
Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Samples
2.2. DNA Extraction and Sequencing
2.3. Assembly and Annotation
2.4. Sequence Analysis
2.5. Phylogenetic Analysis
3. Results
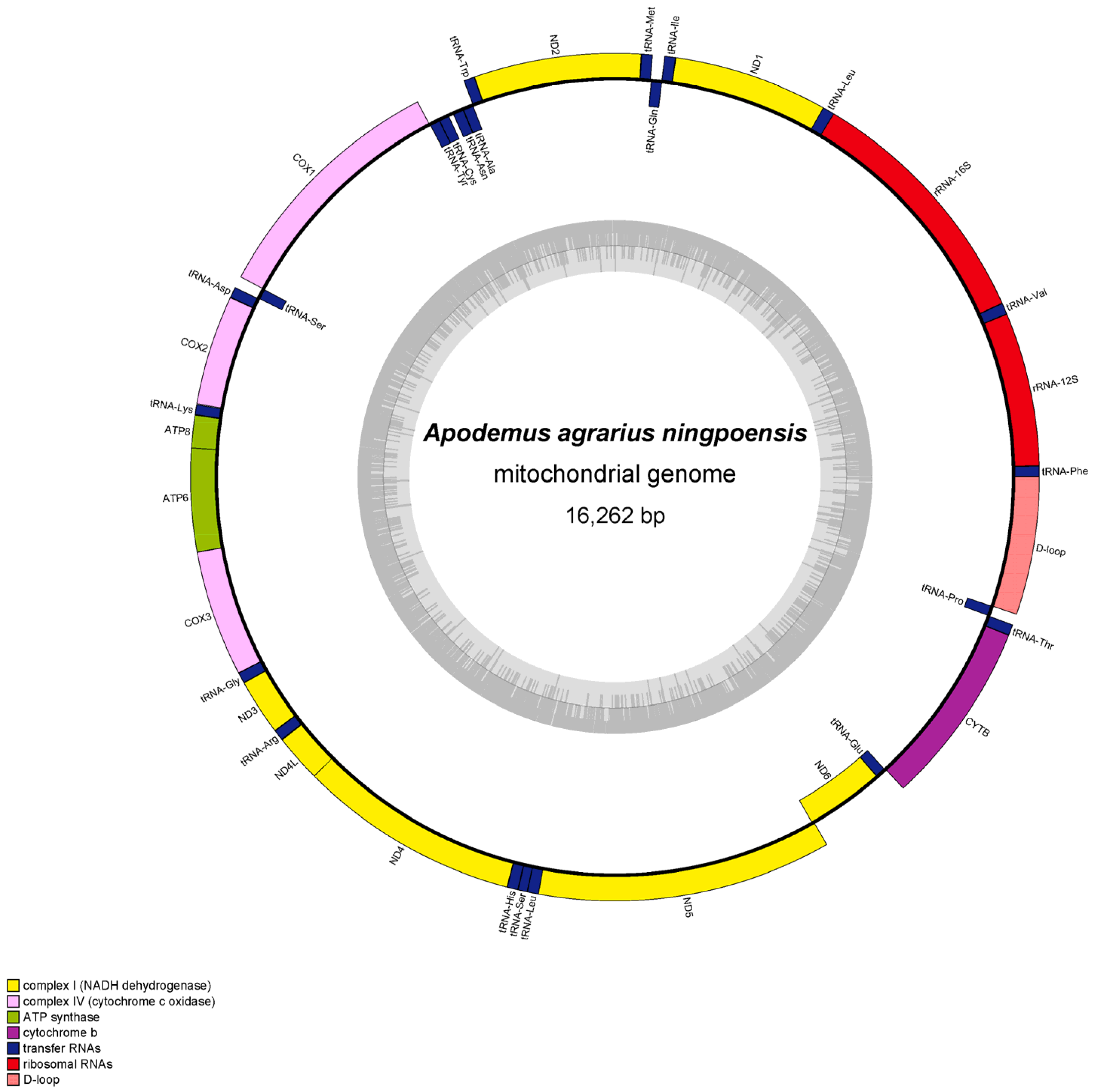
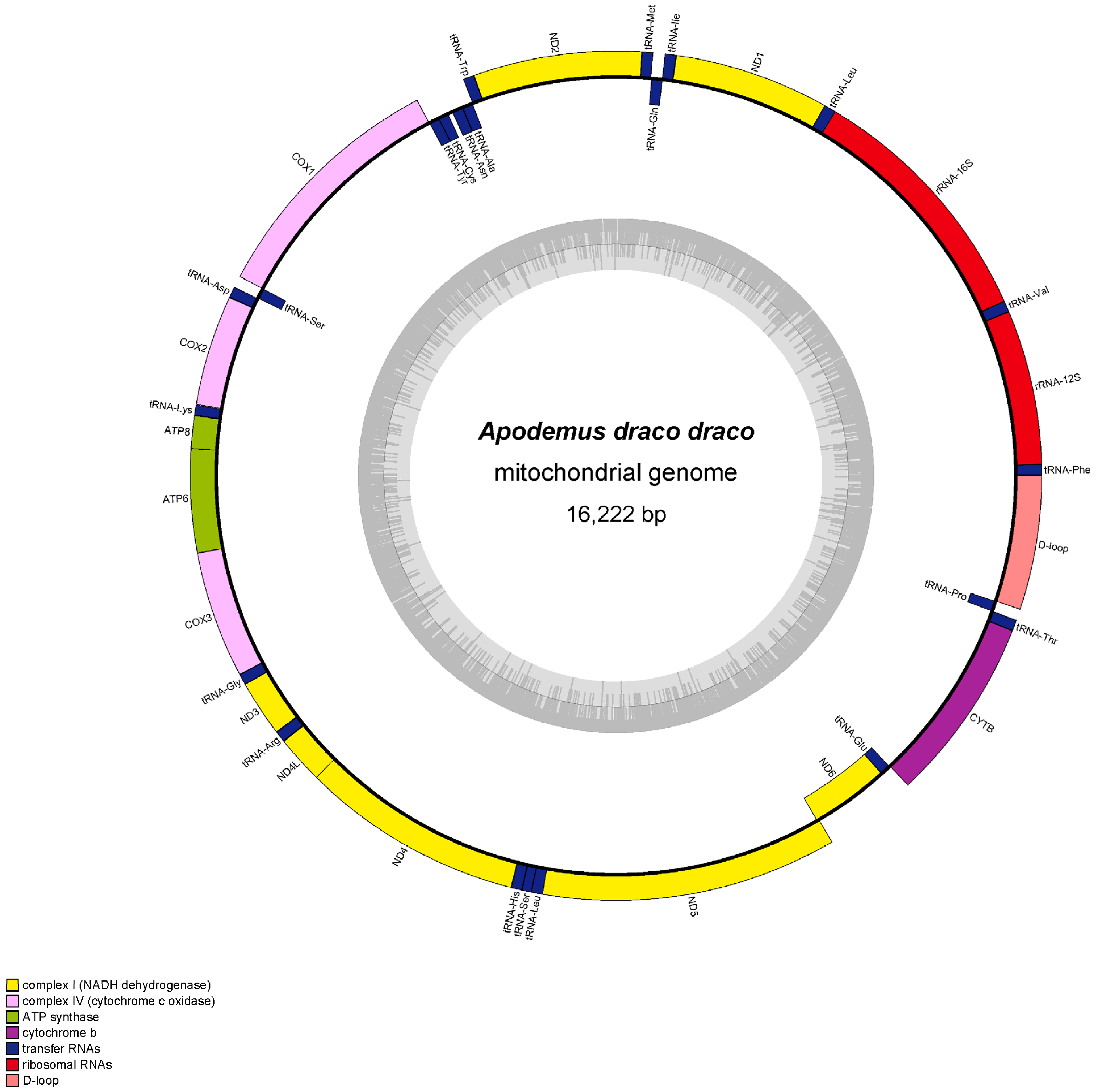
3.1. Features of Mitogenomes
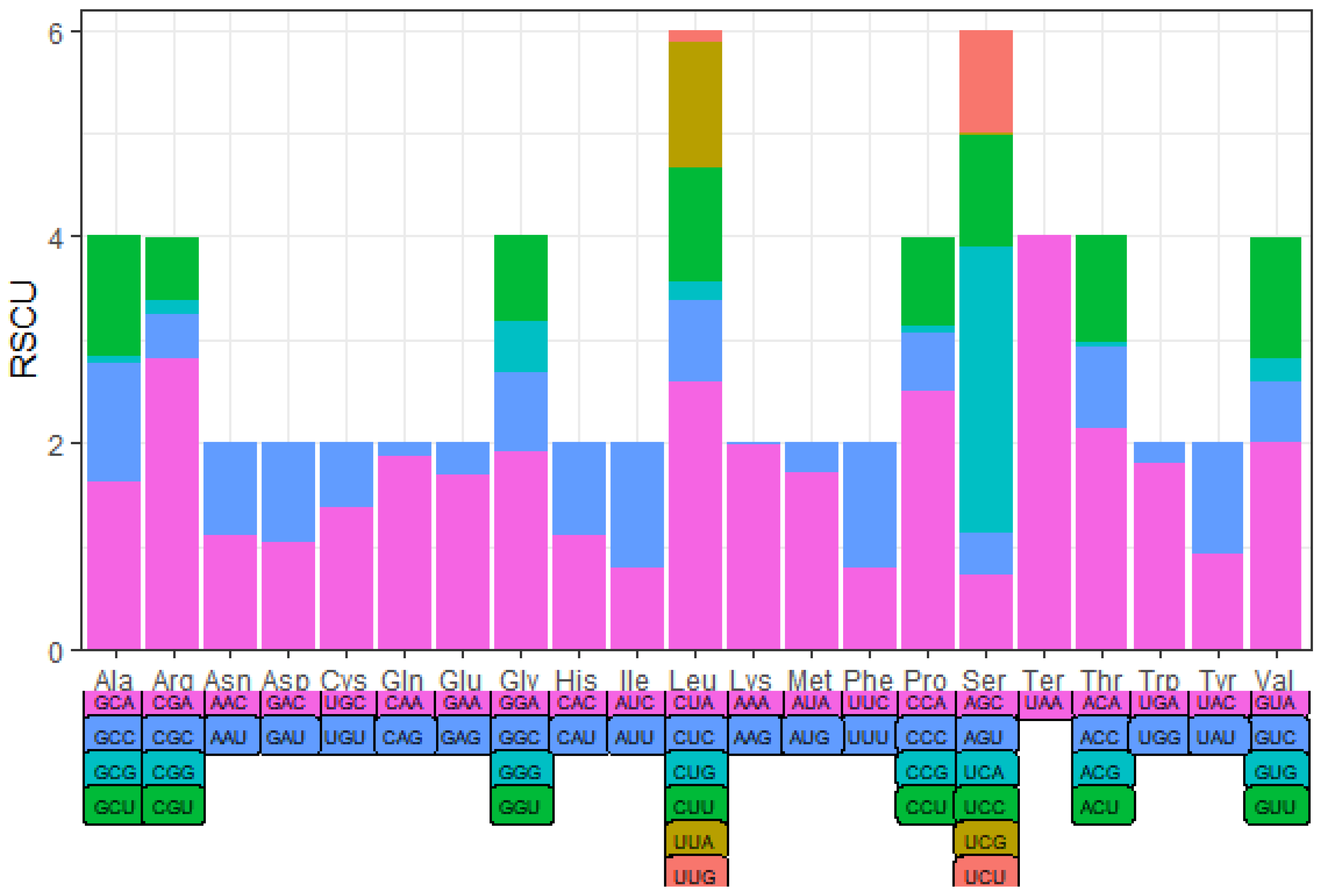
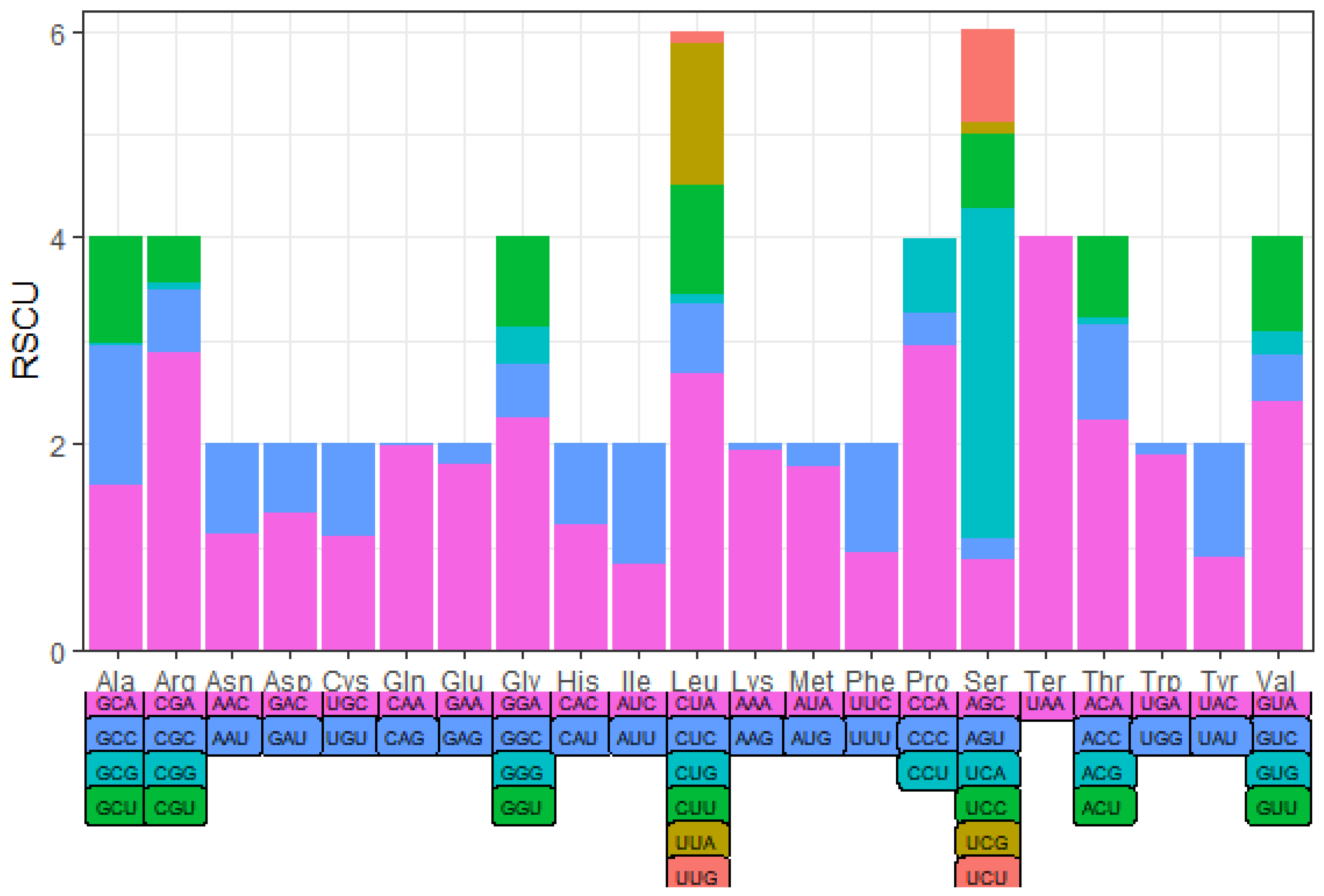
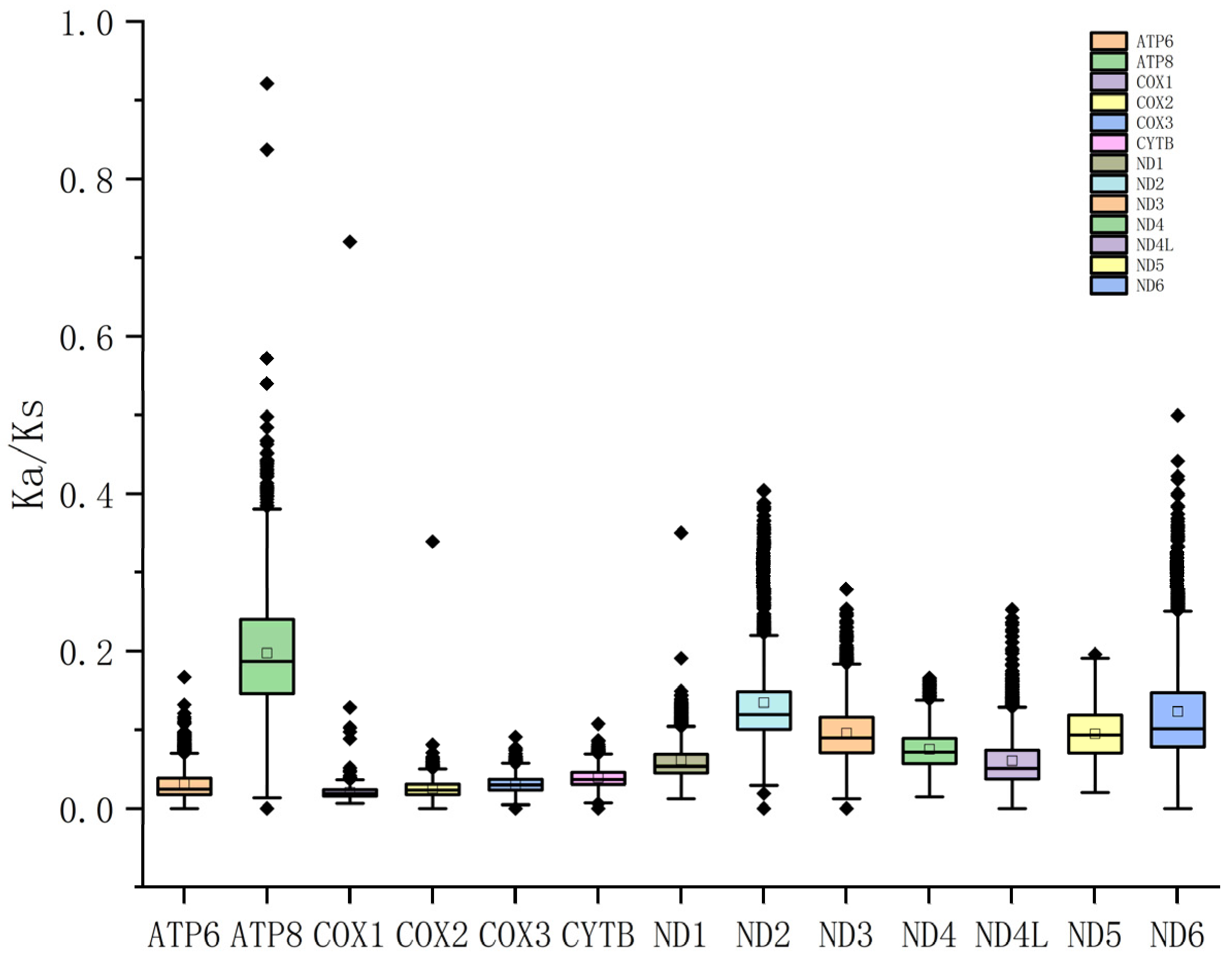
3.2. Protein-Coding Genes and Codon Usage
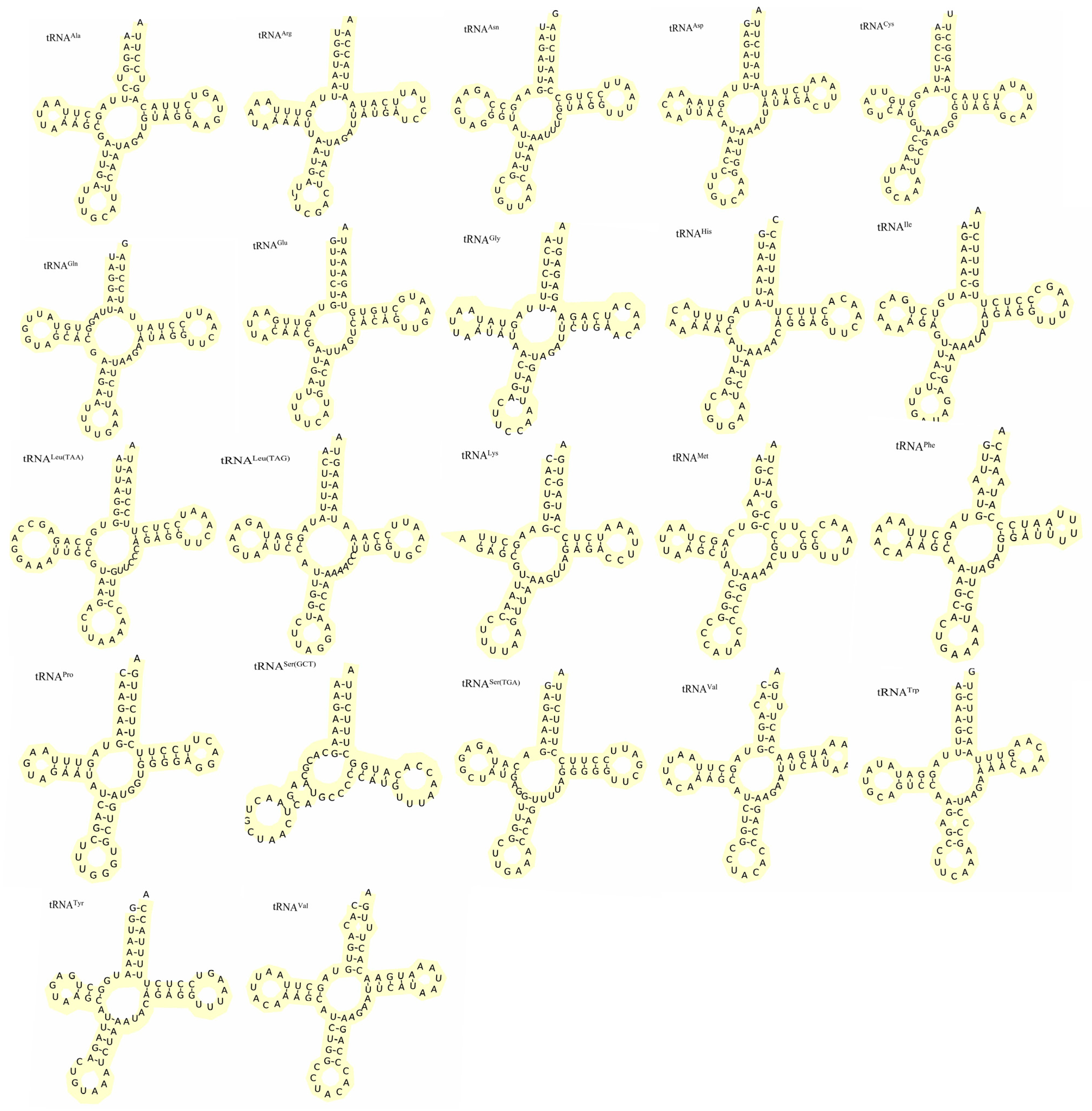
3.3. tRNAs and rRNAs
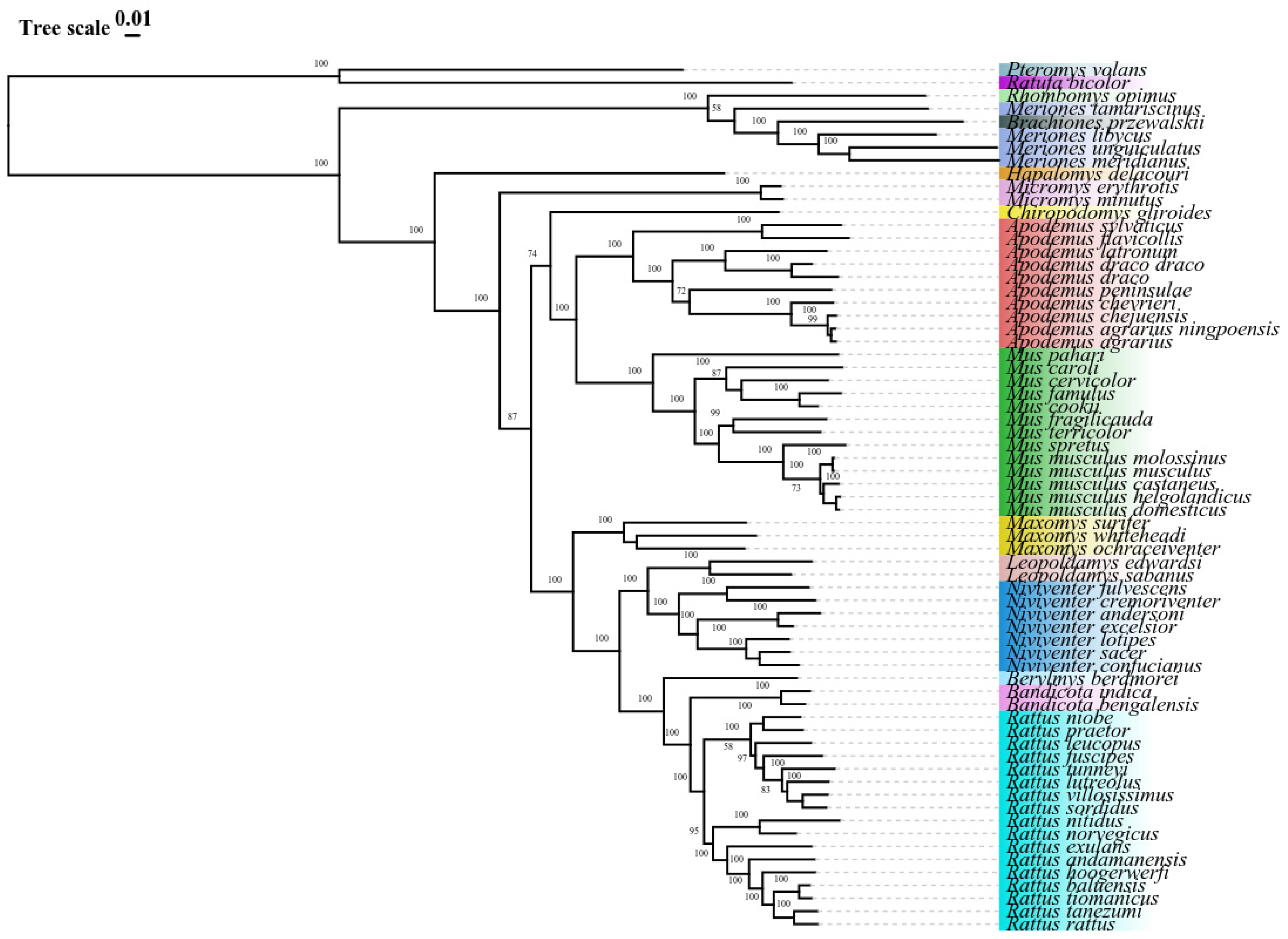
3.4. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tokumasa, H. An introduction to molecular phylogenetic analysis. Rev. Agric. Sci. 2016, 4, 36–45. [Google Scholar] [CrossRef]
- Kryštufek, B.; Lužnik, M.; Buzan, E.V. Mitochondrial cytochrome b sequences resolve the taxonomy of field mice (Apodemus) in the western Balkan refugium. Acta Theriol. 2012, 57, 1–7. [Google Scholar] [CrossRef]
- Atopkin, D.M.; Bogdanov, A.S.; Chelomina, G.N. Genetic variation and differentiation in striped field mouse Apodemus agrarius inferred from RAPD-PCR analysis. Genetika 2007, 43, 804–817. [Google Scholar] [CrossRef]
- Yue, H.; Fan, Z.; Liu, S.; Liu, Y.; Song, Z.; Zhang, X. A mitogenome of the chevrier’s field mouse (Apodemus Chevrieri) and genetic variations inferred from the cytochrome b gene. DNA Cell Biol. 2012, 31, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Jing, M.; Yu, H.; Wu, S.; Wang, W.; Zheng, X. Phylogenetic relationships in genus Niviventer (Rodentia: Muridae) in China inferred from complete mitochondrial cytochrome b gene. Mol. Phylogenet. Evol. 2007, 44, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Li, F.; Zhou, J.; Lv, J.; Tan, Z.; Zhang, Y.; Ge, X. The phylogeny of the Anderson’s white-bellied rat (Niviventer andersoni) based on complete mitochondrial genomes. Ecol. Evol. 2022, 12, e8663. [Google Scholar] [CrossRef]
- Galtier, N.; Nabholz, B.; Glemin, S.; Hurst, G.D.D. Mitochondrial DNA as a marker of molecular diversity: A reappraisal. Mol. Ecol. 2009, 18, 4541–4550. [Google Scholar] [CrossRef]
- An, J.; Fan, C.; Fu, Z.; Zhang, H.; Yang, P. Analyses of the complete mitochondrial genome of Paraconiothyrium sp. and gene rearrangement diversity in the Pleosporales. Diversity 2022, 14, 601. [Google Scholar] [CrossRef]
- Wang, X.; Pei, J.; Bao, P.; Cao, M.; Guo, S.; Song, R.; Song, W.; Liang, C.; Yan, P.; Guo, X. Mitogenomic diversity and phylogeny analysis of yak (Bos grunniens). BMC Genom. 2021, 22, 325. [Google Scholar] [CrossRef]
- Sullivan, K.A.; Platt, R.N.; Bradley, R.D.; Ray, D.A. Whole mitochondrial genomes provide increased resolution and indicate paraphyly in deer mice. BMC Zool. 2017, 2, 11. [Google Scholar] [CrossRef]
