
A New Species of the Genus Robertgurneya Apostolov & Marinov, 1988 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Jeju Island, Korea



Abstract
:1. Introduction
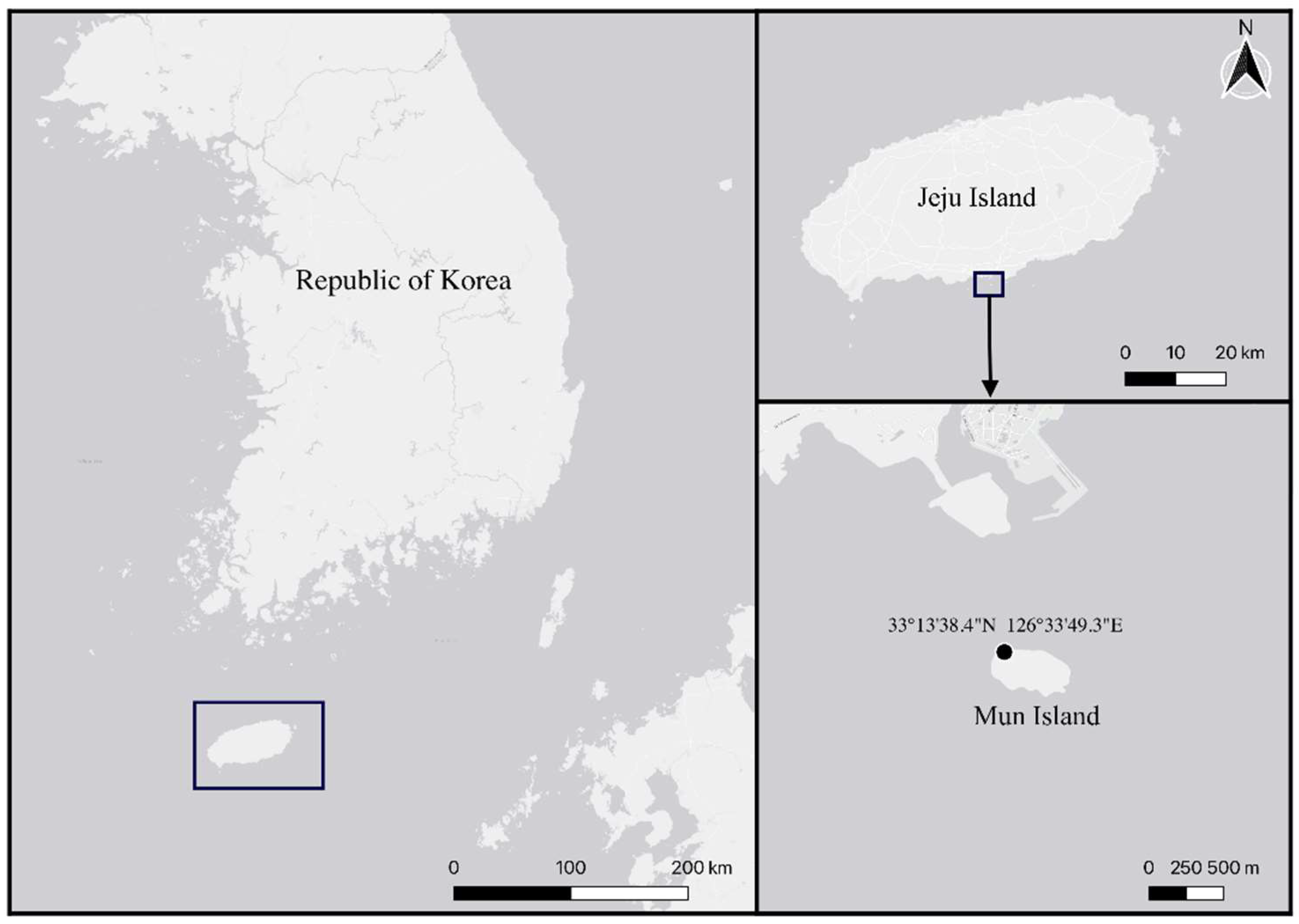
2. Materials and Methods
2.1. Specimen Collection and Observation
2.2. DNA Extraction and Amplification
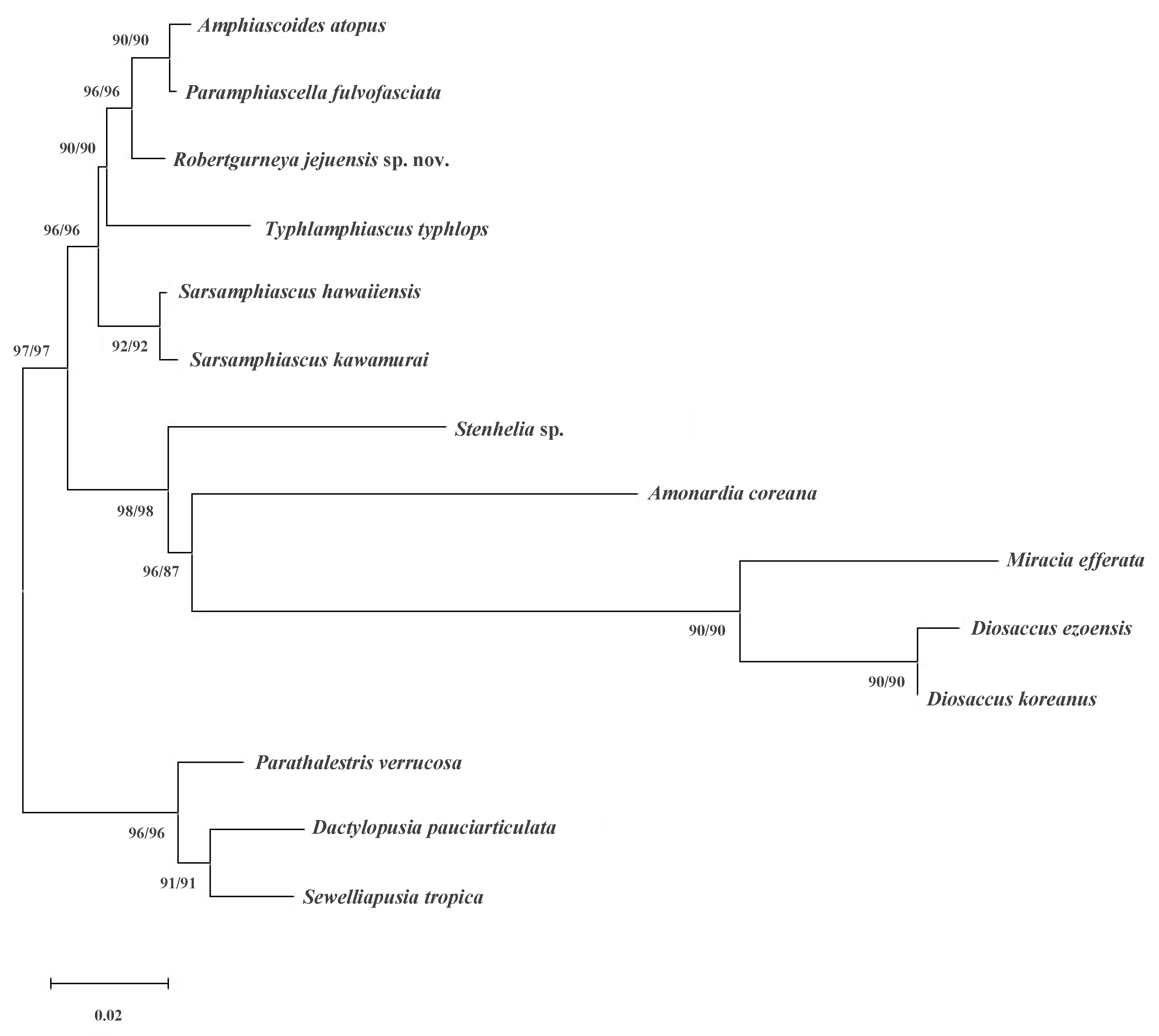
2.3. Phylogenetic Analyses
3. Results
3.1. Systematics
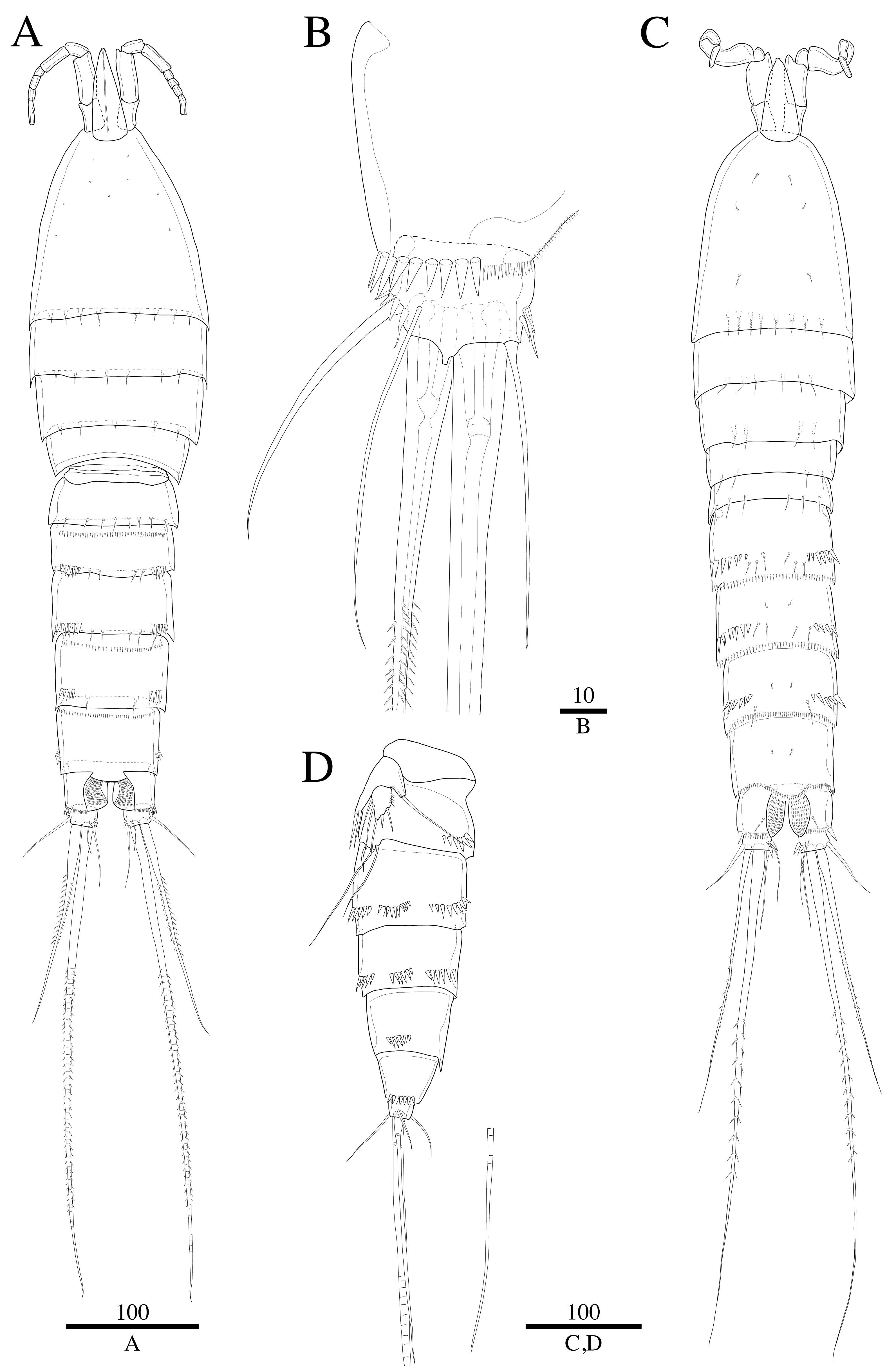
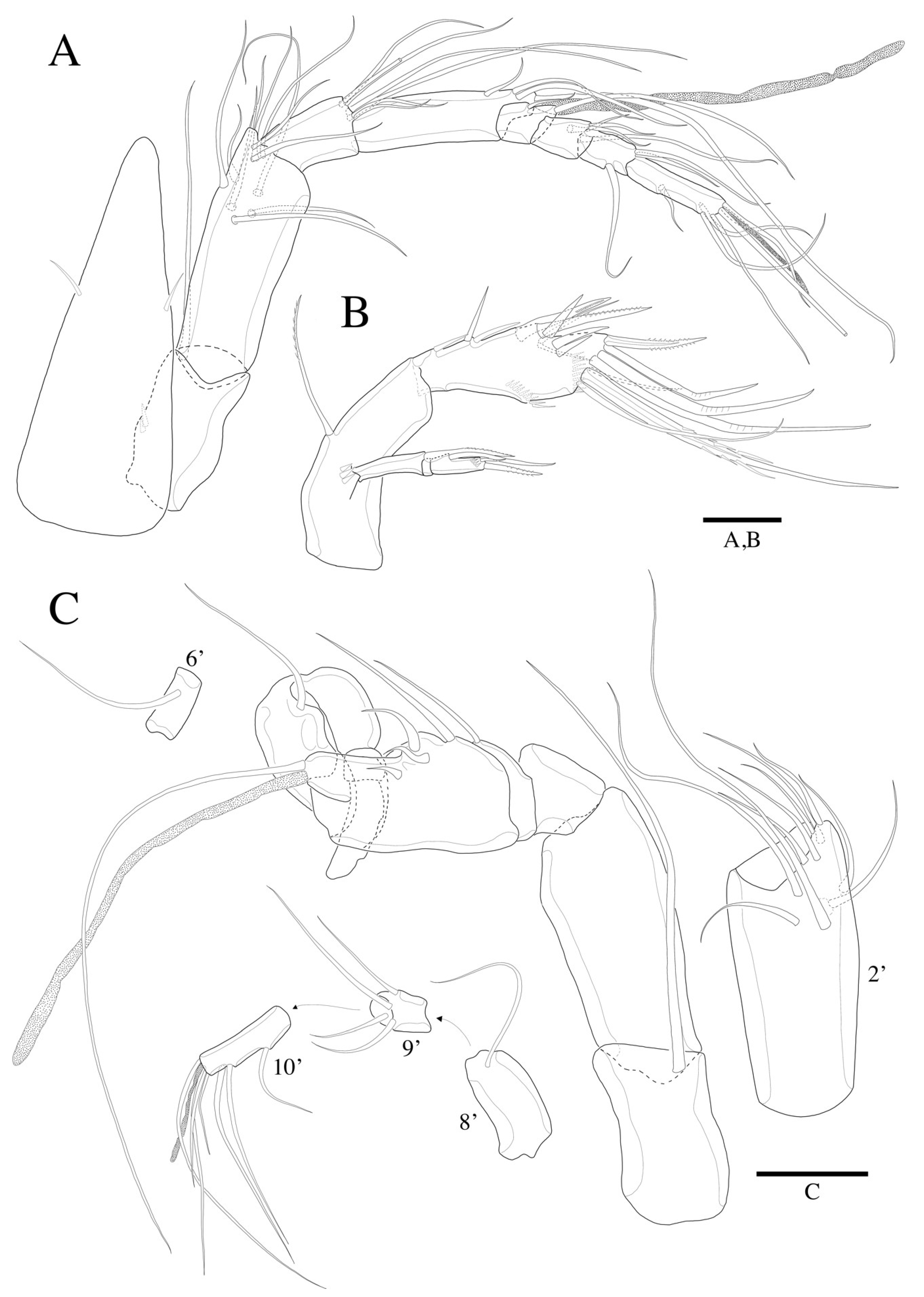
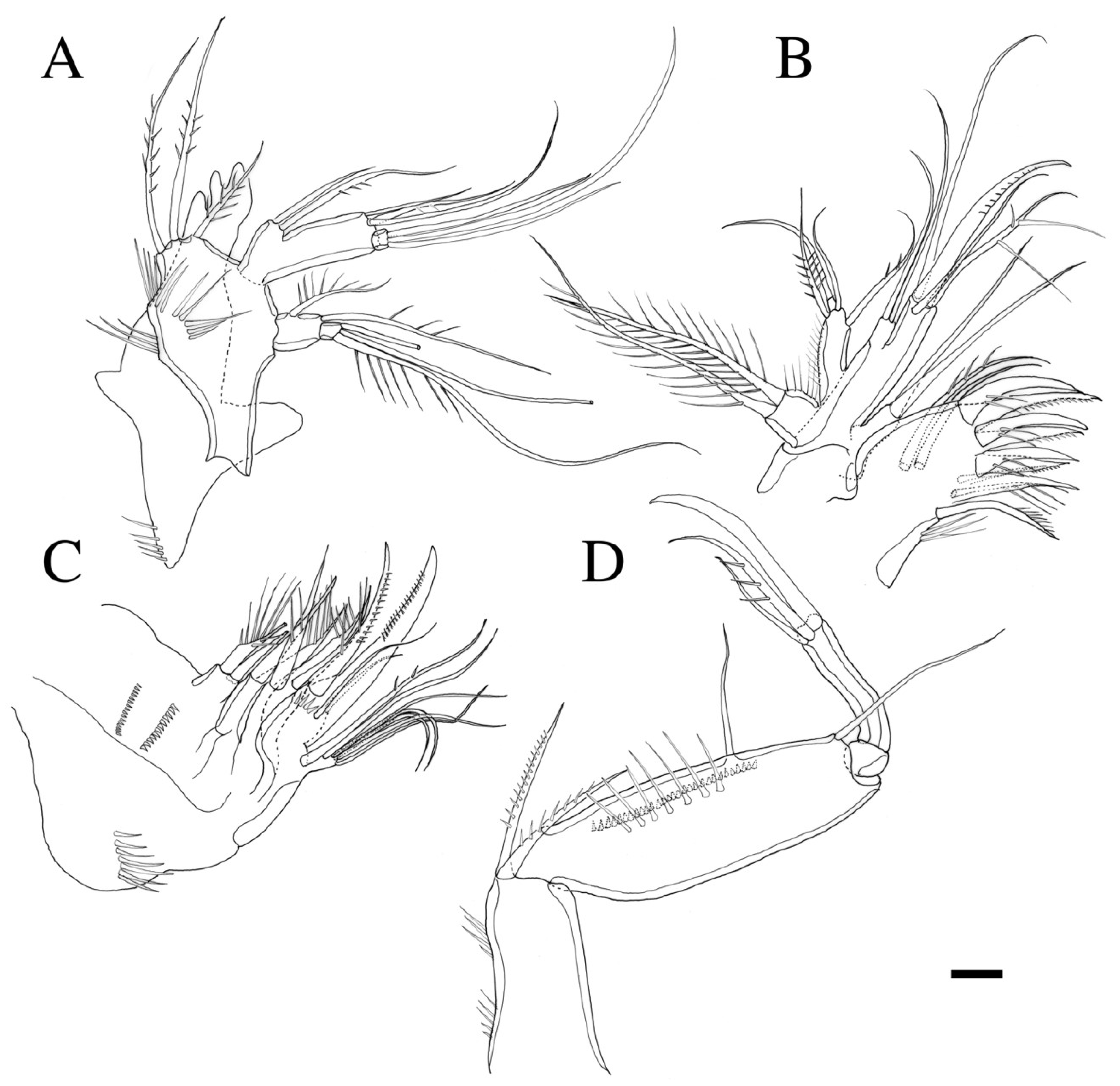
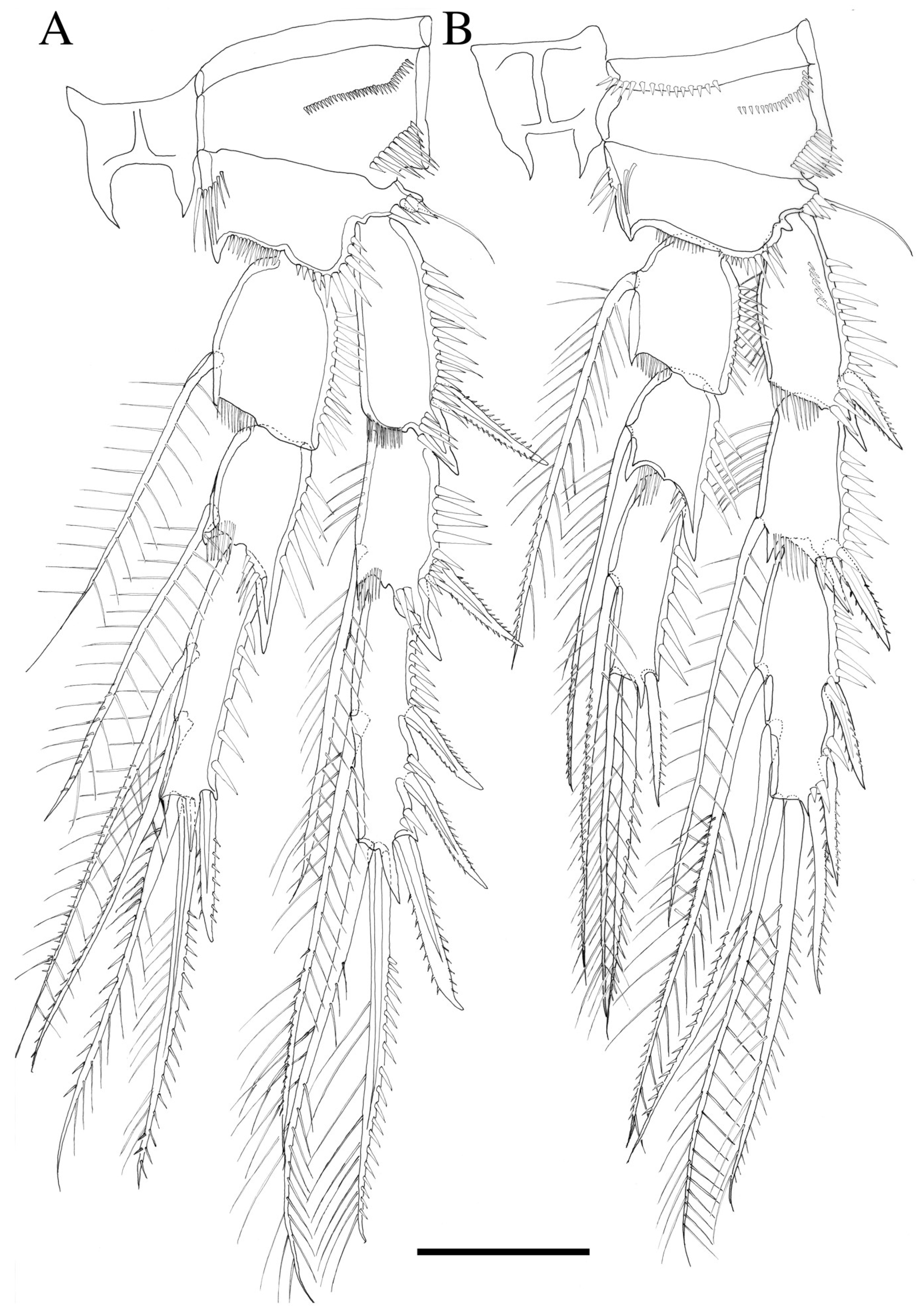
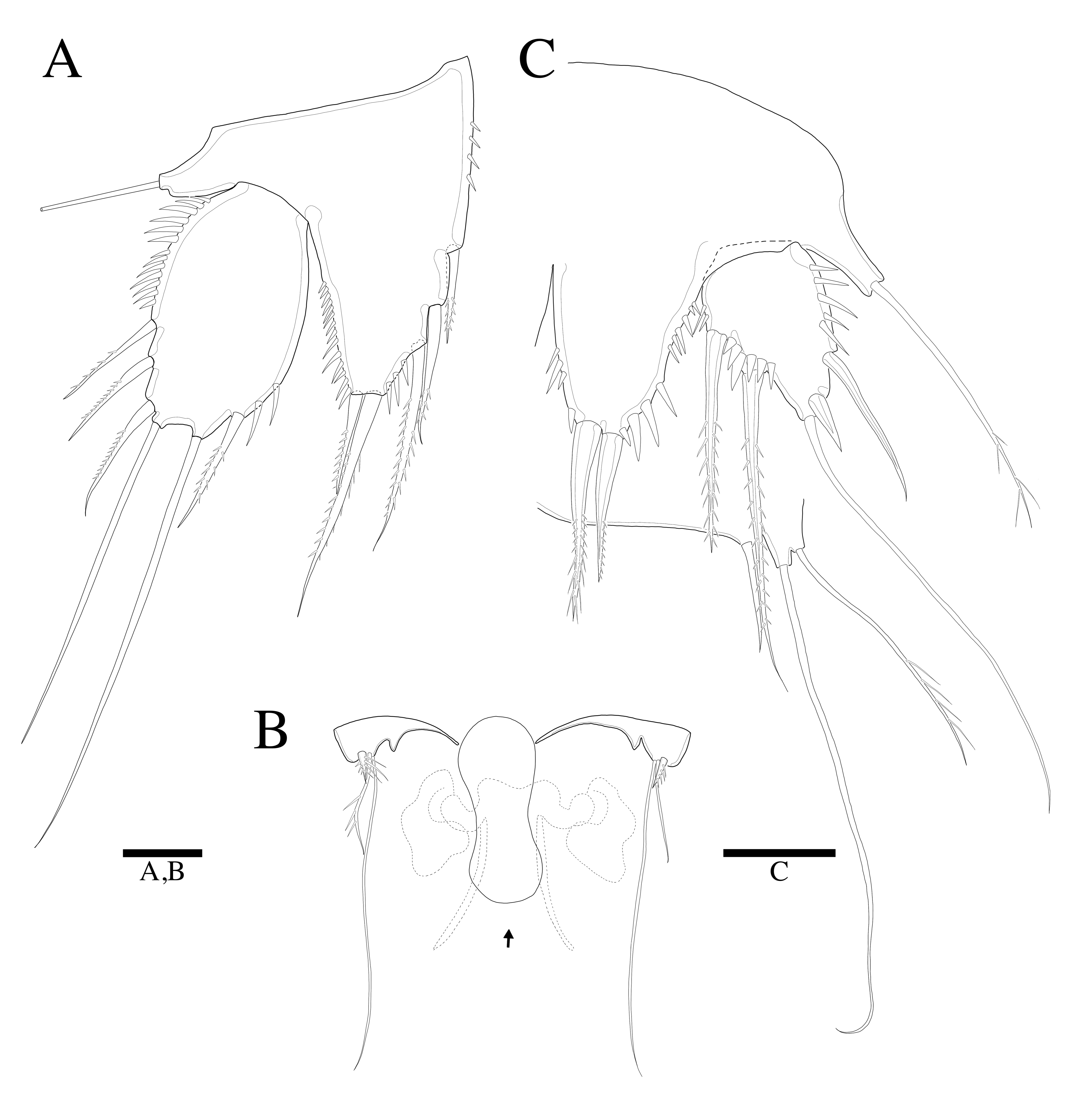
3.2. Description
3.2.1. Description of Female
3.2.2. Description of Male
4. Discussion
- P1 enp-2 without inner seta …………………………………………………................R. remaneiP1 enp-2 with inner seta ……………………………………………………………....…........... 2
- P2 enp-1 without inner seta; P3 enp-3 with one inner seta; female P5 setae benp: exp = 4:5 ……………………………………………………………………………....................... R. brevipesP2 enp-1 with inner seta; P3 enp-3 with two inner setae; female P5 setae benp: exp = 5:6…………………………………………………………………………………………............... 3
- P5 exp l/b = 1.5 …………………………………………………………………………..… ..........4P5 exp l/b > 1.5 ………………………………………………………………………..….............. 7
- P1 enp-1/exp = 1; P1 enp-1 inner seta reaching 2/3 of enp-3……………….......... R. hopkinsiP1 enp-1 > exp; P1 enp-1 inner seta reaching the middle of enp-3 ……………………....... 5
