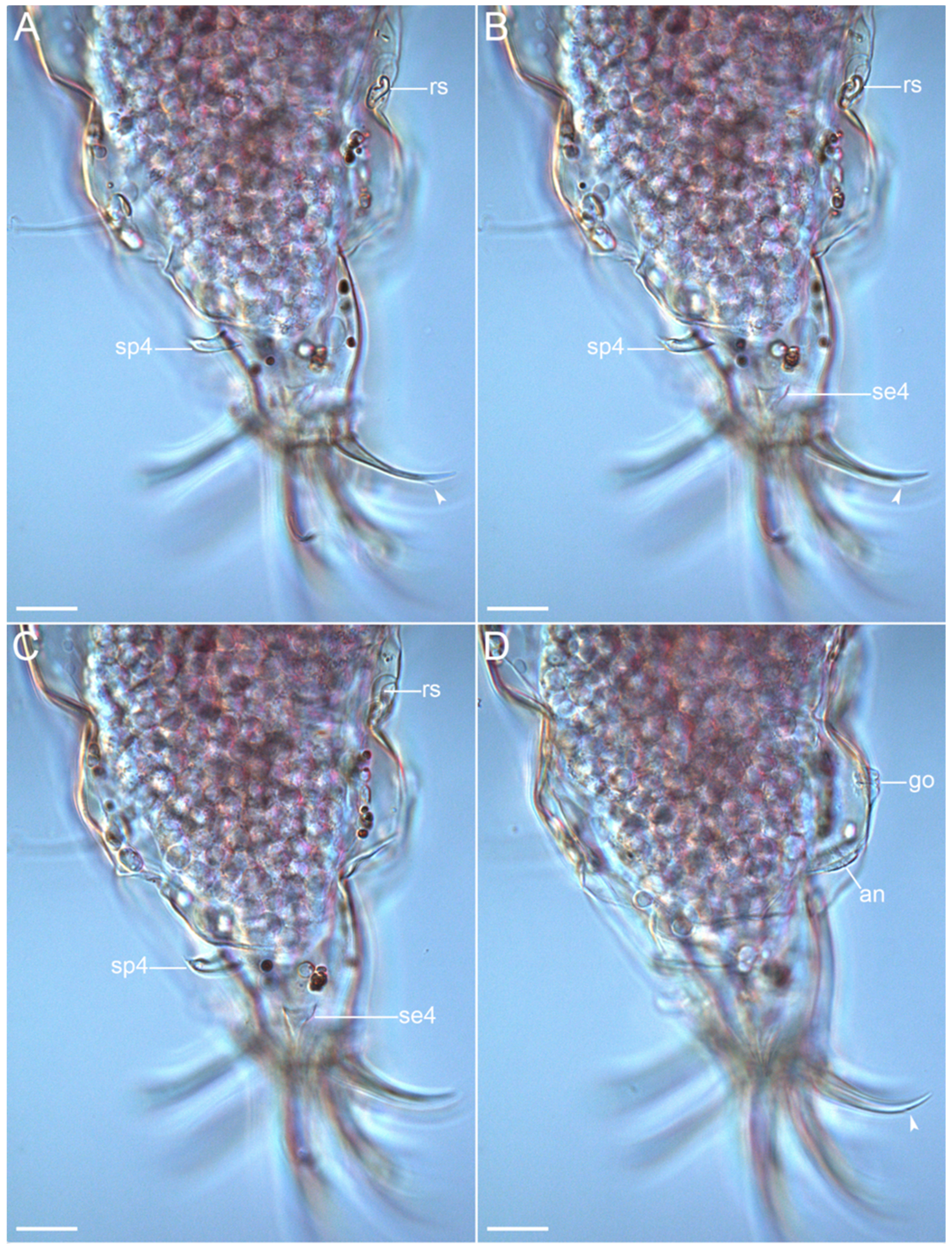{kind=link}
{kind=link}
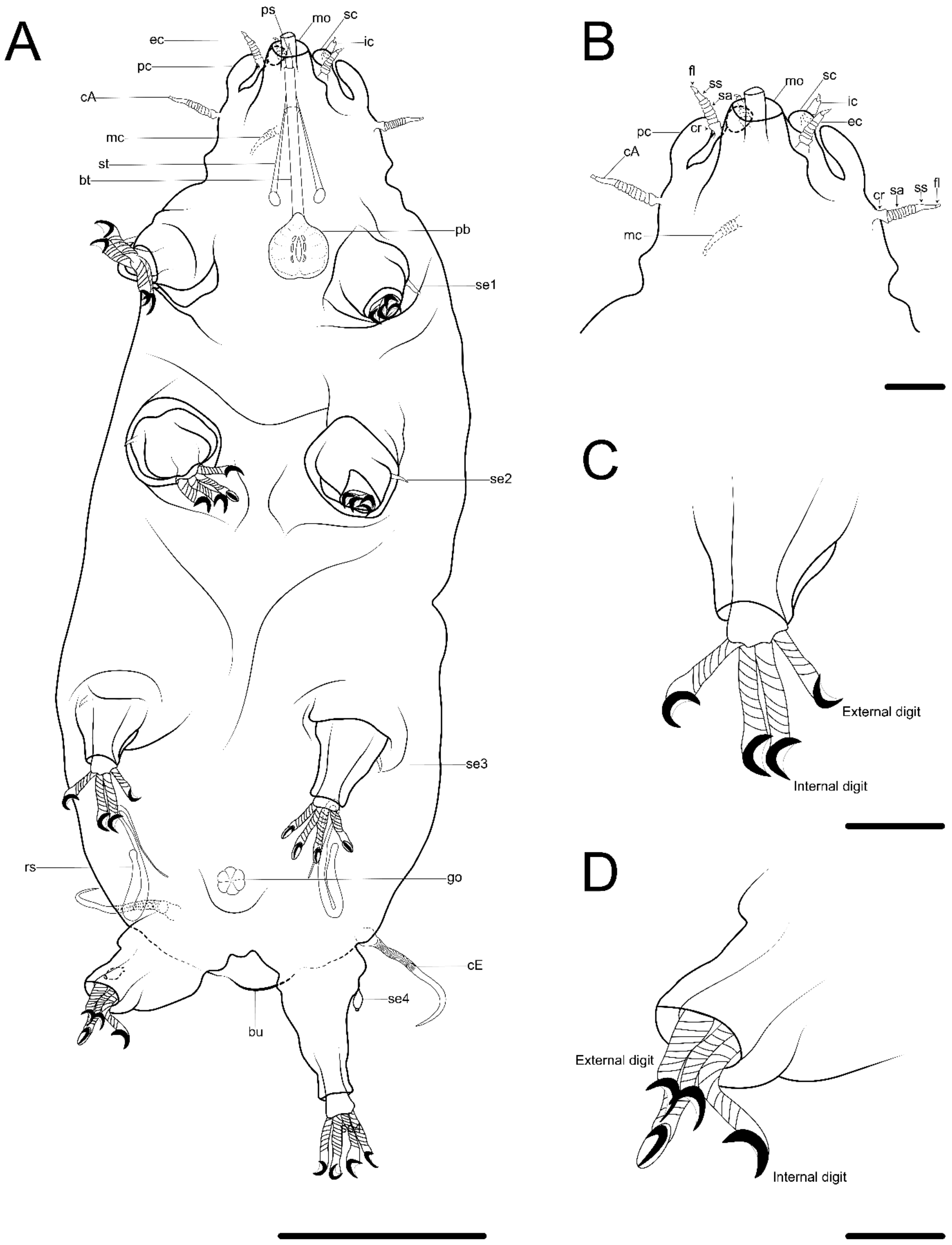{kind=link}
{kind=link}
{kind=link}
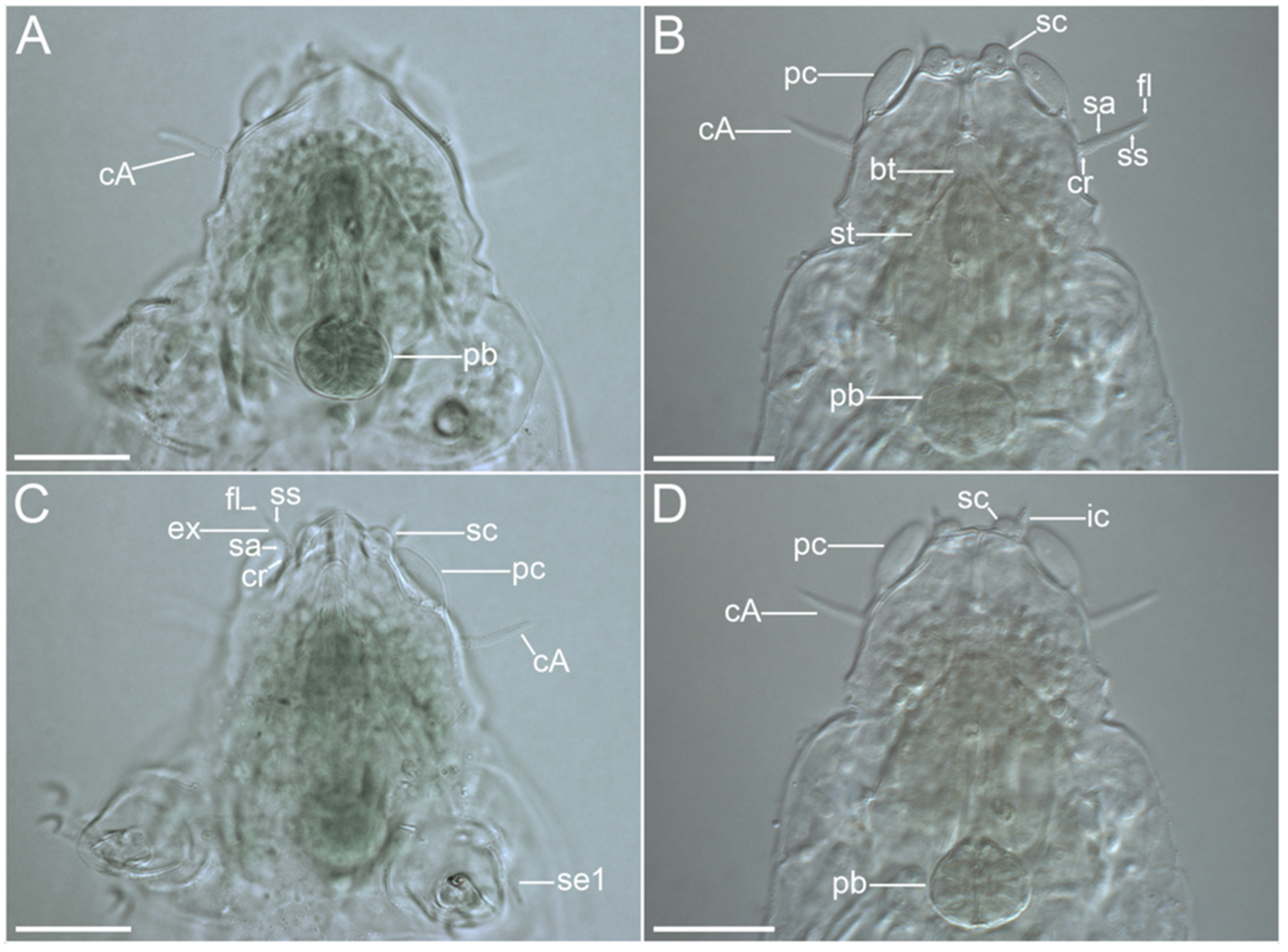{kind=link}
{kind=link}
{kind=link}
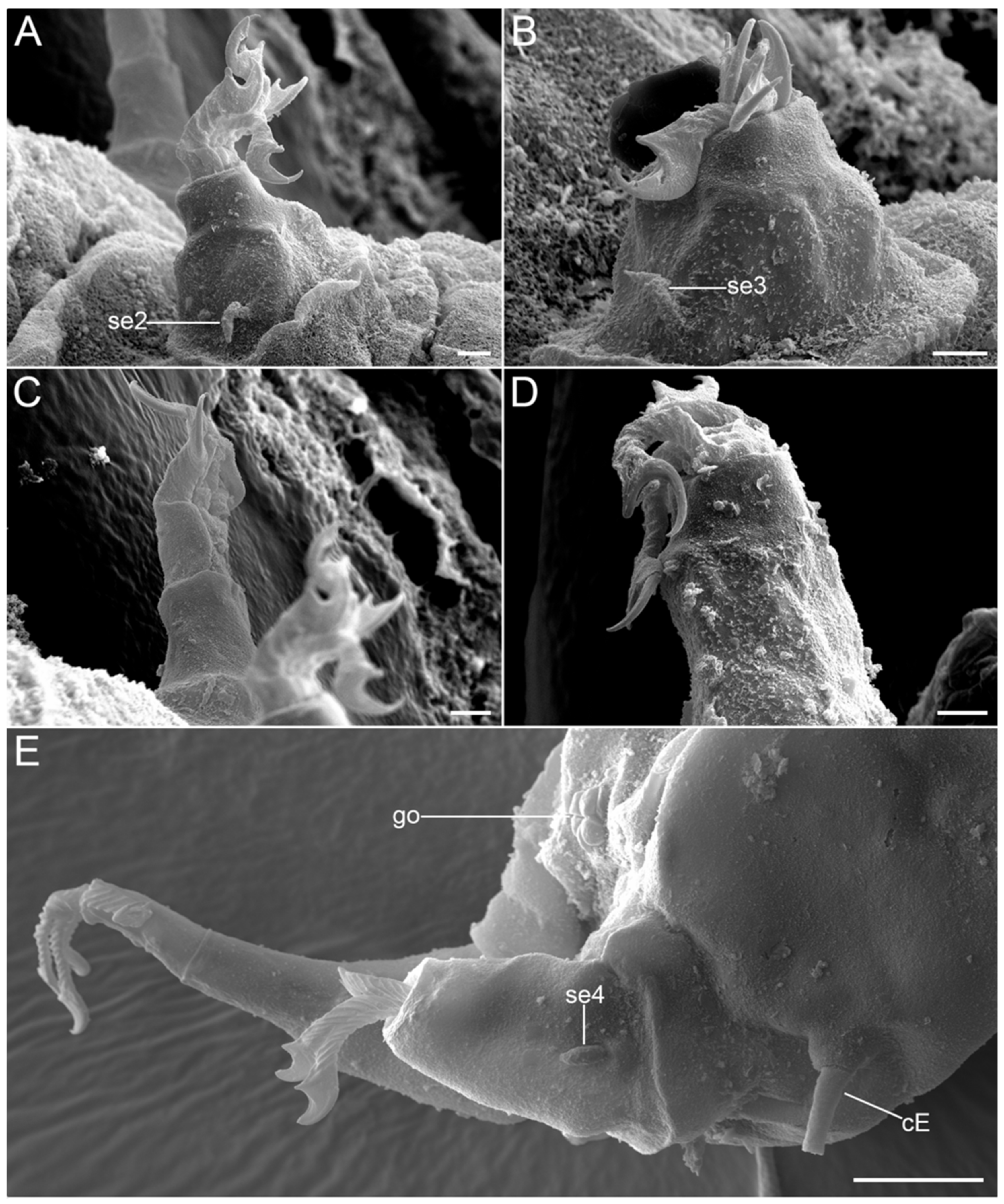{kind=link}
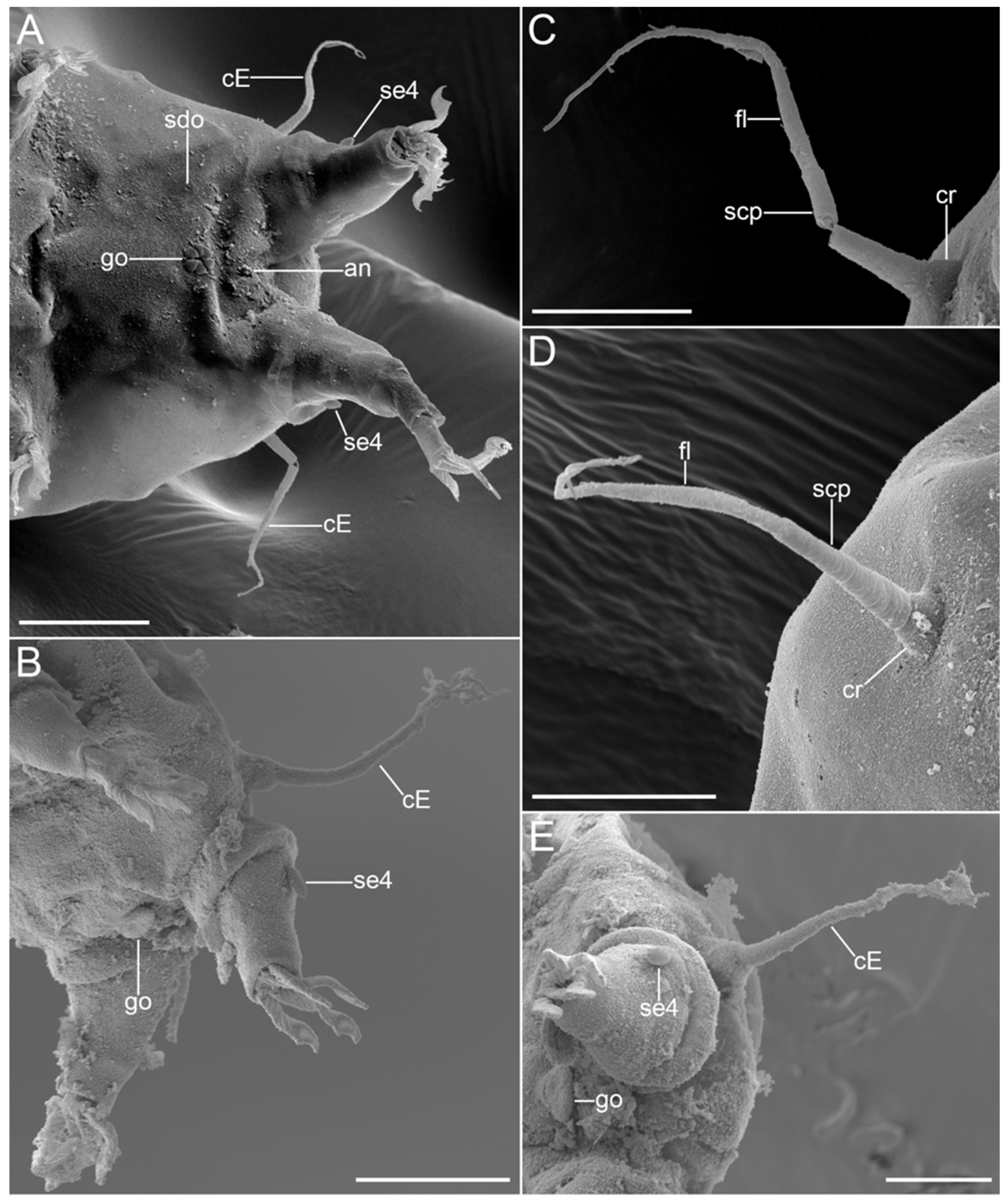{kind=link}
{kind=link}
{kind=link}
{kind=link}
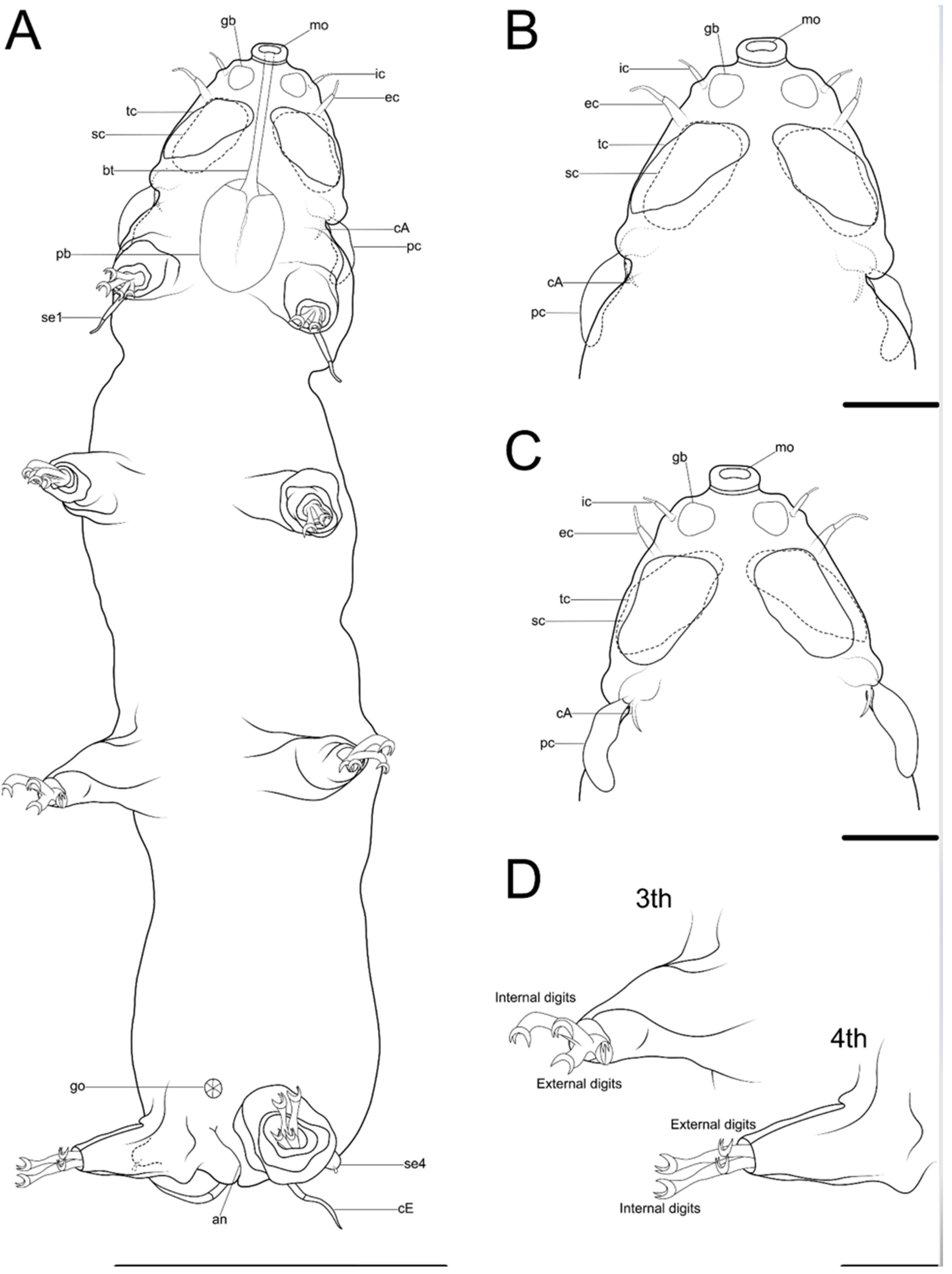{kind=link}
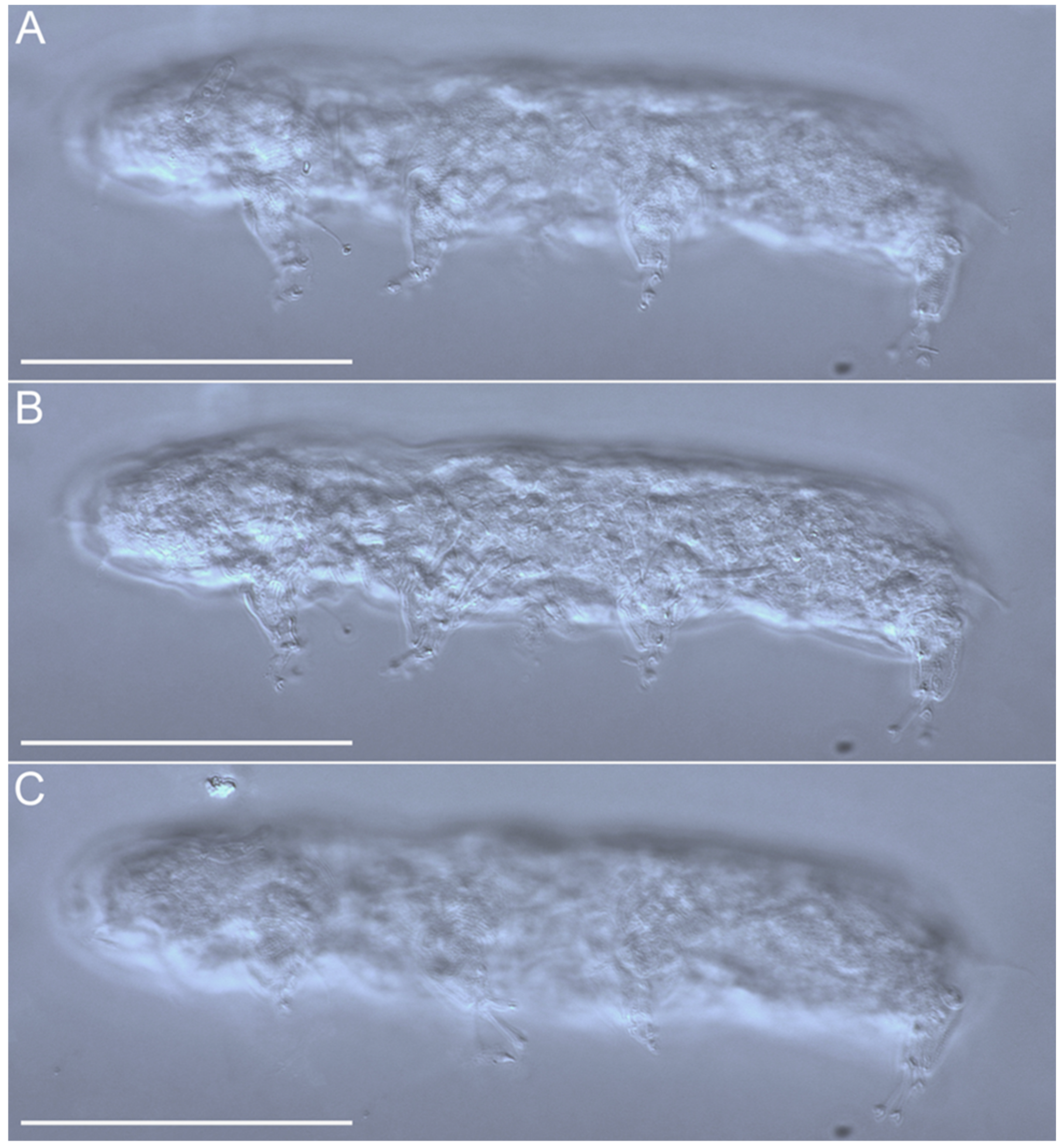{kind=link}
{kind=link}
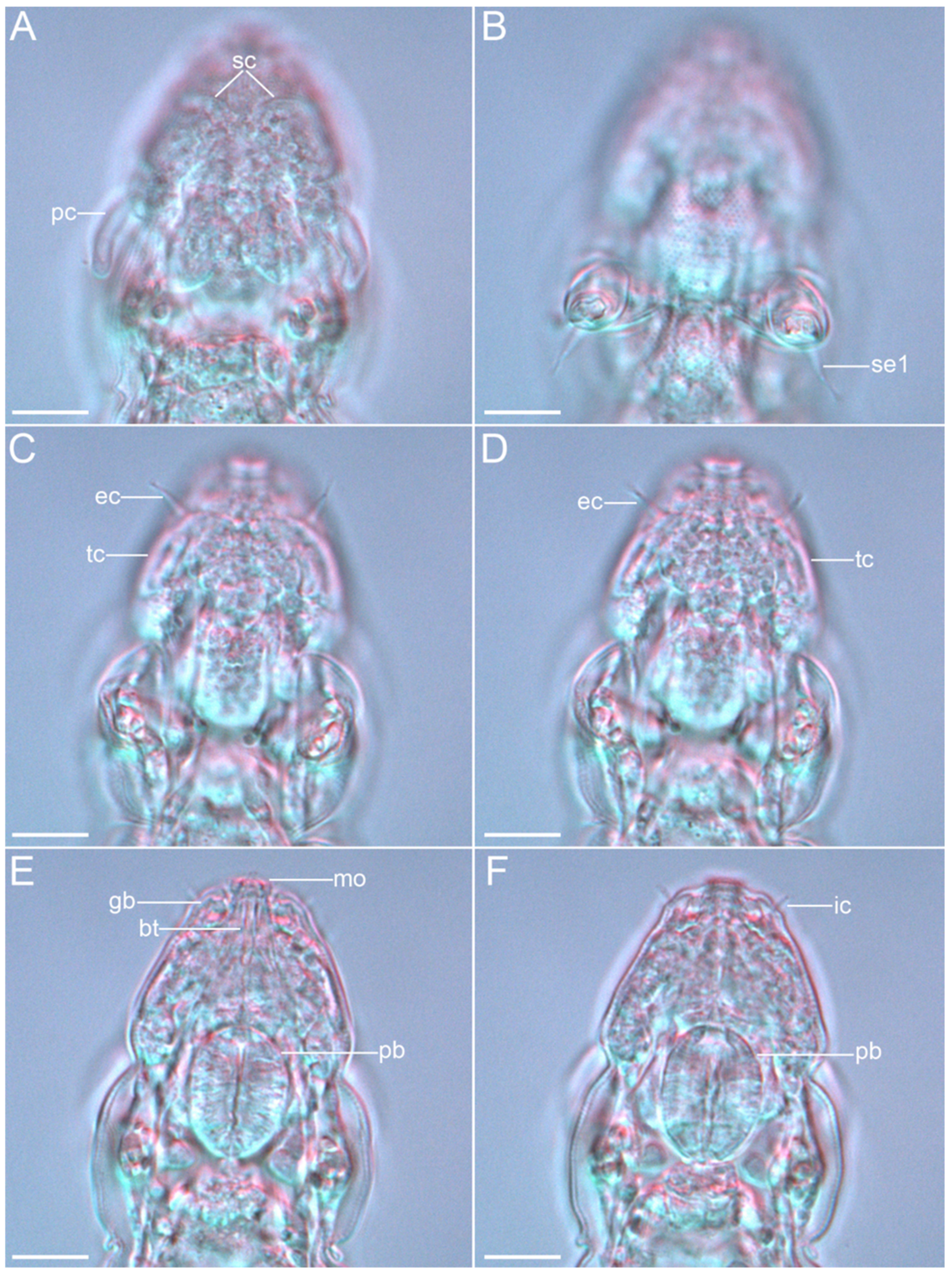{kind=link}
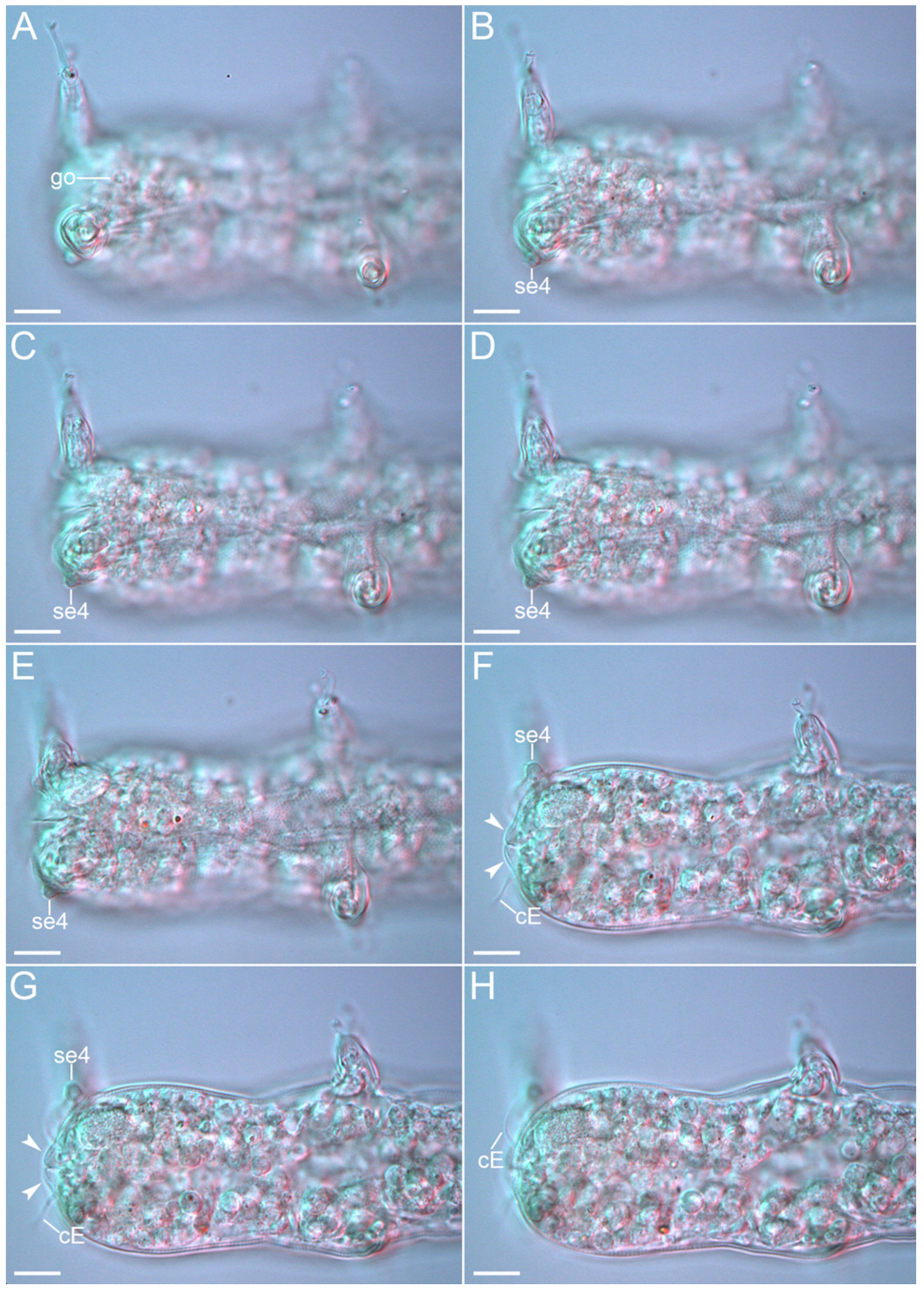{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Holotype, Male | Paratype, Male | Paratype, Female |
|---|---|---|---|
| Body length | 494 | 542 | 557 |
| Body width | 104 | 134 | 113 |
| Internal cirri | 9.6 | 8.7 | 6.9 |
| External cirri | 10.1 | 9.3 | 10.4 |
| Median cirrus | 5.3 | 5.3 | 4.2 |
| Primary clava | 8.7 | 8.4 | 9.4 |
| Cirri A | 8.6 | 6.3 | 7.3 |
| Cirri B | 7.0 | 7.0 | 7.1 |
| Cirri C | 38.0 | 21.3 | 22.7 |
| Cirri E | 36.3 | 32.7 | 30.2 |
| Sensory organ on leg I | 6.3 | ? | 9.9 |
| Sensory organ on leg II | 8.4 | 9.9 | 7.8 |
| Sensory organ on leg III | 9.1 | 9.3 | ? |
| Sensory organ on leg IV | 19.1 | 16.9 | 15.1 |
| Process on leg IV | 15.3 | 17.6 | 14.1 |
| Leg I: Claw I | 26.2 | 25.6 | 20.6 |
| Claw II | 36.8 | 33.0 | 26.3 |
| Claw III | 32.1 | 30.3 | ? |
| Claw IV | 26.4 | 24.0 | ? |
| Leg IV: Claw I | 48.3 | 44.2 | 49.2 |
| Claw II | 55.7 | 67.6 | 64.0 |
| Claw III | 56.5 | 58.0 | 68.5 |
| Claw IV | 45.1 | 49.6 | 51.3 |
| Gonopore–anus (distance) | 16.9 | 15.4 | 22.5 |
3.1.1. Diagnosis
3.1.2. Type Locality
3.1.3. Type Material
3.1.4. Type Repository
3.1.5. Etymology
3.1.6. Description of the Holotype
3.1.7. Differential Diagnosis
3.2. Taxonomy











| Character | Holotype, Female | Paratypes | |
|---|---|---|---|
| Males (n = 6) | Females (n = 4) | ||
| Body length | 230.7 | 188.4–276.7 ± 35.5 (n = 6) | 195.9–246.7 ± 21.1 (n = 4) |
| Body width | 92.4 | 75.8–95.6 ± 9.0 (n = 5) | 74.5–79.9 ± 3.8 (n = 2) |
| Head width | 48.3 | 34.9–44.1 ± 3.5 (n = 5) | ? |
| Median cirrus | 8.2 | ? | 6.3–9.9 ± 1.6 (n = 4) |