- Kim, H.R.; Park, Y.C. The complete mitochondrial genome of the striped field mouse, Apodemus agrarius (Rodentia, Murinae) from Korea. Mitochondrial DNA 2012, 23, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Feijó, A.; Cheng, J.; Lu, L.; Liu, R.; Abramov, A.V.; Xia, L.; Wen, Z.; Zhang, W.; Shi, L.; et al. Evolutionary history of field mice (Murinae: Apodemus), with emphasis on morphological variation among species in China and description of a new species. Zool. J. Linn. Soc. Lond. 2019, 187, 518–534. [Google Scholar] [CrossRef]
- Moshtaghi, S.; Darvish, J.; Mirshamsi, O.; Mahmoudi, A. Cryptic species diversity in the genus Allactaga (Rodentia: Dipodidae) at the edge of its distribution range. Folia Zool. 2016, 65, 142–147. [Google Scholar] [CrossRef]
- Plese, B.; Kenny, N.J.; Rossi, M.E.; Cárdenas, P.; Schuster, A.; Taboada, S.; Koutsouveli, V.; Koutsouveli, V. Mitochondrial evolution in the Demospongiae (Porifera): Phylogeny, divergence time, and genome biology. Mol. Phylogenet. Evol. 2020, 155, 107011. [Google Scholar] [CrossRef] [PubMed]
- Dowling, D.; Friberg, U.; Lindell, J. Evolutionary implications of non-neutral mitochondrial genetic variation. Trends Ecol. Evol. 2008, 23, 546–554. [Google Scholar] [CrossRef]
- Ye, W.; Wang, J.; Zhao, X.; Liu, H.; Zhu, S. Mitochondrial genomes of two Lycosa spiders (Araneae, Lycosidae): Genome description and phylogenetic implications. Diversity 2022, 14, 538. [Google Scholar] [CrossRef]
- Wu, B.; Zhou, P.; He, J.; Cui, B.; Chen, F.; Sun, L. The taxonomic research of rodents with related target genes. Chin. J. Vector Biol. Control 2016, 27, 520–524. [Google Scholar] [CrossRef]
- Steppan, S.J.; Schenk, J.J. Muroid rodent phylogenetics: 900-species tree reveals increasing diversification rates. PLoS ONE 2017, 12, e183070. [Google Scholar] [CrossRef] [Green Version]
- Bellinvia, E. A phylogenetic study of the genus Apodemus by sequencing the mitochondrial DNA control region. J. Zool. Syst. Evol. Res. 2004, 42, 289–297. [Google Scholar] [CrossRef]
- Yalkovskaya, L.; Sibiryakov, P.; Borodin, A. Phylogeography of the striped field mouse (Apodemus agrarius Pallas, 1771) in light of new data from central part of Northern Eurasia. PLoS ONE 2022, 17, e276466. [Google Scholar] [CrossRef]
- Moritz, C.; Dowling, T.E.; Brown, W.M. Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annu. Rev. Ecol. Syst. 1987, 18, 269–292. [Google Scholar] [CrossRef]
- Ding, L.; Luo, G.; Zhou, Q.; Sun, Y.; Liao, J. Comparative mitogenome analysis of gerbils and the mitogenome phylogeny of Gerbillinae (Rodentia: Muridae). Biochem. Genet. 2022, 60, 2226–2249. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Yan, C.; Tu, F.; Yang, C.; Ma, W.; Fan, Z.; Song, Z.; Owens, J.; Liu, S.; Zhang, X. Two novel mitogenomes of Dipodidae species and phylogeny of Rodentia inferred from the complete mitogenomes. Biochem. Syst. Ecol. 2015, 60, 123–130. [Google Scholar] [CrossRef]
