

Analysis of Prey Composition in Eurasian Reed Warblers’ Acrocephalus scirpaceus Droppings at Four Breeding Sites in Italy

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.1.1. Mirandola (Mortizzuolo)
2.1.2. Pergusa Lake
2.1.3. Gela Lake
2.1.4. Vendicari
2.2. Data Collection
2.3. Statistical Analysis
3. Results
3.1. Diet Composition
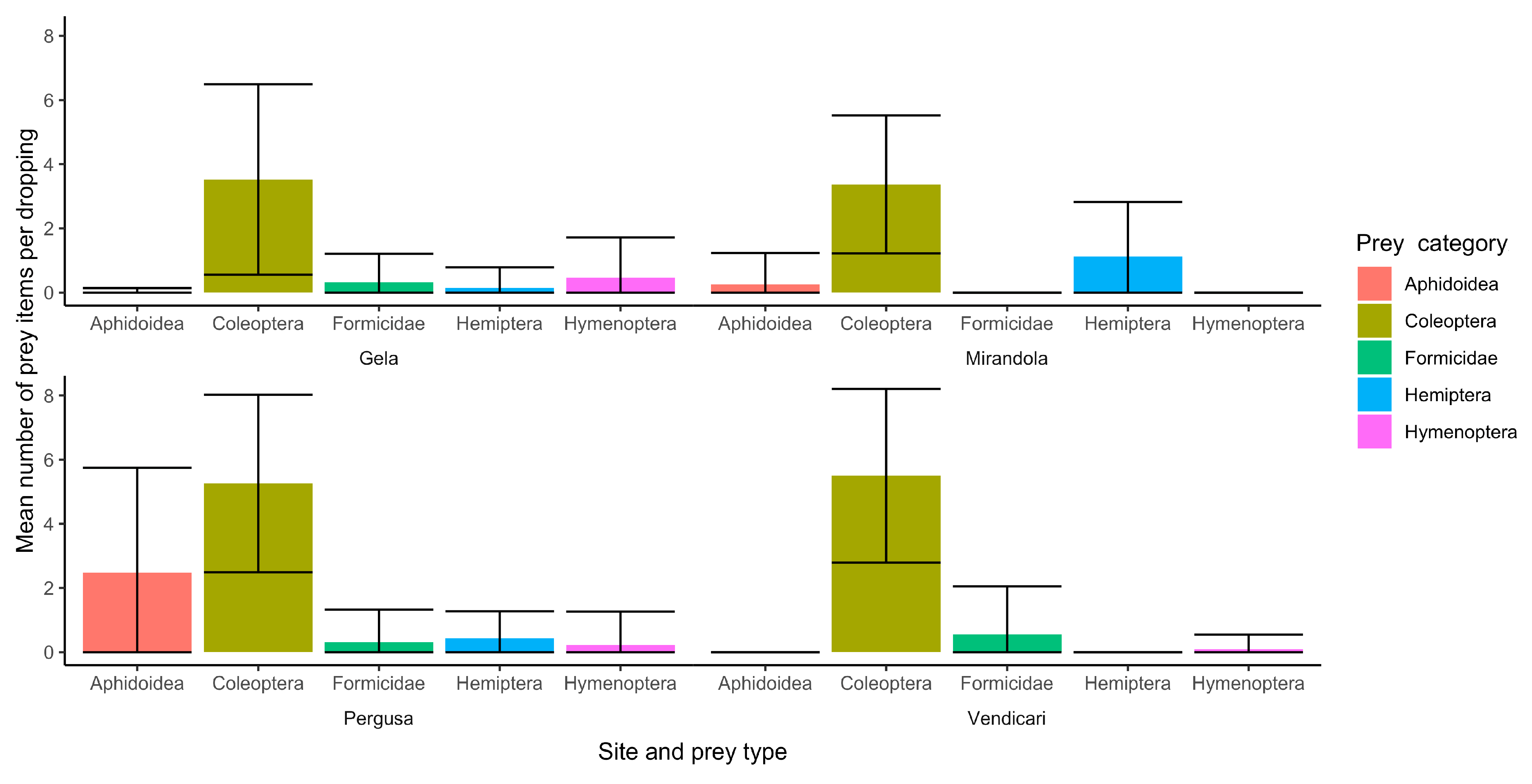
3.1.1. Mirandola
3.1.2. Pergusa Lake
3.1.3. Gela Lake
3.1.4. Vendicari
3.2. Diet Composition Analysis
3.3. Habitat Occurrences Versus Diet Composition
3.4. Prey Brought by the Reed Warbler to the Common Cuckoo
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Catchpole, C.K. Habitat selection and breeding success in the Reed Warbler (Acrocephalus scirpaceus). J. Anim. Ecol. 1974, 43, 363–380. [Google Scholar] [CrossRef]
- Leisler, B.; Schulze-Hagen, K. The Reed Warblers. Diversity in a Uniform Bird Family; KNNV: Zeist, The Netherlands, 2011. [Google Scholar]
- Hering, J.; Winkler, H.; Steinheimer, F.D. A new subspecies of Eurasian Reed Warbler Acrocephalus scirpaceus in Egypt. Bull. Br. Ornithol. Club 2016, 136, 101–128. [Google Scholar]
- Green, R.E.; Davies, N.B. Feeding ecology of Reed and Sedge Warblers. Wicken Fen Group Rep. 1972, 4, 8–14. [Google Scholar]
- Cramp, S. The Birds of the Western Palaearctic. Vol. VI. Warblers; Oxford University Press: Oxford, UK, 1992. [Google Scholar]
- Leisler, B.; Heidrich, P.; Schulze-Hagen, K.; Wink, M. Taxonomy and phylogeny of Reed Warblers (genus Acrocephalus) based on mtDNA sequences and morphology. J. Ornithol. 1997, 138, 469–496. [Google Scholar] [CrossRef]
