
Thermal Tolerance of Cyclops bohater (Crustacea: Copepoda); Selection of Optimal and Avoided Conditions in Experimental Conditions

 ,
,  ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
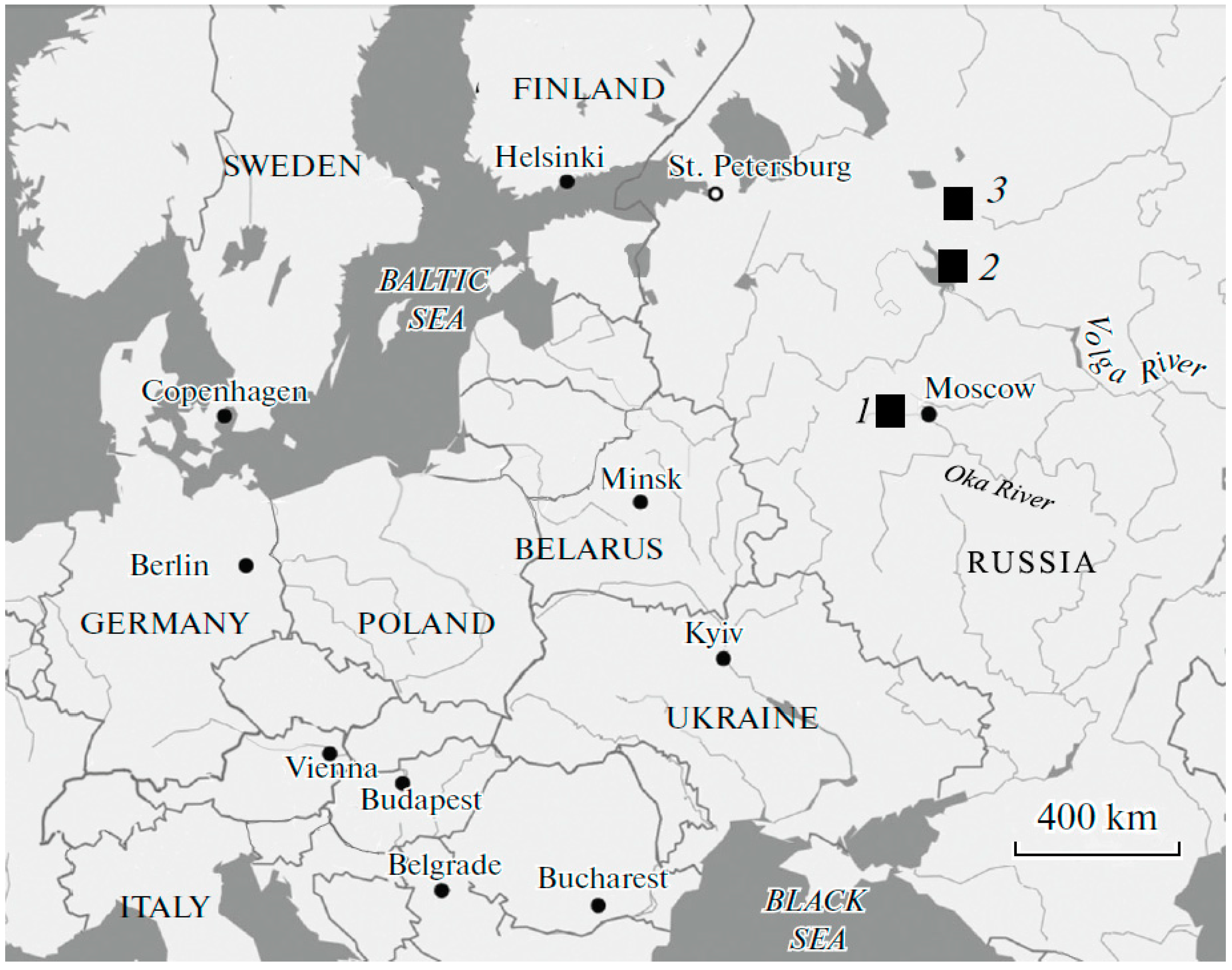
2.1. Water Bodies in Which Field Distributions Were Analyzed
2.2. Collection of Zooplankton in the Field
2.3. Molecular Analysis
2.4. Zooplankton Collection for Testing
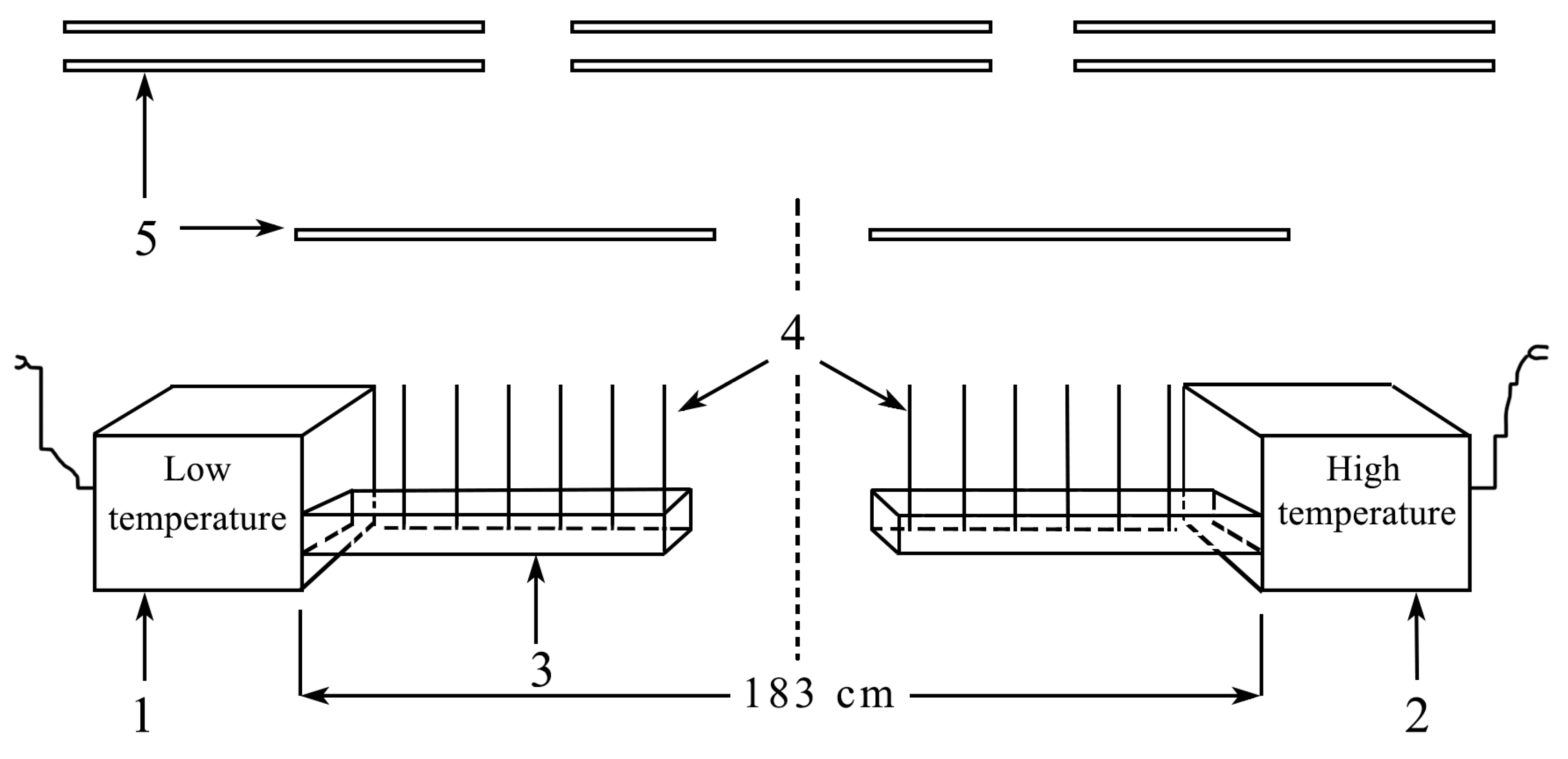
2.5. Description of Thermogradient Apparatus
2.6. Experimental Procedures
2.7. Calculation of Preferred and Avoided Temperatures
3. Results
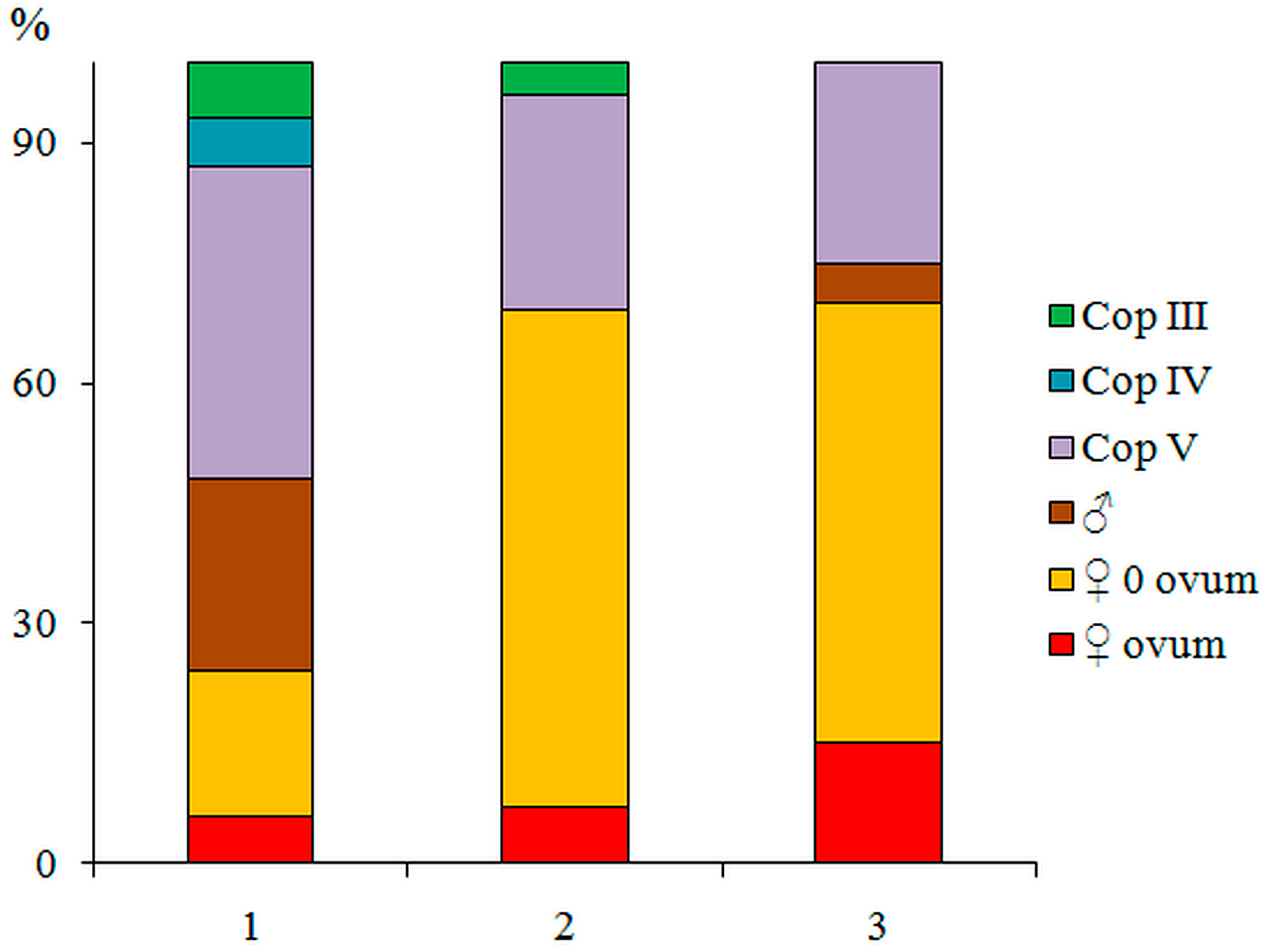
3.1. Field Observation on the Distribution of C. bohater
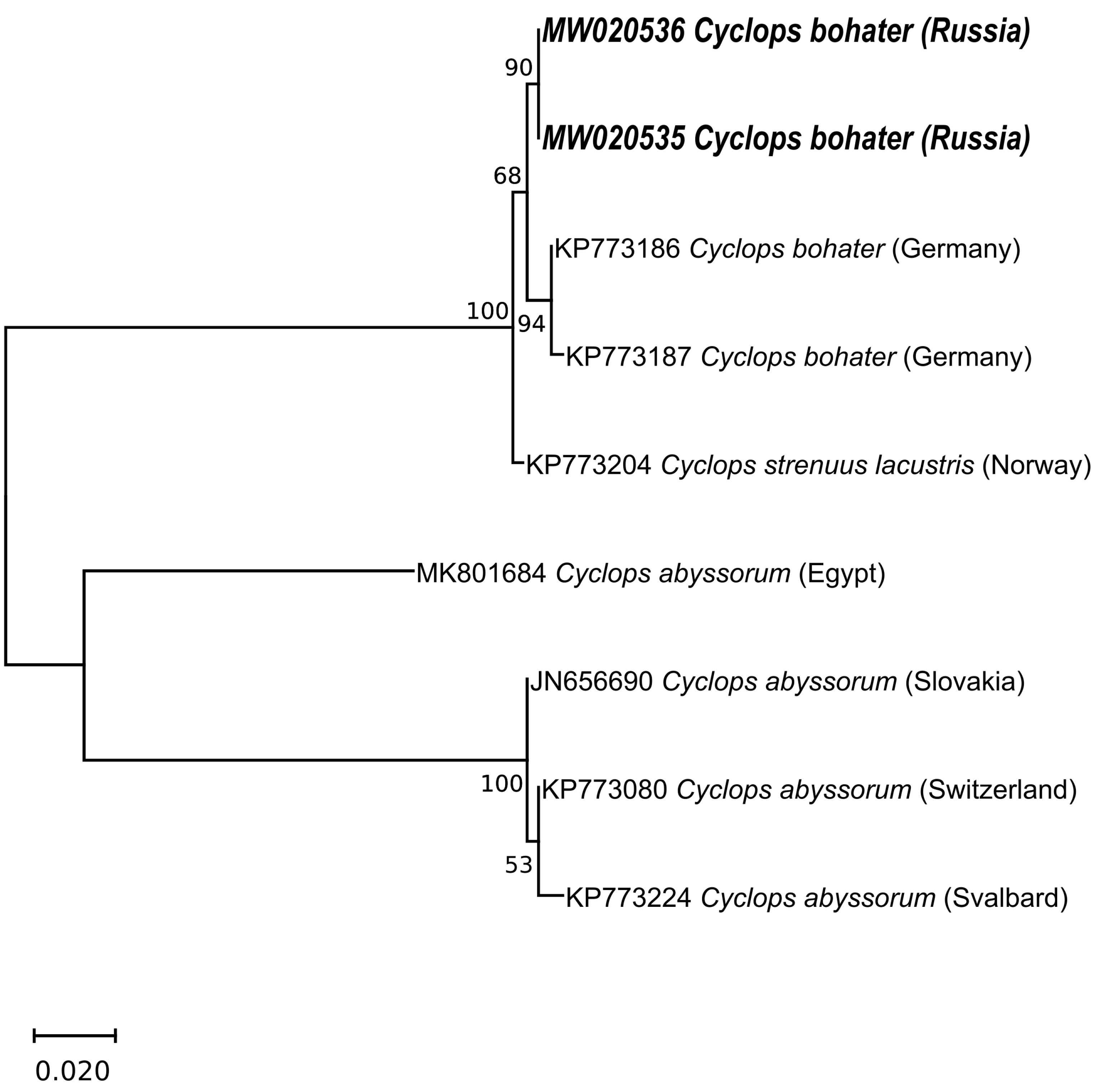
3.2. Molecular Data
3.3. Experimental Data
3.3.1. CTM Determination
3.3.2. Determination of FTP and TNP
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, W.W.; Casterlin, M.E. Behavioral thermoregulation and the “final preferendum” paradigm. Am. Zool. 1979, 19, 211–224. [Google Scholar] [CrossRef]
- Beitinger, T.L.; Bennett, W.A.; McCauley, R.W. Temperature tolerances of North American freshwater fishes exposed to dynamic changes in temperature. Environ. Biol. Fishes 2000, 58, 237–275. [Google Scholar] [CrossRef]
- Sunday, J.; Bates, A.E.; Dulvy, N. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [Green Version]
- Geraldes, A.M.; Boavida, M.J. What factors affect the pelagic cladocerans of the meso-eutrophic Azibo Reservoir? Ann. Limnol. 2004, 40, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Verbitsky, V.B.; Verbitskaya, T.I. Ecological optimum and effect of delayed action of a factor. Dokl. Biol. Sci. 2007, 416, 386–388. [Google Scholar] [CrossRef]
- Geerts, A.N.; Vanoverbeke, J.; Vanschoenwinkel, B.; Van Doorslaer, W.; Feuchtmayr, H.; Atkinson, D.; Moss, B.; Davidson, T.A.; Sayer, C.D.; De Meester, L. Rapid evolution of thermal tolerance in the water flea Daphnia. Nat. Clim. Chang. 2015, 5, 665–668. [Google Scholar] [CrossRef] [Green Version]