- P5 exp distal margin swollen between setae II and III in both sexes …………...... R. smithiP5 exp distal margin normal, oval-shaped ………………………………………..….…........ 6
- P1 basis inner spine of male reaching tip of P1 exp ……………………....... R. falklandiensisP1 basis inner spine of male reaching P1 exp-2 …………………………….…........ R. diversa
- Male P5 exp with six setae ……………………………………………………......... R. simulansMale P5 exp with five setae …………………………………………………………..….......... 8
- Female P5 exp l/b = 2; P5 benp without ornamentation between seta II and III …........... 9Female P5 exp l/b < 2; P5 benp with ornamentation between seta II and III ……….…………………………………………………………….…....................... R. jejuensis sp. nov.
- P1 exp/enp-1 = 2/3; female P5 exp distal margin swollen………...…...…...... R. donghaensisP1 exp/enp-1 = 4/5; female P5 exp distal margin normal, oval-shaped ……….……........ 10
- A1 first/second segment length ratio = 0.38; female P5 exp outer setae short and bulbous; P5 benp distal setae length ratio I/II = 1/4...…………….............… R. similis bulbamphiascoidesA1 first/second segment length ratio = 1; female P5 exp outer setae slender and pinnate; P5 benign distal setae length ratio I/II = 1 …………………………………....... R. similis similis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boxshall, G.A.; Halsey, S.H. An Introduction to Copepod Diversity; Ray Society: London, UK, 2004. [Google Scholar]
- Willen, E. Notes on the Systematic Position of the Stenheliinae (Copepoda, Harpacticoida) Within the Thalestridimorpha and Description of Two New Species from Motupore Island, Papua New Guinea. Cah. Biol. Mar. 2002, 43, 27–42. [Google Scholar]