- Procházka, P.; Stokke, B.G.; Jensen, H.; Fainová, D.; Bellinvia, E.; Fossøy, F.; Vilkan, J.R.; Bryja, J.; Soler, M. Low genetic differentiation among Reed Warbler Acrocephalus scirpaceus populations across Europe. J. Avian Biol. 2011, 42, 103–113. [Google Scholar] [CrossRef]
- Olsson, U.; Rguibi-Idrissi, H.; Copete, J.L.; Arroyo Matos, J.L.; Provost, P.; Amezian, M.; Alström, P.; Jiguet, F. Mitochondrial phylogeny of the Eurasian/African Reed Warbler complex (Acrocephalus, Aves). Disagreement between morphological and molecular evidence and cryptic divergence: A case for resurrecting Calamoherpe ambigua Brehm 1857. Mol. Phyl. Evol. 2016, 102, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Campobello, D.; Sealy, S.G. Evolutionary significance of antiparasite, antipredator and learning phenotypes of avian nest defence. Sci. Rep. 2018, 8, 10569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, M.; Ceraulo, M.; Tuliozi, B.; Buscaino, G.; Mazzola, S.; Sala, L.; Dal Zotto, M.; Campobello, D. Decoupled Acoustic and Visual Components in the Multimodal Signals of the Common Cuckoo (Cuculus canorus). Front. Ecol. Evol. 2021, 9, 725858. [Google Scholar] [CrossRef]
- Tolman, D.; Campobello, D.; Rönkä, K.; Kluen, E.; Thorogood, R. Reed Warbler hosts do not fine-tune mobbing defenses during the breeding season, even when cuckoos are rare. Front. Ecol. Evol. 2021, 9, 725467. [Google Scholar] [CrossRef]
- Birdlife IUCN. Available online: http://datazone.birdlife.org/userfiles/file/Species/erlob/supplementarypd-fs/22714722_acrocephalus_scirpaceus.pdf (accessed on 8 December 2022).