- Sunday, J.; Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Gravel, S.; Hargreaves, A.L.; Leiva, F.P.; Verberk, W.C.E.P.; Olalla-Tarraga, M.A.; Morales-Castilla, I. Thermal tolerance patterns across latitude and elevation. Phil. Trans. R. Soc. B 2019, 374, 20190036. [Google Scholar] [CrossRef] [Green Version]
- Blenckner, T.; Adrian, R.; Livingstone, D.M.; Jennings, E.; Weyenmeyer, G.; George, D.G.; Jankowski, T.; Jarvinen, M.; Aonghusa, C.N.; Noges, T.; et al. Large-scale climatic signatures in lakes across Europe: A meta-analysis. Glob. Chang. Biol. 2007, 13, 1314–1326. [Google Scholar] [CrossRef] [Green Version]
- Cavieres, G.; Bogdanovich, J.M.; Bozinovic, F. Ontogenetic thermal tolerance and performance of ectotherms at variable temperatures. J. Evol. Biol. 2016, 29, 1462–1468. [Google Scholar] [CrossRef]
- Nichelmann, M. Some characteristics of the biological optimum temperature. J. Therm. Biol. 1983, 8, 69–71. [Google Scholar] [CrossRef]
- Dïaz, F.; Sierra, E.; Deniss, A.; Rodriguez, L. Behavioural thermoregulation and critical thermal limits of Macrobrachium acanthurus (Wiegman). J. Therm. Biol. 2002, 27, 423–428. [Google Scholar] [CrossRef]
- Hernandez, R.M.; Bückle, R.L.F. Thermal preference area for Macrobrachium tenellum in the context of global climatic change. J. Therm. Biol. 1997, 22, 309–313. [Google Scholar] [CrossRef]
- Angilletta, M.J. Thermal Adaptation. A Theoretical and Empirical Synthesis; Oxford University Press Inc.: New York, NY, USA, 2009; p. 302. [Google Scholar] [CrossRef]
- Lamkemeyer, T.; Zeis, B.; Paul, R.J. Temperature acclimation influences temperature related behaviour as well as oxygen transport physiology and biochemistry in the water flea Daphnia magna. Can. J. Zool. 2003, 81, 237–249. [Google Scholar] [CrossRef]
- Angilletta, M.J.; Bennett, A.F.; Guderly, H.; Navas, C.A.; Seebacher, F.; Wilson, R.S. Coadaptation: A unifying principle in evolutionary thermal biology. Physiol. Biochem. Zool. 2006, 79, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Tepler, S.; Mach, K.; Denny, M. Preference versus performance: Body temperature of the intertidal snail Chlorostoma funebralis. Biol. Bull. 2011, 220, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Verbitskii, V.B.; Verbitskaya, T.I. Thermal preference and avoidance in Cladoceran Daphnia magna Strauss (Crustacea, Cladocera) acclimated to constant temperature. Biol. Bull. Russ. 2012, 39, 93–98. [Google Scholar] [CrossRef]