- Song, S.J.; Yun, S.G.; Chang, C.Y. New Records on Three Harpacticoid Copepods Associated with Marine Macroalgae in Korea. Fish. Aquat. Sci. 1999, 2, 189–198. [Google Scholar]
- Song, S.J.; Rho, H.S.; Kim, W. A New Species of Amonardia (Copepoda: Harpacticoida: Miraciidae) from the Cultivated Brown Alga, Undaria pinnatifida. Integr. Biosci. 2007, 11, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y. Three Miraciid Copepods (Harpacticoida, Miraciidae) from South Korea. Anim. Syst. Evol. Divers. 2009, 25, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Nam, E.; Lee, W. First Record of the Genus Sinamphiascus (Copepoda: Harpacticoida) from Korean Waters. J. Species Res. 2012, 1, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Karanovic, T.; Kim, K. New Insights into Polyphyly of the Harpacticoid Genus Delavalia (Crustacea, Copepoda) Through Morphological and Molecular Study of an Unprecedented Diversity of Sympatric Species in a Small South Korean Bay. Zootaxa 2014, 3783, 1–96. [Google Scholar] [CrossRef]
- Karanovic, T. Cladistic and Quantitative Shape Analyses of Five New Syntopic Sarsamphiascus (Copepoda, Harpacticoida): Problems and Solutions for Diosaccin Systematics and Taxonomy. Syst. Biodivers. 2020, 18, 810–833. [Google Scholar] [CrossRef]