- Bellavance, V.; Bélisle, M.; Savage, J.; Pelletier, F.; Garant, D. Influence of agricultural intensification on prey availability and nestling diet in Tree Swallows (Tachycineta bicolor). Canad. J. Zool. 2018, 9, 1053–1065. [Google Scholar] [CrossRef] [Green Version]
- Chernetsov, N.; Manukyan, A. Feeding strategy of Reed Warblers Acrocephalus scirpaceus on migration. Avian Ecol. Behav. 1999, 3, 59–68. [Google Scholar]
- Kazlauskas, R.; Pukas, A.; Meldazyte, R. On Feeding of Warblers (Acrocephalus) in Reproductive Period, Western Lithuania. In Ecology of Birds of Lithuanian SSR. Part 3. Human Impact on the Avifauna and Its Conservation; Valius, M., Ed.; Vilnius, Lithuania, 1986; pp. 130–149. [Google Scholar]
- Dyrcz, A. Die Nestlingsuchrung bei Drosselrohrsänger Acrocephalus arundinaceus und Teichrohrsänger Acrocephalus scirpaceus au den Teichen bei Miliez in Polen und zwei Seen in der Westschweiz. Die Ornithol. Beob. 1979, 76, 305–316. [Google Scholar]
- Bibby, C.J.; Thomas, D.K. Breeding and diets of the Reed Warbler at a rich and a poor site. Bird Study 1985, 32, 19–31. [Google Scholar] [CrossRef]
- Grim, T.; Honza, M. Effect of habitat on the diet of Reed Warbler (Acrocephalus scirpaceus) nestlings. Folia Zool. 1996, 45, 31–34. [Google Scholar]
- Henry, C. Le nourrissage des jeunes chez la Rousserolle effarvette (Acrocephalus scirpaceus). Description du régime et effort de chasse des parents. Le Gerfaut 1977, 67, 369–394. [Google Scholar]
- Henry, C. Le concept de niche écologique illustré par les cas des populations congénériques sympatriques du genre Acrocephalus. Terre et Vie (Rev. Ecol.) 1979, 33, 457–492. [Google Scholar] [CrossRef]
- Bussmann, C. Ökologische Sonderung der Rohrsänger Südfrankreichs aufgrund von Nahrungsstudien. Vogelwarte 1979, 30, 84–101. [Google Scholar]
- Bibby, C.J.; Green, R.E. Autumn migration strategies of Reed and Sedge Warblers. Ornis Scand. 1981, 12, 1–12. [Google Scholar] [CrossRef]
- Kerbiriou, C.; Bargain, B.; Le Viol, I.; Pavoine, S. Diet and fuelling of the globally threatened aquatic warbler at autumn migration stopover as compared with two congeners. Anim. Cons. 2011, 14, 261–270. [Google Scholar] [CrossRef]
- Rguibi Idrissi, H.; Lefebvre, G.; Poulin, B. Diet of Reed Warblers Acrocephalus scirpaceus at two stopover sites in Morocco during autumn migration. Rev. Ecol. (Terre Vie) 2004, 59, 491–502. [Google Scholar]
- Poulin, B.; Lefebvre, G. Additional information on the use of tartar emetic in determining the diet of tropical birds. Condor 1995, 97, 897–902. [Google Scholar] [CrossRef]
- Davies, N.B. Food, flocking and territorial behaviour of the Pied Wagtail Molacilla alba yarellii Gould in winter. J. Anim. Ecol. 1976, 45, 235–254. [Google Scholar] [CrossRef]
- Davies, N.B. Prey selection and social behaviour in wagtails (Aves: Motacillidae). J. Anim. Ecol. 1977, 46, 37–57. [Google Scholar] [CrossRef]
- Wolda, H. Food Availability for an Insectivore and How to Measure It. In Avian Foraging: Theory, Methodology, and Applications. Studies in Avian Biology; Morrison, M.L., Ralph, C.J., Verner, J., Jehl, J.R., Jr., Eds.; Cooper Ornithological Society: San Diego, CA, USA, 1990; pp. 38–43. [Google Scholar]
- Halsch, C.A.; Shapiro, A.M.; Fordyce, J.A.; Nice, C.C.; Thorne, J.H.; Waetjen, D.P.; Forister, M.L. Insects and recent climate change. Proc. Natl. Acad. Sci. USA 2021, 118, e2002543117. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Hobson, K. Climate change: Aerial insectivores struggle to keep pace with earlier pulses of nutritious acquatic foods. Curr. Biol. 2022, 32, R267–R269. [Google Scholar] [CrossRef]
- Climate ADAPT. Available online: https://climate-adapt.eea.europa.eu/en/countries-regions/countries/italy (accessed on 1 November 2022).
- Halupka, L.; Borowiec, M.; Neubauer, G.; Halupka, K. Fitness consequences of longer breeding seasons of a migratory passerine under changing climatic conditions. J. Anim. Ecol. 2021, 90, 1655–1665. [Google Scholar] [CrossRef]
- Campobello, D.; Sealy, S.G. Avian brood parasitism in a Mediterranean region: Hosts and habitat preferences of Common Cuckoos Cuculus canorus. Bird Study 2009, 56, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Campobello, D.; Sealy, S.G. Avian brood parasitism in Italy: Another perspective. Avocetta 2020, 44, 21–27. [Google Scholar]
- Davies, N.B.; Brooke, M.D.L. An experimental study of co-evolution between the cuckoo, Cuculus canorus, and its hosts. I. Host egg discrimination. J. Anim. Ecol. 1989, 58, 207–224. [Google Scholar]
- Grim, T. An exceptionally high diversity of hoverflies (Syrphidae) in the food of the Reed Warbler (Acrocephalus scirpaceus). Biologia 2006, 61, 235–239. [Google Scholar] [CrossRef]
- Entomology Forum. Available online: http://www.entomologiitaliani.net/public/forum/phpBB3 (accessed on 10 January 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.r-project.org (accessed on 1 November 2022).