- Paladino, V.K.; Spotila, J.R.; Schubauer, J.P.; Kowalski, K.T. The critical thermal maximum: A technique used to elucidate physiological stress and adaptation in fish. Can. Biol. Rev. 1980, 39, 115–122. [Google Scholar]
- Diaz, F.; Buckle, L.F. Effect of the critical thermal maximum on the preferred temperatures of Ictalurus punctatus exposed to constant and fluctuating temperatures. J. Therm. Biol. 1999, 24, 155–160. [Google Scholar] [CrossRef]
- Koźmiński, Z. Morphometrische und ökologische Untersuchungen an Cyclopiden der strenuus-Gruppe. Int. Rev. Ges. Hydrobiol. Hydrogr. 1936, 33, 161–240. [Google Scholar] [CrossRef]
- Monchenko, V.I. Gnathostomatous Cyclopoida. Cyclops (Cyclopidae). Fauna Ukraine; Markevich, O.P., Ed.; Naukova Dumka: Kyiv, Ukraine, 1974; pp. 192–194. (In Ukrainian) [Google Scholar]
- Holynska, M.; Dimante-Deimantovica, I. Redescription and taxonomic notes on Cyclops bohater Koźmiński, 1933 and Cyclops lacustris G.O. Sars, 1863 (Arthropoda, Crustacea), with an identification key to the Cyclops species of Fenno-Scandinavia. Eur. J. Taxon. 2016, 212, 1–31. [Google Scholar] [CrossRef]
- Holynska, M.; Wyngaard, G.A. Towards a phylogeny of Cyclops (Copepoda): (in) congruences among morphology, molecules and zoogeography. Zool. Scr. 2019, 48, 376–398. [Google Scholar] [CrossRef]
- Lazareva, V.I.; Zhdanova, S.M. Copepod Cyclops bohater (Crustacea, Copepoda) in the European Part of Russia. Inland Water Biol. 2020, 13, 528–538. [Google Scholar] [CrossRef]
- Frisch, D. Dormancy, dispersal and the survival of cyclopoid copepods (Cyclopoida, Copepoda) in a lowland floodplain. Freshw. Biol. 2002, 47, 1269–1281. [Google Scholar] [CrossRef] [Green Version]
- Rivier, I.K. Cold-Water Zooplankton of Lakes of the Upper Volga Basin; Yakovlev, V.N., Ed.; Publisher Permyakov: Izhevsk, Russia, 2012; p. 390. (In Russian) [Google Scholar]
- Einsle, U. Revision der Gattung Cyclops s. str. speziell der abyssorum–gruppe. Mem. Ist. Ital. Idrobiol. Dott. Marco De Marchi 1975, 32, 57–219. [Google Scholar]