| Internal cirrus | 7.4 | 5.6–8.5 ± 1.1 (n = 5) | 5.6–7.4 ± 0.9 (n = 3) |
| External cirrus | 11.3 | 8.7–12.2 ± 1.4 (n = 6) | 8.8–11.5 ± 1.4 (n = 3) |
| Cirrus A | 14.3 | 12.1–14.0 ± 0.8 (n = 5) | 11.6–13.9 ± 1.7 (n = 2) |
| Primary clava | 12.0 | 11.3–13.9 ± 1.0 (n = 6) | 10.9–12.6 ± 0.8 (n = 4) |
| Secondary clava | 4.8 | 3.4–4.5 ± 0.5 (n = 6) | 4.0–4.6 ± 0.2 (n = 4) |
| Cirrus E | ? | 30.4–66.0 ± 15.5 (n = 6) | 29.2–41.8 ± 6.7 (n = 3) |
| Sensory organ on leg I | 5.8 | 5.1–6.8 ± 0.8 (n = 5) | 5.6–5.8 ± 0.1 (n = 2) |
| Sensory organ on leg II | 6.0 | 4.8–6.6 ± 0.8 (n = 4) | 4.5–6.1 ± 1.1 (n = 2) |
| Sensory organ on leg III | 5.1 | 4.6–5.2 ± 0.3 (n = 3) | ? |
| Sensory organ on leg IV | 4.8 | 4.2–4.9 ± 0.3 (n = 6) | 3.6–5.1 ± 0.8 (n = 3) |
| Internal digits on legs I–III | 8.9 | 9.7–12.7 ± 1.3 (n = 6) | 10.2–12.0 ± 0.9 (n = 4) |
| External digits on legs I–III | 7.1 | 7.0–9.5 ± 0.9 (n = 6) | 7.3–9.4 ± 0.9 (n = 4) |
| Internal digits legs IV | 12.7 | 11.3–19.1 ± 2.8 (n = 6) | 14.2–15.9 ± 0.9 (n = 3) |
| External digits legs IV | 10.3 | 8.1–14.4 ± 2.3 (n = 6) | 9.9–12.2 ± 1.2 (n = 3) |
| Caudodorsal bulge | 5.4 | 2.9–7.8 ± 1.9 (n = 6) | 2.5–9.1 ± 3.1 (n = 4) |
3.2.1. Diagnosis
3.2.2. Type Locality
3.2.3. Type Material
3.2.4. Type Repository
3.2.5. Etymology
3.2.6. Description of the Holotype
3.2.7. Differential Diagnosis
3.3. Taxonomy
3.3.1. Material Examined
3.3.2. Morphological Comments
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.R.; Bartels, P.J.; Guil, N. Tardigrade ecology. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 163–210. [Google Scholar] [CrossRef]
- Hengherr, S.; Schill, R.O. Environmental adaptations: Cryobiosis. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 295–310. [Google Scholar] [CrossRef]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species, (41st Edition: 12-05-2022); 2009–2022. Available online: http://www.evozoo.unimore.it/site/home/tardigrade-tools/documento1080026927.html (accessed on 15 September 2022).