- İbiş, O. Whole mitochondrial genome sequence and phylogenetic relationships of Williams’s jerboa (Scarturus williamsi) from Turk. PeerJ 2020, 8, e9569. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Y.; Xiao, Q.; Lin, Y.; Du, Y.; Lin, C.; Ward-Fear, G.; Hu, C.; Qu, Y.; Li, H. Characterization of the complete mitochondrial genome of the many-lined sun skink (Eutropis multifasciata) and comparison with other Scincomorpha species. Genomics 2021, 113, 2526–2536. [Google Scholar] [CrossRef]
- Oh, D.; Kim, T.; Chang, M.; Han, S.; Oh, H.; Kim, S. Mitochondrial genome sequences of the striped field mice Apodemus agrarius coreae and Apodemus agrarius chejuensis. Mitochondrial DNA 2011, 22, 165–167. [Google Scholar] [CrossRef]
- Wei, H.; Jia, Q.; Li, F.; Liu, Y.; Chen, S.; Yong, B. Sequencing and analysis of complete mitochondrial genome of Apodemus draco (Rodentia: Arvicolinae). Mitochondrial DNA Part A 2016, 27, 2740–2741. [Google Scholar] [CrossRef]
- Liu, X.; Wei, F.; Li, M.; Jiang, X.; Feng, Z.; Hu, J. Molecular phylogeny and taxonomy of wood mice (genus Apodemus Kaup, 1829) based on complete mtDNA cytochrome b sequences, with emphasis on Chinese species. Mol. Phylogenet. Evol. 2004, 33, 1–15. [Google Scholar] [CrossRef]
- Liu, S.; He, K.; Chen, S.; Jin, W.; Murphy, R.W.; Tang, M.; Liao, R.; Li, F. How many species of Apodemus and Rattus occur in China? A survey based on mitochondrial cyt b and morphological analyses. Zool. Res. 2018, 39, 309–320. [Google Scholar] [CrossRef]
- Xiong, Y.; You, F.; Chen, X.; Chen, Y.; Wen, Y.; Chen, Q. Detection and phylogenetic analysis of porcine bocaviruses carried by murine rodents and house shrews in China. Transbound. Emerg. Dis. 2018, 66, 259–267. [Google Scholar] [CrossRef]
- Dubey, S.; Michaux, J.; Brünner, H.; Hutterer, R.; Vogel, P. False phylogenies on wood mice due to cryptic cytochrome-b pseudogene. Mol. Phylogenet. Evol. 2009, 50, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Braband, A.; Schierwater, B.; Stadler, P.F. Genetic aspects of mitochondrial genome evolution. Mol. Phylogenet. Evol. 2013, 69, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 3, 353–358. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2019, 20, 348–355. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. Dnasp v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Yang, Q.; Wu, Y.; Jiang, X.; Liu, S.; Li, B.; Yang, G.; Li, M.; Zhou, J.; Li, S.; et al. Catalogue of mammals in China (2021). Acta Theriol. Sin. 2021, 41, 487–501. [Google Scholar] [CrossRef]