- Bang, H.W. A New Species of the Genus Robertgurneya (Copepoda: Harpacticoida: Miraciidae) from the East Sea of Korea. Korean J. Environ. Biol. 2021, 39, 590–603. [Google Scholar] [CrossRef]
- Apostolov, A.M.; Marinov, T.M. Copepoda, Harpacticoida (morski kharpaktikoidi). Izd Vo Na Bŭlgarskata Akad. a Na Nauk. 1988, 18, 1–384. [Google Scholar]
- Lang, K. Monographie der Harpacticiden (Vorläufige Mitteilung); Almqvist & Wiksellls Boktryckeri: Upssala, Sweden, 1944. [Google Scholar]
- Lang, K. Monographie der Harpacticiden; Håkan Ohlsson: Lund, Sweden, 1948; Volume 2, pp. 1–1682. [Google Scholar]
- Huys, R. Unresolved Cases of Type Fixation, Synonymy and Homonymy in Harpacticoid Copepod Nomenclature (Crustacea: Copepoda). Zootaxa 2009, 2183, 1–99. [Google Scholar] [CrossRef]
- Gómez, S. A New Species of Robertgurneya Apostolov Marinov, 1988, with an Illustrated Record of R. rostrata (Gurney, 1927), an Amended Genus Diagnosis and Comments on R. Soyeri (Apostolov, 1974) and R. spinulosa (Sars, 1911) (Harpacticoida: Miraciidae). Zootaxa 2020, 4861, 4861. [Google Scholar] [CrossRef]
- Huys, R.; Gee, J.M.; Moore, C.G.; Hamond, R. Marine and Brackish Water Harpacticoid Copepods. Published for the Linnean Society of London and the Estuarine and Coastal Sciences Association by Field Studies Council. Sypopses Br. Fauna (New Ser.) London U.K. 1996, 51, 1–352. [Google Scholar]