- Mcinnes, S.J.; Jørgensen, A.; Michalczyk, Ł. 20 Years of Zootaxa: Tardigrada (Ecdysozoa: Panarthropoda). Zootaxa 2021, 4979, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, L.; Bartels, P.J.; Roszkowska, M.; Nelson, D.R. The zoogeography of marine Tardigrada. Zootaxa 2015, 4037, 1–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.; Sattarova, V.V.; Katrynski, L.; Martínez Arbizu, P. New insights from the deep: Meiofauna in the Kuril-Kamchatka Trench and adjacent abyssal plain. Prog. Oceanogr. 2019, 173, 192–207. [Google Scholar] [CrossRef]
- Fontoura, P.; Bartels, P.J.; Jørgensen, A.; Kristensen, R.M.; Hansen, J.G. A dichotomous key to the genera of the marine heterotardigrades (Tardigrada). Zootaxa 2017, 4294, 1–45. [Google Scholar] [CrossRef]
- Topstad, L.; Guidetti, R.; Majaneva, M.; Ekrem, T. Multi-marker DNA metabarcoding reflects tardigrade diversity in different habitats. Genome 2021, 64, 217–231. [Google Scholar] [CrossRef]
- Bartels, P.J.; Apodaca, J.J.; Mora, C.; Nelson, D.R. A global biodiversity estimate of a poorly known taxon: Phylum Tardigrada. Zool. J. Linn. Soc. 2016, 178, 730–736. [Google Scholar] [CrossRef] [Green Version]
- Tchesunov, A.V. A new tardigrade species of the genus Neostygarctus Grimaldi de Zio et al., 1982 (Tardigrada, Arthrotardigrada) from the Great Meteor Seamount, Northeast Atlantic. Eur. J. Taxon. 2018, 479, 1–17. [Google Scholar] [CrossRef]
- Bai, L.; Wang, X.; Zhou, Y.; Lin, S.; Meng, F.; Fontoura, P. Moebjergarctus clarionclippertonensis, a new abyssal tardigrade (Arthrotardigrada, Halechiniscidae, Euclavarctinae) from the Clarion-Clipperton Fracture Zone, North-East Pacific. Zootaxa 2020, 4755, 561–575. [Google Scholar] [CrossRef]
- Gomes-Júnior, E.; Santos, É.; da Rocha, C.M.C.; Santos, P.J.P.; Fontoura, P. The deep-sea genus Coronarctus (Tardigrada, Arthrotardigrada) in Brazil, south-western Atlantic Ocean, with the description of three new species. Diversity 2020, 12, 63. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.G.; Kristensen, R.M. A new genus and five new species of the subfamily Florarctinae (Tardigrada, Arthrotardigrada). Eur. J. Taxon. 2021, 762, 149–184. [Google Scholar] [CrossRef]