- Song, S.; Yong, H.; Lim, P.; Tan, J. Mitochondrial genome of Rattus tiomanicus (Rodentia: Muridae) and molecular phylogeny of Murinae. Sains Malays. 2021, 50, 953–965. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Balakirev, A.E.; Abramov, A.V.; Rozhnov, V.V. Revision of the genus Leopoldamys (Rodentia, Muridae) as inferred from morphological and molecular data, with a special emphasis on the species composition in continental Indochina. Zootaxa 2013, 3640, 521–549. [Google Scholar] [CrossRef] [Green Version]
- Reyes, A.; Gissi, C.; Pesole, G.; Saccone, C. Asymmetrical directional mutation pressure in the mitochondrial genome of mammals. Mol. Biol. Evol. 1998, 15, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Triant, D.A.; DeWoody, J.A. Accelerated molecular evolution in Microtus (Rodentia) as assessed via complete mitochondrial genome sequences. Genetica 2006, 128, 95–108. [Google Scholar] [CrossRef]
- Yakovchuk, P.; Protozanova, E.; Frank-Kamenetski, M.D. Base-stacking and base-pairing contributions into thermal stability of the DNA double helix. Nucleic Acids Res. 2006, 34, 564–574. [Google Scholar] [CrossRef]
- Charneski, C.A.; Honti, F.; Bryant, J.M.; Hurst, L.D.; Feil, E.J. Atypical AT skew in firmicute genomes results from selection and not from mutation. PLoS Genet. 2011, 7, e1002283. [Google Scholar] [CrossRef] [Green Version]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef]
- Wei, H.; Li, F.; Wang, X.; Wang, Q.; Chen, G.; Zong, H.; Chen, S. The characterization of complete mitochondrial genome and phylogenetic relationship within Rattus genus (Rodentia: Muridae). Biochem. Syst. Ecol. 2017, 71, 179–186. [Google Scholar] [CrossRef]
- Jiang, X.; Gao, J.; Ni, L.; Hu, J.; Li, K.; Sun, F.; Xie, J.; Bo, X.; Gao, C.; Xiao, J.; et al. The complete mitochondrial genome of Microtus fortis calamorum (Arvicolinae, Rodentia) and its phylogenetic analysis. Gene 2012, 498, 288–295. [Google Scholar] [CrossRef]
- Wolstenholme, D.R. Animal mitochondria DNA: Structure and evolution. Int. Rev. Cytol. 1992, 141, 173–216. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Chen, Q.; Pu, D.; Chen, Z.; Liu, Y.; Liu, X. The first mitochondrial genomes of the family Haplodiplatyidae (Insecta: Dermaptera) reveal intraspecific variation and extensive gene rearrangement. Biology 2022, 11, 807. [Google Scholar] [CrossRef]
- Lavrov, D.V.; Brown, W.M.; Boore, J.L. A novel type of RNA editing occurs in the mitochondrial tRNAs of the centipede Lithobius forficatus. Proc. Natl. Acad. Sci. USA 2000, 97, 13738–13742. [Google Scholar] [CrossRef] [Green Version]
- Lecompte, E.; Aplin, K.; Denys, C.; Catzeflis, F.; Chades, M.; Chevret, P. Phylogeny and biogeography of African Murinae based on mitochondrial and nuclear gene sequences, with a new tribal classification of the subfamily. BMC Evol. Biol. 2008, 1, 199. [Google Scholar] [CrossRef] [Green Version]