- Wells, J.B.J. An Annotated Checklist and Keys to the Species of Copepoda Harpacticoida (Crustacea). Zootaxa 2007, 1568, 1–872. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.D.; Schrank, B.; Huynh, C.; Shownkeen, R.; Waterston, R.H. A Genetic Mapping System in Caenorhabditis elegans Based on Polymorphic Sequence-Tagged Sites. Genetics 1992, 131, 609–624. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Endo, K. Molecular Phylogeny of Ostracoda (Crustacea) Inferred from 18S Ribosomal DNA Sequences: Implication for Its Origin and Diversification. Mar. Biol. 2003, 143, 23–38. [Google Scholar] [CrossRef]
- Bucklin, A.; Ortman, B.D.; Jennings, R.M.; Nigro, L.M.; Sweetman, C.J.; Copley, N.J.; Sutton, T.; Wiebe, P.H.A. “Rosetta Stone” for Metazoan Zooplankton: DNA Barcode Analysis of Species Diversity of the Sargasso Sea (Northwest Atlantic Ocean). Deep Sea Res. II 2010, 57, 2234–2247. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Huys, R.; Mackenzie-Dodds, J.; Llewellyn-Hughes, J. Cancrincolidae (Copepoda, Harpacticoida) Associated with Land Crabs: A Semiterrestrial Leaf of the Ameirid Tree. Mol. Phylogenet. Evol. 2009, 51, 143–156. [Google Scholar] [CrossRef]
- Yeom, J.; Lee, W. A New Species of the Genus Sarsamphiascus Huys, 2009 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Hawaii. PeerJ 2020, 8, e8506. [Google Scholar] [CrossRef] [Green Version]
- Lim, B.J.; Bang, H.W.; Moon, H.; Back, J. Integrative Description of Diosaccus koreanus sp. nov.(Hexanauplia, Harpacticoida, Miraciidae) and Integrative Information on Further Korean Species. ZooKeys 2020, 927, 1–35. [Google Scholar] [CrossRef]