- Fujimoto, S.; Hansen, J.G. Revision of Angursa (Arthrotardigrada: Styraconyxidae) with the description of a new species from Japan. Eur. J. Taxon. 2019, 2019, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, C.; Martínez Arbizu, P. Unexpectedly higher metazoan meiofauna abundances in the Kuril–Kamchatka Trench compared to the adjacent abyssal plains. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 111, 60–75. [Google Scholar] [CrossRef]
- Brandt, A.; Brix, S.; Riehl, T.; Malyutina, M. Biodiversity and biogeography of the abyssal and hadal Kuril-Kamchatka trench and adjacent NW Pacific deep-sea regions. Prog. Oceanogr. 2020, 181, 102232. [Google Scholar] [CrossRef]
- McIntyre, A.D.; Warwick, R.M. Meiofauna techniques. In Methods for the Study of Marine Benthos; Blackwell: Oxford, UK, 1984. [Google Scholar]
- Guil, N.; Jørgensen, A.; Kristensen, R. An upgraded comprehensive multilocus phylogeny of the Tardigrada tree of life. Zool. Scr. 2019, 48, 120–137. [Google Scholar] [CrossRef]
- Møbjerg, N.; Jørgensen, A.; Kristensen, R.M.; Cardoso Neves, R.A. Morphology and functional anatomy. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 57–94. [Google Scholar] [CrossRef]
- Perry, E.; Miller, W.R.; Kaczmarek, Ł. Recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2019, 4608, 145–154. [Google Scholar] [CrossRef]
- Renaud-Mornant, J. Bathyal and abyssal Coronarctidae (Tardigrada), description of new species and phylogenetical significance. In Biology of Tardigrades. Selected Symposia and Monographs U.Z.I; Bertolani, R., Ed.; Mucchi Editore: Modena, Italy, 1987; Volume 1, pp. 93–101. [Google Scholar]
- Romano, F., III; Gallo, M.; D’Addabbo, R.; Accogli, G.; Bagueley, J.; Montagna, P. Deep-sea tardigrades in the northern Gulf of Mexico with a description of a new species of Coronarctidae (Tardigrada: Arthrotardigrada), Coronarctus mexicus. J. Zool. Syst. Evol. Res. 2011, 49, 48–52. [Google Scholar] [CrossRef]
- Bussau, C. New deep-sea Tardigrada (Arthrotardigrada, Halechiniscidae) from a manganese nodule area of the Eastern South Pacific. Zool. Scr. 1992, 21, 79–92. [Google Scholar] [CrossRef]
- Pollock, L.W. Angursa bicuspis n. g., n. sp., a new marine arthrotardigrade from the western North Atlantic. Trans. Am. Microsc. Soc. 1979, 98, 558–562. [Google Scholar] [CrossRef]