- Skaug, H.; Fournier, D.; Nielsen, A.; Magnusson, A.; Bolker, B. Glmmadmb Package (0.6 7.1 edn). 2013. Available online: http://glmmadmb.r-forge.r-project.org/ (accessed on 1 June 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csörgő, T. Nádirgó (Acrocephalus arundinaceus) és cserrego nádiposzáta (Acrocephalus scirpaceus) populációk táplálkozási—Niche vizsgálata [Diet niche-study on the populations of the Great Reed Warbler (Acrocephalus arundinaceus) and the Reed Warbler (Acrocephalus scirpaceus). Puszta 1983, 1, 71–80. [Google Scholar]
- Karasov, W.H. Digestion in birds: Chemical and physiological determinants and ecological implications. In Avian Foraging: Theory, Methodology and Applications. Studies in Avian Biology n° 13; Morrison, M.L., Ralph, C.J., Verner, J., Jehl, J.R., Jr., Eds.; Cooper Orn. Soc.: San Diego, CA, USA, 1990; pp. 391–415. [Google Scholar]
- Schifani, E.; University of Parma, Parma, Italy. Personal communication, 2021.
- Di Maggio, R.; Campobello, D.; Sarà, M. Lesser kestrel diet and agricultural intensification in the Mediterranean: An unexpected win-win solution? J. Nat. Cons. 2018, 45, 122–130. [Google Scholar] [CrossRef]
- Triolo, S.; Campobello, D.; Sarà, M. Diurnal habitat suitability for a Mediterranean steppeland bird, identified by Ecological Niche Factor Analysis. Wildl. Res. 2011, 38, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Brix, H. The European research project on reed die-back and progression (EUREED). Limnologica 1999, 29, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Møller, A.P.; Saino, N.; Adamík, P.; Ambrosini, R.; Antonov, A.; Campobello, D.; Stokke, B.G.; Fossøy, F.; Lehikoinen, E.; Martin-Vivaldi, M.; et al. Rapid change in host use of the common cuckoo Cuculus canorus linked to climate change. Proc. R Soc. B Biol. Sci. 2011, 278, 733–738. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference, Country | Sample Method | N of Samples | Araneae | Coleoptera | Heteroptera | Auchenorrhyncha, Aphidoidea | Diptera | Others |
|---|---|---|---|---|---|---|---|---|
| [14] Chernetsov and Manukyan (1999), Russia | Collar | 170 | 37 1 | 79 1 | 29 1 | 18.2 1 | 79 1 | - |
| [15] Kazlauskas Pukas and Meldazyte (1986) in [5] Cramp (1992), Lithuania | Collar | >2000 | 6.9 | 0.7 | 0.4 | 4 | 83.1 | 4.9 |
| [16] Dyrcz (1979), Poland | Collar 2 | 2549 | 2.7 | 0 | 0 | 4.3 | 71.9 | 21.1 |
| [4] Davies and Green (1976), UK | Collar | 120 | 5.8 | 5 | 0 | 10.9 | 64.9 | 13.3 |
| [17] Bibby and Thomas (1985), UK | Fecal | 1710 | 18 | 12.5 | 2.1 | 17.6 | 24.2 | 25.6 |
| [18] Grim and Honza (1996), Czech Republic | Collar 2 | 94 | 6.2 | 1.2 | 2.3 | 13.4 | 67.6 | 9.2 |
| [19,20] Henry (1977, 1979), France | Collar 2 | 2159 | 5.1 | 0.9 | 0.3 | 44.8 | 35.4 | 8.2 |
| [21] Bussmann (1979), France | Collar | 1776 | 15.8 | 0.9 | 0.4 | 14.3 | 51.2 | 32.6 |
| [22] Bibby and Green (1983), France | Fecal | 55 | 7.2 | 13.3 | 3.5 | 19.5 | 23.8 | 10 |