- Wells, J.B.J. Keys to Aid in the Identification of Marine Harpacticoid Copepods; Department of Zoology, University of Aberdeen: Aberdeen, Scotland, 1976; pp. 1–215. [Google Scholar]
- Lang, K. Copepoda Harpacticoida from the Californian Pacific Coast. K. Sven. Vetensk. Handl. 1965, 10, 1–560. [Google Scholar]
- Gómez, S.; Corgosinho, P.H.C.; Rivera-Sánchez, K.I. Proposal of New Genera and Species of the Subfamily Diosaccinae (Copepoda: Harpacticoida: Miraciidae). Eur. J. Taxon. 2021, 759, 1–62. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Author, Year | NCBI No. | Authors |
|---|---|---|---|---|---|
| Miraciidae Dana, 1846 | Amphiascoides | atopus | Lotufo and Fleeger, 1995 | KC815328 | Gómez et al., 2013, unpublished data |
| Paramphiascella | fulvofasciata | Rosenfield and Coull, 1974 | EU380293 | Huys et al., 2009 [22] | |
| Typhlamphiascus | typhlops | (Sars G.O., 1906) | EU380292 | Huys et al., 2009 [22] | |
| Sarsamphiascus | kawamurai | (Ueda and Nagai, 2005) | MN541394 | Yeom and Lee, 2020 [23] | |
| Sarsamphiascus | hawaiiensis | Yeom and Lee, 2020 | MN496456 | Yeom and Lee, 2020 [23] | |
| Stenhelia | sp. | EU380291 | Huys et al., 2009 [22] | ||
| Amonardia | coreana | Song, Rho, and Kim W., 2007 | KT030261 | Baek and Hwang, 2015, unpublished data | |
| Diosaccus | ezoensis | Itô, 1974 | KR048740 | Baek and Hwang, 2015, unpublished data | |
| Diosaccus | koreanus | Lim, Bang, Moon, and Back, 2020 | MT002900 | Lim et al., 2020 [24] | |
| Miracia | efferata | Dana, 1849 | EU380294 | Huys et al., 2009 [22] | |