- Noda, H. Description of a new subspecies of Angursa biscupis Pollock (Heterotardigrada, Halechiniscidae) from Tanabe Bay, Japan. Publ. Seto Mar. Biol. Lab. 1985, 30, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Renaud-Mornant, J. Tardigrada. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washigton, DC, USA, 1988; pp. 357–361. [Google Scholar]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer Verlag: Berlin/Heidelberg, Germany, 2009; p. 527. [Google Scholar] [CrossRef]
- Hansen, J.G.; Jørgensen, A.; Kristensen, R.M. Preliminary studies of the tardigrade fauna of the Faroe Bank. Zool. Anzeiger A J. Comp. Zool. 2001, 240, 385–393. [Google Scholar] [CrossRef]
- Hansen, J.G. The ongoing investigation of the Faroe Bank tardigrade fauna. In Proceedings of the BIOFAR Symposium, Tórshavn, Faroe Islands, 24–26 April 2003, North-East Atlantic Marine Benthic Oorganisms in the Faroes—Taxonomy, Distribution and Ecology, Tórshavn, Faroe Islands; Frøᵭskapparit Supplementum; Føroya Frỏðskaparfelag: Tórshavn, Faroe Islands, 2005; pp. 220–223. [Google Scholar]
- Jörgensen, A.; Kristensen, R.M. A New tanarctid Arthrotardigrade with buoyant bodies. Zool. Anzeiger A J. Comp. Zool. 2001, 240, 425–439. [Google Scholar] [CrossRef]
- Hansen, J.G.; D’Addabbo, M.G.; de Zio Grimaldi, S. A Comparison of morphological characters within the genus Rhomboarctus (Tardigrada: Heterotardigrada) with the description of two new species. Zool. Anzeiger A J. Comp. Zool. 2003, 242, 83–96. [Google Scholar] [CrossRef]
- Kristensen, R.M.; Sørensen, M.V.; Hansen, J.G.; Zeppilli, D. A New species of Neostygarctus (Arthrotardigrada) from the Condor Seamount in the Azores, Northeast Atlantic. Mar. Biodivers. 2015, 45, 453–467. [Google Scholar] [CrossRef]
- Pérez-Pech, W.A.; Hansen, J.G.; Demilio, E.; de Jesús-Navarrete, A.; Mendoza, I.M.; Olivares, A.R.; Vargas-Espositos, A. First records of marine tardigrades of the genus Coronarctus (Tardigrada, heterotardigrada, arthrotardigrada) from Mexico. Check List 2019, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bartels, P.J.; Fontoura, P.; Nelson, D.R. Marine tardigrades of the Bahamas with the description of two new species and updated keys to the species of Anisonyches and Archechiniscus. Zootaxa 2018, 4420, 43–70. [Google Scholar] [CrossRef]