- Alhajeri, B.H.; Hunt, O.J.; Steppan, S.J. Molecular systematics of gerbils and deomyines (Rodentia: Gerbillinae, Deomyinae) and a test of desert adaptation in the tympanic bulla. J. Zool. Syst. Evol. Res. 2015, 53, 312–330. [Google Scholar] [CrossRef]
- Ito, M.; Jiang, W.; Sato, J.J.; Zhen, Q.; Jiao, W.; Goto, K.; Sato, H.; Ishiwata, K.; Oku, Y.; Chai, J.; et al. Molecular phylogeny of the subfamily Gerbillinae (Muridae, Rodentia) with emphasis on species living in the Xinjiang-Uygur Autonomous Region of China and based on the mitochondrial cytochrome b and cytochrome c oxidase subunit II genes. Zool. Sci. 2010, 27, 269–278. [Google Scholar] [CrossRef]
- Renaud, S.; Chevret, P.; Michaux, J. Morphological vs. molecular evolution: Ecology and phylogeny both shape the mandible of rodents. Zool. Scr. 2007, 36, 525–535. [Google Scholar] [CrossRef]
- Motokawa, M.; Wu, Y.; Harada, M.; Shintaku, Y.; Jiang, X.L.; Li, Y.C. Karyotypes of field mice of the genus Apodemus (Mammalia: Rodentia) from China. Zool. Res. 2018, 39, 348–355. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Accession No. |
|---|---|---|
| Apodemus | Apodemus chevrieri | NC_017599.1 |
| Apodemus | Apodemus latronum | NC_019585.1 |
| Apodemus | Apodemus flavicollis | MN122902.1 |
| Apodemus | Apodemus peninsulae | NC_016060.1 |
| Apodemus | Apodemus draco | NC_019584.1 |
| Apodemus | Apodemus chejuensis | NC_016662.1 |
| Apodemus | Apodemus agrarius | NC_016428.1 |
| Apodemus | Apodemus sylvaticus | NC_049122.1 |
| Rattus | Rattus andamanensis | NC_046686.1 |
| Rattus | Rattus tiomanicus | NC_029888.1 |
| Rattus | Rattus hoogerwerfi | NC_049040.1 |
| Rattus | Rattus baluensis | NC_035621.1 |
| Rattus | Rattus sordidus | NC_014871.1 |
| Rattus | Rattus lutreolus | NC_014858.1 |
| Rattus | Rattus villosissimus | NC_014864.1 |
| Rattus | Rattus tunneyi | NC_014861.1 |
| Rattus | Rattus fuscipes | NC_014867.1 |
| Rattus | Rattus leucopus | NC_014855.1 |
| Rattus | Rattus niobe | NC_023347.1 |
| Rattus | Rattus rattus | NC_012374.1 |
| Rattus | Rattus praetor | NC_012461.1 |
| Rattus | Rattus norvegicus | NC_001665.2 |
| Rattus | Rattus tanezumi | NC_011638.1 |
| Rattus | Rattus exulans | NC_012389.1 |
| Rattus | Rattus nitidus | NC_040919.1 |
| Hapalomys | Hapalomys delacouri | MZ159976.1 |
| Niviventer | Niviventer cremoriventer | NC_035822.1 |
| Niviventer | Niviventer lotipes | NC_065402.1 |
| Niviventer | Niviventer sacer | MZ935252.1 |
| Niviventer | Niviventer excelsior | NC_019617.1 |
| Niviventer | Niviventer confucianus | NC_023960.1 |
| Niviventer | Niviventer fulvescens | NC_028715.1 |
| Niviventer | Niviventer andersoni | NC_060500.1 |
| Leopoldamys | Leopoldamys sabanus | NC_035819.1 |
| Leopoldamys | Leopoldamys edwardsi | NC_025670.1 |
| Mus | Mus musculus musculus | NC_010339.1 |
| Mus | Mus cervicolor | NC_025269.1 |
| Mus | Mus cookii | NC_025270.1 |