| Robertgurneya | jejuensis sp. nov. | Yeom and Lee, 2022 | OP798781 | This study | |
| Dactylopusiidae Lang, 1936 | Dactylopusia | pauciarticulata | Chang and Song, 1997 | KR048735 | Baek and Hwang, 2015, unpublished data |
| Sewelliapusia | tropica | (Sewell, 1940) | EU380299 | Huys et al., 2009 [22] | |
| Thalestridae Sars G.O., 1905 | Parathalestris | verrucosa | Itô, 1970 | MT002906 | Lim et al., 2020 [24] |
| Setal Formulae | Exopod | Endopod |
|---|---|---|
| P1 | 0.0.022 | 1.1.111 |
| P2 | 0.1.123 | 1.2.121 |
| P3 | 0.1.123 | 1.1.221 |
| P4 | 0.1.223 | 1.1.121 |
| R. jejuensis sp. nov. | R. donghaensis | ||
|---|---|---|---|
| Female | Body length | 875 (n = 3) | 795 (n = 5) |
| Maxillule basis setae | 6 | 5 | |
| Maxilla allobasis and enp setae | 9 | 7 | |
| A1 1st/2nd seg length | 0.7 | 0.4 | |
| P1 enp-1 inner seta length | ~middle of enp-3 (long) | ~ enp-2 (short) | |
| P2 exp-2 inner margin ornamentation | present | absent | |
| P3 enp and exp length | enp < exp | enp ≥ exp | |
| P5 benp distal setae length | double | same | |
| P5 benp ornamentation between seta II and III | relatively wide spacing with ornamentation | without | |
| P5 exp shape (l/b ratio) | oval (2) | swollen distal margin (1.7) | |
| P5 exp outer setae type | pinnate | naked | |
| P6 mid seta (A), the longest seta (B) length | 3A = B | 2A = B | |
| CR ornamentation | inner margin | inner distal margin | |
| CR l/b ratio | 0.7 | 1 | |