- Renaud-Mornant, J. Deep-sea tardigrada from the “Meteor” Indian Ocean Expedition. Meteor Forsch. Reihe D Biol. 1975, 21, 54–61. [Google Scholar]
- Renaud-Mornant, J. Tardigrades abyssaux nouveaux de la sous-famille des Euclavarctinae n. subfam (Arthrotardigrada, Halechiniscidae). Bulletin du Muséum National d’Histoire Naturelle, Section A. Zoologie Biologie et Ecologie Animales 1983, 51, 201–219. [Google Scholar]
- Steiner, G. Bathyechiniscus tetronyx n. g., n. sp., Ein neuer mariner Tardigrade. Deutsche Sudpolar Expedition, 1901–1903. Zoology 1926, 10, 479–481. [Google Scholar]
- Renaud-Mornant, J. Sous-famille et genre nouveaux de Tardigrades marins (Arthrotardigrada). Bull. Du Muséum Natl. D’histoire Nat. 1982, 4, 89–94. [Google Scholar]
- Renaud-Mornant, J. Halechiniscidae (Heterotardigrada) de la campagne Benthedi, canal du Mozambique. Bull. Du Muséum Natl. D’histoire Nat. 1984, 6, 67–88. [Google Scholar]
- Bartels, P.J.; Fontaneto, D.; Roszkowska, M.; Nelson, D.R.; Kaczmarek, Ł. Latitudinal gradients in body size in marine tardigrades. Zool. J. Linn. Soc. 2020, 188, 820–838. [Google Scholar] [CrossRef]
- Kristensen, R.M.; Mackness, B.S. First record of the marine tardigrade genus Batillipes (Arthrotardigrada: Batillipedidae) from South Australia with a description of a new species. Rec. South Aust. Mus. 2000, 33, 73–87. [Google Scholar]
- Renaud-Mornant, J. Deux nouveaux Angursa Pollock, 1979, du domaine abyssal (Tardigrada, Arthrotardigrada). Tethys 1981, 10, 161–164. [Google Scholar]
- Villora-Moreno, S. Deep-sea Tardigrada from South Shetland Islands (Antarctica) with description of Angursa antarctica sp. nov. (Arthrotardigrada, Halechiniscidae). Polar Biol. 1998, 19, 336–341. [Google Scholar] [CrossRef]









| Cruise (Year) | Station | Latitude, N | Longitude, E | Depth, m | Mean Abundance (ind./10 cm2) | References | Species |
|---|---|---|---|---|---|---|---|