| Mus | Mus fragilicauda | NC_025287.1 |
| Mus | Mus terricolor | NC_010650.1 |
| Mus | Mus musculus helgolandicus | KP877610.1 |
| Mus | Mus musculus castaneus | NC_012387.1 |
| Mus | Mus musculus domesticus | NC_006914.1 |
| Mus | Mus musculus molossinus | NC_006915.1 |
| Mus | Mus spretus | NC_025952.1 |
| Mus | Mus famulus | NC_030342.1 |
| Mus | Mus pahari | NC_036680.1 |
| Mus | Mus caroli | NC_025268.1 |
| Berylmys | Berylmys berdmorei | NC_036730.1 |
| Brachiones | Brachiones przewalskii | KT834972.1 |
| Maxomys | Maxomys ochraceiventer | NC_056988.1 |
| Maxomys | Maxomys surifer | NC_036732.1 |
| Maxomys | Maxomys whiteheadi | NC_049119.1 |
| Bandicota | Bandicota bengalensis | NC_057104.1 |
| Bandicota | Bandicota indica | KT029807.1 |
| Micromys | Micromys erythrotis | NC_060316.1 |
| Micromys | Micromys minutus | NC_027932.1 |
| Chiropodomys | Chiropodomys gliroides | NC_049121.1 |
| Meriones | Meriones unguiculatus | KF425526.1 |
| Meriones | Meriones tamariscinus | NC_034314.1 |
| Meriones | Meriones meridianus | NC_027684.1 |
| Meriones | Meriones libycus | NC_027683.1 |
| Rhombomys | Rhombomys opimus | MK359635.1 |
| Pteromys | Pteromys volans | NC_019612.1 |
| Ratufa | Ratufa bicolor | NC_023780.1 |
| Gene | Direction | Position (bp) | Size (bp) | Start Codon | Stop Codon | Intergenic Nucleotides |
|---|---|---|---|---|---|---|
| tRNAPhe | + | 1–67\1–68 | 67\68 | 0\0 | ||
| 12S rRNA | + | 68–1022\69–1023 | 955\955 | 0\0 | ||
| tRNAVal | + | 1023–1091\1024–1091 | 69\68 | 0\0 | ||
| 16S rRNA | + | 1092–2665\1092–2660 | 1547\1569 | 0\0 | ||
| tRNALeu | + | 2664–2738\2659–2733 | 75\75 | −2\−2 | ||
| ND1 | + | 2736–3693\2731–3688 | 958\958 | ATA\ATA | T-\T- | −3\−3 |
| tRNAIle | + | 3694–3762\3689–3757 | 69\69 | 0\0 | ||
| tRNAGln | − | 3760–3830\3755–3825 | 71\71 | −3\−3 | ||
| tRNAMet | + | 3835–3903\3830–3898 | 69\69 | 4\4 | ||
| ND2 | + | 3094–4939\3899–4934 | 1036\1036 | ATA\ATA | T-\T- | 0\0 |
| tRNATrp | + | 4940–5007\4935–5002 | 68\68 | 0\0 | ||
| tRNAAla | − | 5009–5077\5004–5072 | 69\69 | 1\1 | ||
| tRNAAsn | − | 5079–5149\5074–5144 | 71\71 | 1\1 | ||
| tRNACys | − | 5181–5248\5176–5241 | 68\66 | 31\31 | ||
| tRNATyr | − | 5249–5315\5242–5308 | 67\67 | 0\0 | ||
| COX1 | + | 5317–6859\5310–6852 | 1543\1543 | ATG\ATG | T-\T- | 1\1 |
| tRNASer | − | 6859–6927\6852–6920 | 69\69 | −1\−1 | ||
| tRNAAsp | + | 6931–6998\6924–6991 | 68\68 | 3\3 | ||
| COX2 | + | 7000–7683\6993–7676 | 684\684 | ATG\ATG | TAA\TAA | 1\1 |
| tRNALys | + | 7687–7750\7680–7743 | 64\64 | 3\3 | ||
| ATP8 | + | 7752–7955\7745–7948 | 204\204 | ATG\ATG | TAA\TAA | 1\1 |
| ATP6 | + | 7913–8593\7906–8586 | 681\681 | ATG\ATG | TAA\TAA | −43\−43 |
| COX3 | + | 8593–9376\8586–9369 | 784\784 | ATG\ATG | T-\T- | −1\−1 |
| tRNAGly | + | 9377–9444\9370–9436 | 68\67 | 0\0 | ||