| [23] Kerbiriou et al. (2011), France | Fecal | 28 | 6.4 | 4 | 2.3 | 22 | 53.8 | 12.2 |
| [16] Dyrcz (1979), Switzerland | Collar 2 | 1839 | 3.7 | 0 | 0 | 0.3 | 91.2 | 4.8 |
| Akriotis in [5] Cramp (1992), Greece | Collar | 1018 | 8.3 | 2.1 | 1.7 | 30.9 | 50.7 | 6.8 |
| [24] Rguibi Idrissi et al. (2004) 3, Morocco | Collar | 88 | 2.7 | 27.4 | 4.4 | 4.5 | 1.9 | 59.1 4 |
| Prey Type | Mirandola (n = 149) | Gela (n = 308) | Vendicari (n = 103) | Pergusa (n = 202) | Total (n = 762) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| No | % | No | % | No | % | No | % | No | % | |
| Araneae | 0 | 0 | 1 | 0.3 | 0 | 0 | 0 | 0 | 1 | 0.01 |
| Coleoptera | 105 | 70.5 | 220 | 71.4 | 76 | 73.8 | 121 | 59.9 | 522 | 68.5 |
| Hymenoptera | 0 | 0 | 24 | 7.8 | 12 | 11.6 | 5 | 2.5 | 41 | 5.4 |
| Heteroptera | 36 | 24.2 | 9 | 2.9 | 0 | 0 | 10 | 4.9 | 55 | 7.2 |
| Aphidoidea | 8 | 5.4 | 3 | 1.0 | 0 | 0 | 57 | 28.2 | 68 | 9 |
| Others | 0 | 0 | 51 | 16.5 | 15 | 14.6 | 9 | 4.5 | 75 | 9.9 |
| Total | 149 | 100 | 308 | 100 | 103 | 100 | 202 | 100 | 762 | 100 |
| Year | 2018 | 2019 | Total | ||
|---|---|---|---|---|---|
| Prey Type | No | % | No | % | No |
| Araneae | 127 | 9.2 | 69 | 5.0 | 196 |
| Coleoptera | 82 | 5.9 | 383 | 28.0 | 465 |
| Diptera | 401 | 28.9 | 461 | 33.7 | 862 |
| Hymenoptera | 31 | 2.2 | 72 | 5.3 | 103 |
| Heteroptera | 675 | 48.7 | 359 | 26.3 | 1034 |
| Aphidoidea | 34 | 2.4 | 20 | 1.5 | 54 |
| Others | 34 | 2.4 | 2 | 0.1 | 36 |
| Total | 1384 | 99.7 | 1366 | 99.9 | 2750 |
| Prey Items | No. | % |
|---|---|---|
| Araneae | 8 | 34.8 |
| Heteroptera (Dimorphopterus blissoides) | 1 | 4.3 |
| Orthoptera Tettigoniidae 1 | 5 | 21.7 |
| Lepidoptera adults (moths) | 2 | 8.7 |
| Lepidoptera larvae | 3 | 13 |
| Others | 4 | 17.4 |
| Total | 23 | 99.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ientile, R.; Tuliozi, B.; Campobello, D.; Borghi, S.; Sala, L.; Dal Zotto, M.; Massa, B. Analysis of Prey Composition in Eurasian Reed Warblers’ Acrocephalus scirpaceus Droppings at Four Breeding Sites in Italy. Diversity 2022, 14, 1134. https://doi.org/10.3390/d14121134
Ientile R, Tuliozi B, Campobello D, Borghi S, Sala L, Dal Zotto M, Massa B. Analysis of Prey Composition in Eurasian Reed Warblers’ Acrocephalus scirpaceus Droppings at Four Breeding Sites in Italy. Diversity. 2022; 14(12):1134. https://doi.org/10.3390/d14121134
Chicago/Turabian StyleIentile, Renzo, Beniamino Tuliozi, Daniela Campobello, Stefano Borghi, Luigi Sala, Matteo Dal Zotto, and Bruno Massa. 2022. "Analysis of Prey Composition in Eurasian Reed Warblers’ Acrocephalus scirpaceus Droppings at Four Breeding Sites in Italy" Diversity 14, no. 12: 1134. https://doi.org/10.3390/d14121134
APA StyleIentile, R., Tuliozi, B., Campobello, D., Borghi, S., Sala, L., Dal Zotto, M., & Massa, B. (2022). Analysis of Prey Composition in Eurasian Reed Warblers’ Acrocephalus scirpaceus Droppings at Four Breeding Sites in Italy. Diversity, 14(12), 1134. https://doi.org/10.3390/d14121134