| SoJaBio 2010 | A3 | 44.80 | 137.25 | 1473 | 0.03 | This study | Angursa sp. |
| SokhoBio 2015 | SB4 | 47.20 48.62 | 149.62 150.00 | 3366 | 0.08 | This study | Moebjergarctus okhotensis sp. nov. |
| This study | Angursa cf. bicuspis | ||||||
| SokhoBio 2015 | SB5 | 48.05 | 150.00 | 1700 | 0.3 | This study | Angursa cf. bicuspis |
| SokhoBio 2015 | SB6 | 46.95 46.27 | 151.08 152.03 | 3351 | 1.23 | This study | Moebjergarctus okhotensis sp. nov. |
| This study | Angursa cf. bicuspis | ||||||
| SokhoBio 2015 | SB7 | 45.60 | 146.38 | 3300 | 1.31 | This study | Angursa cf. bicuspis |
| SokhoBio 2015 | SB9 | 43.97 | 157.33 | 3342 | 0.04 | This study | Angursa cf. bicuspis |
| SokhoBio 2015 | SB11 | 46.23 | 155.55 | 3206 | 1.52 | This study | Angursa cf. bicuspis |
| KuramBio I (2012) | KBI-1 | 46.97 | 154.54 | 5410 | 1.16 | Schmidt and Martinez Arbizu, 2015 [15] | Coronarctus sonne sp. nov. |
| KuramBio I (2012) | KBI-2 | 43.58 | 153.97 | 4869 | 0.02 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio I (2012) | KBI-4 | 42.48 | 153.99 | 4972 | 0.14 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio I (2012) | KBI-5 | 43.04 | 152.99 | 5378 | 1.27 | Schmidt and Martinez Arbizu, 2015 [15] | Coronarctus sonne sp. nov. |
| KuramBio I (2012) | KBI-6 | 42.24 | 151.73 | 5298 | 0.05 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio I (2012) | KBI-7 | 40.58 | 151.00 | 5221 | 0.02 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio I (2012) | KBI-8 | 43.82 | 151.76 | 5128 | 0.02 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio I (2012) | KBI-9 | 45.17 | 153.76 | 5406 | 0.02 | Schmidt and Martinez Arbizu, 2015 [15] | ? |
| KuramBio II (2016) | KBII-4 | 44.66 | 151.47 | 5143 | 0.28 | Schmidt et al., 2019 [6] | ? |
| KuramBio II (2016) | KBII-62 | 45.02 | 151.05 | 5743 | 0.28 | Schmidt et al., 2019 [6] | Angursa sp. |
| KuramBio II (2016) | KBII-74 | 44.11 | 150.60 | 8226 | 0.05 | Schmidt et al., 2019 [6] | ? |
| KuramBio II (2016) | KBII-83 | 44.21 | 150.60 | 5212 | 0.05 | Schmidt et al., 2019 [6] | Angursa sp. |
| KuramBio II (2016) | KBII-96 | 44.80 | 137.25 | 6515 | 0.42 | Schmidt et al., 2019 [6] | ? |