- Einsle, U. The long-term dynamics of crustacean communities in Lake Constance (Obersee, 1962–1986). Schweiz. Z. Hydrol. 1988, 50, 136–165. [Google Scholar] [CrossRef]
- Butorin, N.V.; Mordukhai-Boltovskoy, P.D. (Eds.) The River Volga and Its Life; Junk Publishers: The Hague, The Netherlands; Boston, MA, USA; London, UK, 1979. [Google Scholar]
- Holynska, M.; Dahms, H.-U. New diagnostic microcharacters of the cephalothoracic appendages in Cyclops O. F. Müller, 1776 (Crustacea, Copepoda, Cyclopoida). Zoosystema 2004, 26, 175–198. [Google Scholar]
- Holynska, M. On the morphology and geographical distribution of some problematic South Palearctic Cyclops (Copepoda: Cyclopidae). J. Nat. Hist. 2008, 42, 2011–2039. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Machida, R.J.; Miya, M.U.; Nishida, M.; Nishida, S. Large-scale gene arrangements in the mitochondrial genomes of two calanoid copepods Eucalanus bungii and Neocalanus cristatus (Crustacea), with notes on new versatile primers for the srRNA and COI genes. Gene 2004, 332, 71–78. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Krajiček, M.; Fott, J.; Miracle, M.R.; Ventura, M.; Sommaruga, R.; Kirschner, P.; Černy, M. The genus Cyclops (Copepoda, Cyclopoida) in Europe. Zool. Scr. 2016, 45, 671–682. [Google Scholar] [CrossRef]
- El-khodary, G.; Mona, M.M.; El-sayed, H.S.; Ghoneim, A.Z. Phylogenetic identification and assessment of the nutritional value of different diets for a copepod species isolated from Eastern Harbor coastal region. Egypt. J. Aquat. Res. 2020, 46, 173–180. [Google Scholar] [CrossRef]
- Rosetti, Y.; Rosetti, L.; Cabanac, M. Annual oscillation of preferred temperature in the freshwater snail Lymnaea awicularia: Effect of light and temperature. Anim. Behav. 1989, 37, 897–907. [Google Scholar] [CrossRef]
- Verbitsky, V.B.; Lazareva, V.I.; Medyantseva, E.N.; Malysheva, O.A.; Zhdanova, S.M.; Verbitskaya, T.I.; Grishanin, A.K. The preferred and avoidance temperatures of Cyclops vicinus (Uljanin, 1875) and their relation to the optimal, pessimal and temperature of normal functioning of species. J. Plankton Res. 2017, 39, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Cowles, R.B.; Bogert, C.M. A preliminary study of the thermal requirements of desert reptiles. Bull. Am. Mus. Nat. Hist. 1944, 83, 265–296. [Google Scholar]