| CR lateral setae type | naked | pinnate | |
| Male | Ornamentation on urosome segments surface | Setules on ventral, lateral, and dorsal surface | Setules on ventral surface |
| Ornamentation on A1 seg-1 | absent | present | |
| A1 seg-2 seta type | all naked | 1 pinnate | |
| A1 seg-5 distal surface | without element | 2 pinnate elements | |
| P2 enp-2 two inner setae type | naked | plumose | |
| P2 enp-2 outer margin ornamentation | absent | present | |
| P6 setae type | 3 naked | 1 pinnate and 2 naked |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeom, J.; Lee, W. A New Species of the Genus Robertgurneya Apostolov & Marinov, 1988 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Jeju Island, Korea. Diversity 2022, 14, 1127. https://doi.org/10.3390/d14121127
Yeom J, Lee W. A New Species of the Genus Robertgurneya Apostolov & Marinov, 1988 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Jeju Island, Korea. Diversity. 2022; 14(12):1127. https://doi.org/10.3390/d14121127
Chicago/Turabian StyleYeom, Jisu, and Wonchoel Lee. 2022. "A New Species of the Genus Robertgurneya Apostolov & Marinov, 1988 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Jeju Island, Korea" Diversity 14, no. 12: 1127. https://doi.org/10.3390/d14121127
APA StyleYeom, J., & Lee, W. (2022). A New Species of the Genus Robertgurneya Apostolov & Marinov, 1988 (Copepoda: Harpacticoida: Miraciidae) from a Sublittoral Zone of Jeju Island, Korea. Diversity, 14(12), 1127. https://doi.org/10.3390/d14121127