| ND3 | + | 9445–9792\9437–9784 | 348\348 | ATA\ATT | TAA\TAA | 0\0 |
| tRNAArg | + | 9794–9861\9786–9853 | 68\68 | 1\1 | ||
| ND4L | + | 9864–10160\9856–10152 | 297\297 | ATG\ATG | TAA\TAA | 2\2 |
| ND4 | + | 10154–11531\10146–11523 | 1378\1378 | ATG\ATG | T-\T- | −7\−7 |
| tRNAHis | + | 11532–11599\11524–11590 | 68\67 | 0\0 | ||
| tRNASer | + | 11600–11658\11591–11649 | 59\59 | 0\0 | ||
| tRNALeu | + | 11658–11728\11649–11719 | 71\71 | −1\−1 | ||
| ND5 | + | 11720–13549\11711–13540 | 1830\1830 | ATA\ATA | TAA\TAA | −9\−9 |
| ND6 | − | 13536–14054\13527–14042 | 519\516 | ATG\ATG | TAA\TAA | −14\−14 |
| tRNAGlu | − | 14055–14123\14043–14111 | 69\69 | 0\0 | ||
| CYTB | + | 14127–15270\14115–15258 | 1144\1144 | ATG\ATG | T-\T- | 3\3 |
| tRNAThr | + | 15271–15339\15259–15326 | 69\68 | 0\0 | ||
| tRNAPro | − | 15340–15406\15327–15393 | 67\67 | 0\0 | ||
| D-loop | + | 15407–16262\15394–16222 | 856\829 | 0\0 |
| Apodemus agrarius ningpoensis | Size (bp) | A% | T% | G% | C% | A + T% | G + C% | AT Skew | GC Skew |
|---|---|---|---|---|---|---|---|---|---|
| Mitogenome | 16,262 | 34.01 | 29.74 | 12.47 | 23.79 | 63.74 | 36.26 | 0.067 | −0.312 |
| PCGs | 11,406 | 32.05 | 31.45 | 12.1 | 24.4 | 63.5 | 36.5 | 0.01 | −0.337 |
| tRNAs | 1503 | 34.13 | 30.81 | 18.43 | 16.63 | 64.94 | 35.06 | 0.051 | 0.051 |
| rRNAs | 2529 | 37.92 | 26.49 | 16.77 | 18.82 | 64.41 | 35.59 | 0.177 | −0.058 |
| D-loop | 856 | 32.71 | 31.66 | 11.92 | 23.71 | 64.37 | 35.63 | 0.016 | −0.331 |
| Apodemus draco draco | Size (bp) | A% | T% | G% | C% | A + T% | G + C% | AT Skew | GC Skew |
|---|---|---|---|---|---|---|---|---|---|
| Mitogenome | 16,222 | 35.15 | 28.97 | 12.08 | 23.8 | 64.12 | 35.88 | 0.096 | −0.327 |
| PCGs | 11,403 | 33.33 | 30.64 | 11.68 | 24.34 | 63.97 | 36.03 | 0.042 | −0.352 |
| tRNAs | 1498 | 34.31 | 31.51 | 18.29 | 15.89 | 65.82 | 34.18 | 0.043 | 0.07 |
| rRNAs | 2524 | 39.26 | 25.79 | 16.32 | 18.62 | 65.06 | 34.94 | 0.207 | −0.066 |
| D-loop | 829 | 31.97 | 29.67 | 13.15 | 25.21 | 61.64 | 38.36 | 0.037 | −0.314 |
| Codon | No. | RSCU | Codon | No. | RSCU | Codon | No. | RSCU |
|---|---|---|---|---|---|---|---|---|
| UAA(*) | 7 | 4 | AAA(K) | 98 | 1.98 | CGU(R) | 10 | 0.62 |
| GCA(A) | 93 | 1.62 | AAG(K) | 1 | 0.02 | AGC(S) | 36 | 0.72 |
| GCC(A) | 66 | 1.15 | CUA(L) | 258 | 2.58 | AGU(S) | 20 | 0.4 |
| GCG(A) | 4 | 0.07 | CUC(L) | 80 | 0.8 | UCA(S) | 138 | 2.77 |
| GCU(A) | 67 | 1.17 | CUG(L) | 19 | 0.19 | UCC(S) | 54 | 1.08 |
| UGC(C) | 20 | 1.38 | CUU(L) | 109 | 1.09 | UCG(S) | 2 | 0.04 |
| UGU(C) | 9 | 0.62 | UUA(L) | 123 | 1.23 | UCU(S) | 49 | 0.98 |
| GAC(D) | 37 | 1.04 | UUG(L) | 10 | 0.1 | ACA(T) | 161 | 2.13 |
| GAU(D) | 34 | 0.96 | AUA(M) | 198 | 1.7 | ACC(T) | 60 | 0.79 |
| GAA(E) | 78 | 1.68 | AUG(M) | 35 | 0.3 | ACG(T) | 4 | 0.05 |