- Lowe, C.H.; Vance, V.J. Acclimation of the critical thermal maximum of the reptile Urosaurus ornatus. Science 1955, 122, 73–74. [Google Scholar] [CrossRef]
- Hutchinson, V.H. Critical thermal maxima in salamanders. Physiol. Zool. 1961, 34, 92–125. [Google Scholar] [CrossRef]
- Cox, D.K. Effects of three heating rates on the critical thermal maximum of bluegill. In Thermal Ecology, AEC Symposium Series; Gibbons, J.W., Sharitz, R.R., Eds.; National Technical Information Service: Springfield, MO, USA, 1974; pp. 158–163. [Google Scholar]
- Becker, C.D.; Genoway, R.G. Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environ. Biol. Fishes 1979, 4, 245–256. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: Data to support the onset of spasms as the definitive end point. Can. J. Zool. 1997, 75, 1553–1560. [Google Scholar] [CrossRef]
- Verbitsky, V.B.; Grishanin, A.K.; Malysheva, O.A.; Medyantseva, E.N.; Verbitskaya, T.I. Thermal resistance, preferred and avoidance temperatures of Cyclops strenuus Fisher, 1851, and their relation to optimal, pessimal and tolerant temperatures. Biol. Bull. Russ. 2017, 44, 439–448. [Google Scholar] [CrossRef]
- Reutter, J.M.; Herdendorf, C.E. Laboratory estimates of the seasonal final temperature preferenda of some Lake Erie fish. In Proceedings of the 17th Conference on Great Lakes Research; International Association for Great Lakes Researsh, Haqmilton, ON, Canada, 12–14 August, 1974; Rukavina, N.A., Seddon, J.S., Eds.; Braun-Brufield, Inc.: Jackson, MI, USA, 1974; Part 1; pp. 59–67. [Google Scholar]
- Diaz, H.F.; Espina, S.; Buckle, L.F. Thermal stress responses of Procambarus clarkia. Riv. Ital. Acquacolt. 1994, 29, 149–154. [Google Scholar]
- Hernandez, R.M.; Bückle, R.L.F.; Diaz, H.F. Preferred temperature of Macrobrachium tenellum (Crustacea, Palaemonidae). Riv. Ital. Acquacolt. 1995, 30, 93–96. [Google Scholar]