| KuramBio II (2016) | KBII-101 | 47.20 | 149.62 | 9540 | 0.24 | Schmidt et al., 2019 [6] | ? |
| Character | Specimen 1, Female | Specimen 2, Female | Specimen 3, Male | Specimen 4, Male | Specimen 5, Male | Specimen 6, Male |
|---|---|---|---|---|---|---|
| Body length | 159.1 | 177.7 | 147.8 | 137.7 | 145.9 | 179.3 |
| Body width | 38.4 | ? | 42.7 | 43.1 | 40.3 | ? |
| Head width | 29.4 | ? | 33.7 | 35.5 | 33.3 | ? |
| Internal cirri | 3.2 | 2.4 | ? | 4.4 | ? | ? |
| External cirri | 5.8 | 8.0 | 7.0 | 6.4 | 7.4 | 5.1 |
| Median cirrus | ? | ? | ? | ? | ||
| Cirri A | 4.3 | 3.5 | ? | 7.7 | 7.9 | 4.5 |
| Primary clava | 10.3 | 12.3 | 11.1 | 10.5 | 9.8 | 12.8 |
| Secondary clava | 11.4 | 10.6 | ? | ? | ? | ? |
| Tertiary clava | 10.3 | 10.6 | ? | ? | ? | ? |
| Distance between secondary clavae | 2.8 | ? | ? | ? | ? | ? |
| Cirri E | 12.8 | 12.4 | 14.1 | 13.8 | ? | 14.7 |
| Sensory organ leg I | 12.1 | 12.2 | ? | 7.6 | ? | 12.1 |
| Sensory organ leg II | ? | ? | ? | ? | ? | ? |
| Sensory organ leg III | ? | ? | ? | ? | ? | ? |
| Papillae of legs IV | 3.2 | 4.0 | 4.3 | 4.2 | ? | 3.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saulenko, A.A.; Maiorova, A.S.; Martínez Arbizu, P.; Mordukhovich, V.V. Deep-Sea Tardigrades from the North-Western Pacific, with Descriptions of Two New Species. Diversity 2022, 14, 1086. https://doi.org/10.3390/d14121086
Saulenko AA, Maiorova AS, Martínez Arbizu P, Mordukhovich VV. Deep-Sea Tardigrades from the North-Western Pacific, with Descriptions of Two New Species. Diversity. 2022; 14(12):1086. https://doi.org/10.3390/d14121086
Chicago/Turabian StyleSaulenko, Anastasiia A., Anastassya S. Maiorova, Pedro Martínez Arbizu, and Vladimir V. Mordukhovich. 2022. "Deep-Sea Tardigrades from the North-Western Pacific, with Descriptions of Two New Species" Diversity 14, no. 12: 1086. https://doi.org/10.3390/d14121086
APA StyleSaulenko, A. A., Maiorova, A. S., Martínez Arbizu, P., & Mordukhovich, V. V. (2022). Deep-Sea Tardigrades from the North-Western Pacific, with Descriptions of Two New Species. Diversity, 14(12), 1086. https://doi.org/10.3390/d14121086