| GAG(E) | 15 | 0.32 | AAC(N) | 92 | 1.1 | ACU(T) | 78 | 1.03 |
| UUC(F) | 97 | 0.79 | AAU(N) | 76 | 0.9 | GUA(V) | 85 | 2.01 |
| UUU(F) | 148 | 1.21 | CCA(P) | 125 | 2.51 | GUC(V) | 25 | 0.59 |
| GGA(G) | 101 | 1.92 | CCC(P) | 28 | 0.56 | GUG(V) | 9 | 0.21 |
| GGC(G) | 40 | 0.76 | CCG(P) | 3 | 0.06 | GUU(V) | 50 | 1.18 |
| GGG(G) | 26 | 0.5 | CCU(P) | 43 | 0.86 | UGA(W) | 93 | 1.81 |
| GGU(G) | 43 | 0.82 | CAA(Q) | 77 | 1.88 | UGG(W) | 10 | 0.19 |
| CAC(H) | 53 | 1.1 | CAG(Q) | 5 | 0.12 | UAC(Y) | 60 | 0.92 |
| CAU(H) | 43 | 0.9 | CGA(R) | 45 | 2.81 | UAU(Y) | 70 | 1.08 |
| AUC(I) | 146 | 0.79 | CGC(R) | 7 | 0.44 | |||
| AUU(I) | 225 | 1.21 | CGG(R) | 2 | 0.12 |
| Codon | No. | RSCU | Codon | No. | RSCU | Codon | No. | RSCU |
|---|---|---|---|---|---|---|---|---|
| UAA(*) | 7 | 4 | AUU(I) | 216 | 1.16 | CGU(R) | 7 | 0.44 |
| GCA(A) | 89 | 1.6 | AAG(K) | 3 | 0.06 | CGA(R) | 46 | 2.88 |
| GCC(A) | 76 | 1.36 | AAA(K) | 99 | 1.94 | AGU(S) | 10 | 0.2 |
| GCG(A) | 1 | 0.02 | CUC(L) | 66 | 0.67 | UCA(S) | 163 | 3.21 |
| GCU(A) | 57 | 1.02 | CUG(L) | 8 | 0.08 | UCC(S) | 37 | 0.73 |
| UGC(C) | 16 | 1.1 | CUU(L) | 104 | 1.06 | UCG(S) | 5 | 0.1 |
| UGU(C) | 13 | 0.9 | UUA(L) | 134 | 1.37 | UCU(S) | 46 | 0.9 |
| GAC(D) | 48 | 1.33 | UUG(L) | 13 | 0.13 | AGC(S) | 44 | 0.87 |
| GAU(D) | 24 | 0.67 | CUA(L) | 264 | 2.69 | ACC(T) | 71 | 0.92 |
| GAA(E) | 84 | 1.79 | AUG(M) | 28 | 0.23 | ACG(T) | 4 | 0.25 |
| GAG(E) | 10 | 0.21 | AUA(M) | 211 | 1.77 | ACU(T) | 61 | 0.79 |
| UUC(F) | 116 | 0.95 | AAU(N) | 72 | 0.88 | ACA(T) | 173 | 2.24 |
| GGA(G) | 118 | 2.26 | CCC(P) | 16 | 0.32 | GUG(V) | 10 | 0.24 |
| GGC(G) | 27 | 0.52 | CCU(P) | 36 | 0.73 | GUU(V) | 38 | 0.91 |
| GGG(G) | 19 | 0.36 | CCA(P) | 145 | 2.94 | GUA(V) | 101 | 2.42 |
| GGU(G) | 45 | 0.86 | CAG(Q) | 1 | 0.02 | UGA(W) | 98 | 1.9 |
| CAC(H) | 60 | 1.21 | CAA(Q) | 81 | 1.98 | UGG(W) | 5 | 0.1 |
| CAU(H) | 39 | 0.79 | CGC(R) | 10 | 0.62 | UAC(Y) | 59 | 0.91 |
| AUC(I) | 157 | 0.84 | CGG(R) | 1 | 0.06 | UAU(Y) | 70 | 1.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, D.; Zhou, L.; Xue, J.; Xia, Q.; Meng, L. Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae. Diversity 2022, 14, 1089. https://doi.org/10.3390/d14121089
Wu D, Zhou L, Xue J, Xia Q, Meng L. Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae. Diversity. 2022; 14(12):1089. https://doi.org/10.3390/d14121089
Chicago/Turabian StyleWu, Di, Lizhi Zhou, Jiezhong Xue, Qiliang Xia, and Lei Meng. 2022. "Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae" Diversity 14, no. 12: 1089. https://doi.org/10.3390/d14121089
APA StyleWu, D., Zhou, L., Xue, J., Xia, Q., & Meng, L. (2022). Characterization of Two New Apodemus Mitogenomes (Rodentia: Muridae) and Mitochondrial Phylogeny of Muridae. Diversity, 14(12), 1089. https://doi.org/10.3390/d14121089