- Johansen, P.H.; Cross, J.A. Elfects of sexual maturation and sex steroid hormone treatment on the temperature preference of the guppy, Poecilia reticulata (Peters). Can. J. Zool. 1980, 58, 586–588. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Kelsch, S.W. Temperature selection and performance by blue- gills: Evidence for selection in response to available power. Trans. Am. Fish. Soc. 1996, 112, 424–430. [Google Scholar] [CrossRef]
- R Develpment Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: http://www.R-project.org/ (accessed on 4 April 2021).
- Einsle, U. Crustacea, Copepoda: Calanoida und Cyclopoida; Schwoerbel, J., Zwick, P., Eds.; Süsswasserfauna von Mitteleuropa, 8; Gustav Fischer Verlag: Stuttgart, Germany, 1993; pp. 120–142. [Google Scholar]
- Maurer, V.; Zeh, M. Crustaceen Plankton der Grossen Berner Seen; HYDRA-Institut, Gewässer- und Bodenschutzlabor des Kantons: Bern, Switzerland, 1999. [Google Scholar]
- Stanković, I.; Ternjej, I. The first record of Cyclops bohater Kozminski (Copepoda, Cyclopoida) in Croatia and the Balkan peninsula. Nat. Croat. 2007, 16, 189–199. [Google Scholar]
- Alekseev, V.R.; Sukhikh, N.M. Molecular genetic characteristics of the deepwater Cyclops bohater Kozminski, 1933 (Cyclopiformes, Copepoda) population from Lake Glubokoe (Moscow Region) and place of the species in the genus system. In Hydrobiological Station on the Glubokoe Lake: Proceedings; Korovchinsky, N.M., Ed.; KMK Scientific Press Ltd.: Moscow, Russia, 2022; Volume 12, pp. 92–104. (In Russian) [Google Scholar]
- Chown, S.L.; Terblanche, J.S. Physiological diversity in insects: Ecological and evolutionary contexts. Adv. Insect Physiol. 2007, 33, 50–152. [Google Scholar] [CrossRef]
- Rohr, J.R.; Civitello, D.J.; Cohen, J.M.; Roznik, E.A.; Sinervo, B.; Dell, A.I. The complex drivers of thermal acclimation and breadth in ectotherms. Ecol. Lett. 2018, 21, 1425–1439. [Google Scholar] [CrossRef] [Green Version]
- Verbitsky, V.B.; Grishanin, A.K.; Zhdanova, S.M.; Lazareva, V.I.; Malysheva, O.A.; Medyantseva, E.N. Temperature reactions in twelve species of freshwater Cyclopoida copepods. Zool. Zhurnal 2016, 95, 815–825. [Google Scholar] [CrossRef]
- Gunderson, A.R.; Stillman, J.H. Plasticity in thermal tolerance has limited potential to buffer ectotherms from global warming. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282, 20150401. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
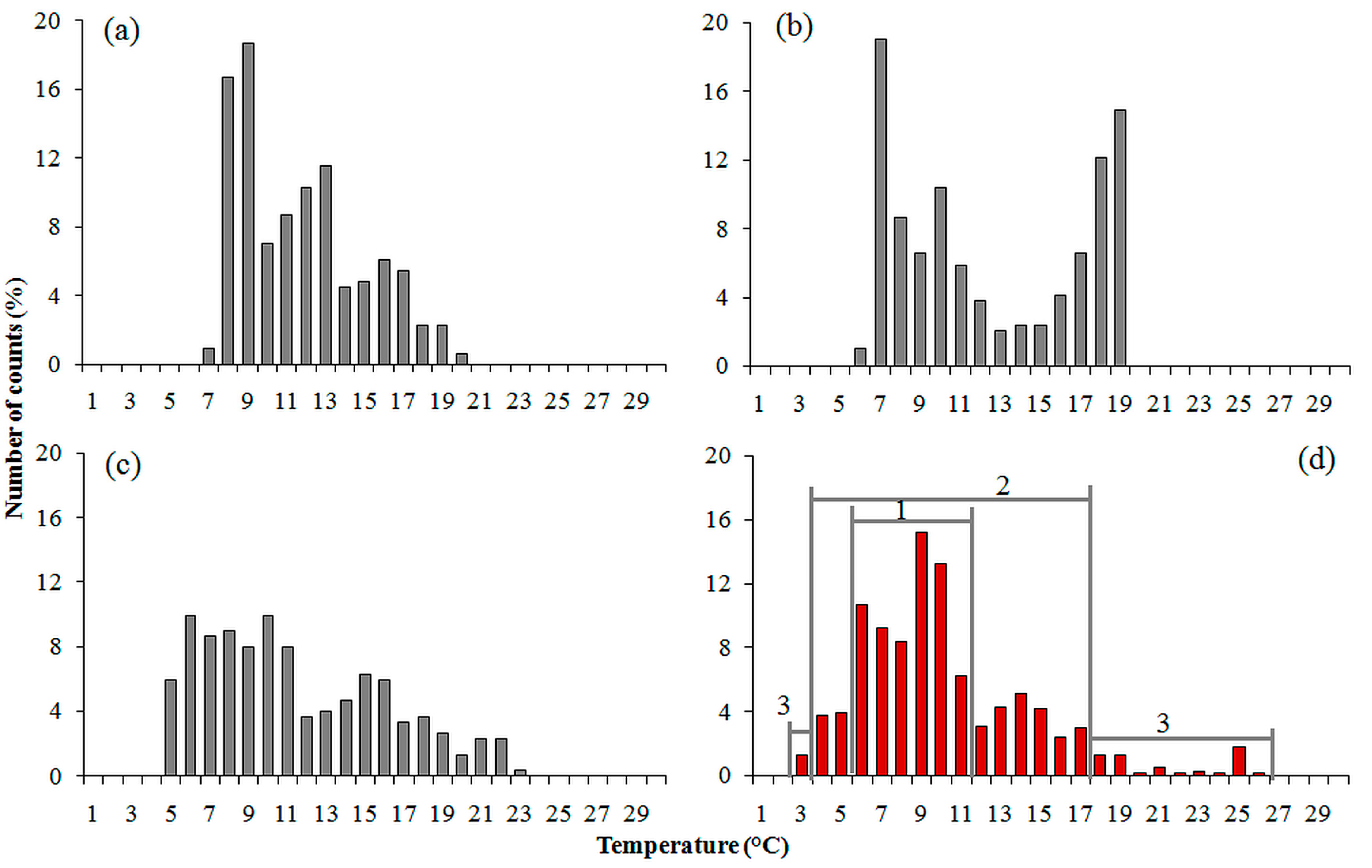{kind=link}
| Monogram | Toponym | Latitude (N) | Longitude (IE) | Area (km2) | Maximum Depth (cm) | Observation Times | Water Temperature (°C) |
|---|---|---|---|---|---|---|---|
| RR | Rybinsk Reservoir | 59°05′58″ | 38°27′30″ | 4550 | 2100 | December March | 0.3–1.7 0.7–3.4 |
| LF | Lake Ferapontovskoe | 59°57′09″ | 38°34′07″ | 1.5 | 3200 | February July | 1.1–1.5 8–11 |
| LG | Lake Glubokoe | 55°45′11″ | 36°30′18″ | 0.6 | 2700 | June–July | 5–9 |
| Waterbody | Latitude (N), Longitude (IE) | The Period of Maximum Reproduction | T, °C for Eggs Development | Authors |
|---|---|---|---|---|
| Rybinsk Reservoir | 59°05′58″, 38°27′30″ | March | 0.7–3.4 | present study |
| Lake Ferapontovskoe | 59°57′09″, 38°34′07″ | July | 8–11 * | present study |
| July | 8–12 * | [26] | ||
| February–March | 1.1–5.0 | [26] | ||
| Lake Glubokoe | 55°45′11″, 36°30′18″ | June–July | 5–9 * | present study |
| Lake Wigry | 54°02′12″, 23°05′54″ | February | – | [20] |
| Lake Marien | 53°01′11″, 14°16′44″ | November–February | – | [25] |
| Lake Constance, Lake Zurich | 47°36′06″, 09°27′09″ 47°14′51″, 08°40′39″ | January–February January–February | – – | [53] [53] |
| June–July | – | [20] | ||
| Lake Biel | 47°04′51″, 07°09′40″ | November–February | – | [54] |
| Lake Čingi-Lingi | 46°08′11″, 17°03′11″ | August–September | – | [55] |
| Time, Days | PTm (°C) | Median of PT (°C) | Lowe Quartile of PT | Upper Quartile of PT | Range of PT (°C) | Readings in Range of PT (%) | TNP (°C) | Readings in Range of TNP (%) | W-Criterion | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 10.2 ± 1.8 | 10 | 9 | 12 | 8–13 | 73 | 8–17 | 94 | 0.919 | <0.001 |
| 2 | 7.9 ± 1.9 | 7 | 6 | 9 | 6–12 | 70 | 6–20 | 94 | 0.833 | <0.001 |
| 3 | 9.9 ± 1.8 | 10 | 9 | 11 | 7–14 | 77 | 7–20 | 99 | 0.868 | <0.001 |
| 4 | 8.5 ± 1.6 18.6 ± 0.5 | 8 19 | 7 18 | 10 19 | 7–12 18–19 | 64 23 | 7–19 | 100 | 0.822 | <0.001 |
| 5 | 8.8 ± 1.7 17.9 ± 1.0 | 8 18 | 7 17 | 10 19 | 7–12 16–19 | 54 38 | 7–19 | 99 | 0.890 | <0.001 |
| 6 | 8.8 ± 2.4 16.6 ± 1.2 | 8 17 | 7 15 | 11 18 | 6–13 15–18 | 59 23 | 6–19 | 98 | 0.939 | <0.001 |
| 7 | 9.7 ± 2.7 | 9 | 7 | 11 | 6–15 | 82 | 5–18 | 96 | 0.940 | <0.001 |
| 8 | 9.9 ± 3.3 | 10 | 7 | 12 | 5–16 | 84 | 5–19 | 94 | 0.940 | <0.001 |
| 9 | 10.2 ± 2.9 | 10 | 8 | 13 | 6–15 | 78 | 6–19 | 95 | 0.954 | <0.001 |
| 10 | 8.0 ± 1.6 a | 9 | 7 | 9 | 5–10 | 63 | 5–17 | 92 | 0.916 | <0.001 |
| 11 | 7.3 ± 2.5 a | 8 | 5 | 9 | 3–11 | 72 | 3–17 | 99 | 0.964 | <0.001 |
| 12 | 8.9 ± 1.6 a | 9 | 7 | 10 | 6–11 | 63 | 6–17 | 92 | 0.959 | <0.001 |
| 13 | 8.4 ± 1.8 a | 9 | 7 | 10 | 6–11 | 68 | 6–19 | 93 | 0.874 | <0.001 |
| 14 | 8.0 ± 1.5 a | 8 | 7 | 10 | 6–10 | 66 | 5–17 | 92 | 0.798 | <0.001 |
| 15 | 7.3 ± 2.0 a | 8 | 6 | 9 | 4–10 | 77 | 4–16 | 94 | 0.894 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazareva, V.; Mayor, T.; Malysheva, O.; Medyantseva, E.; Zhdanova, S.; Grishanin, A.; Verbitsky, V. Thermal Tolerance of Cyclops bohater (Crustacea: Copepoda); Selection of Optimal and Avoided Conditions in Experimental Conditions. Diversity 2022, 14, 1106. https://doi.org/10.3390/d14121106
Lazareva V, Mayor T, Malysheva O, Medyantseva E, Zhdanova S, Grishanin A, Verbitsky V. Thermal Tolerance of Cyclops bohater (Crustacea: Copepoda); Selection of Optimal and Avoided Conditions in Experimental Conditions. Diversity. 2022; 14(12):1106. https://doi.org/10.3390/d14121106
Chicago/Turabian StyleLazareva, Valentina, Tatyana Mayor, Olga Malysheva, Elena Medyantseva, Svetlana Zhdanova, Andrey Grishanin, and Vladimir Verbitsky. 2022. "Thermal Tolerance of Cyclops bohater (Crustacea: Copepoda); Selection of Optimal and Avoided Conditions in Experimental Conditions" Diversity 14, no. 12: 1106. https://doi.org/10.3390/d14121106
APA StyleLazareva, V., Mayor, T., Malysheva, O., Medyantseva, E., Zhdanova, S., Grishanin, A., & Verbitsky, V. (2022). Thermal Tolerance of Cyclops bohater (Crustacea: Copepoda); Selection of Optimal and Avoided Conditions in Experimental Conditions. Diversity, 14(12), 1106. https://doi.org/10.3390/d14121106