
Genetic Detection and a Method to Study the Ecology of Deadly Cubozoan Jellyfish



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Database Creation
2.2. Assay Design
2.3. Assay Validation and Optimisation
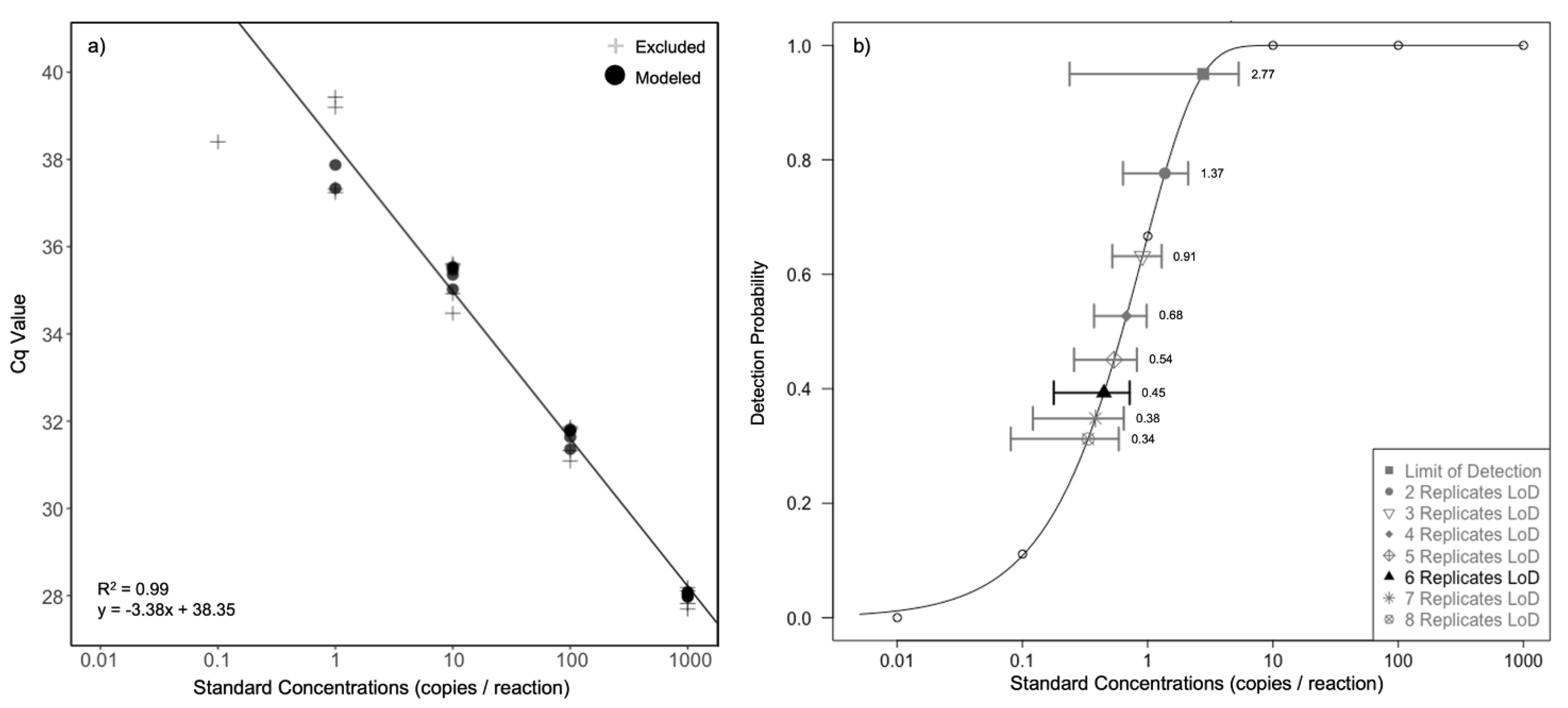
2.4. Assay Efficiency and Sensitivity
2.5. Endogenous Control Assay and Multiplexed qPCR
2.6. eDNA Collection and Preservation
2.7. eDNA Extraction and Purification
2.8. Quantitative PCR
2.9. In Situ Validation of Multiplexed Assays
2.10. Effect of Temperature upon C. fleckeri eDNA Decay
2.10.1. Collection of Study Organisms
2.10.2. Experimental Design
3. Results
3.1. Assay Design and In Vitro Validation
3.2. Assay Efficiency and Sensitivity
3.3. Multiplexed qPCR Assays
3.4. In Situ Validation
3.5. Chironex fleckeri eDNA Decay and the Influence of Temperature
4. Discussion
4.1. Assay Development and Informative Controls
4.2. eDNA Decay and Temperature Influence
4.3. Application and Implications of Cubozoan Detection with eDNA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castellani, C.; Edwards, M. Marine Plankton: A Practical Guide to Ecology, Methodology, and Taxonomy; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Kingsford, M. Biotic and abiotic structure in the pelagic environment: Importance to small fishes. Bull. Mar. Sci. 1993, 53, 393–415. [Google Scholar]
- Kingsford, M.J.; Becken, S.; Bordehore, C.; Fuentes, V.L.; Pitt, K.A.; Yangihara, A.A. Empowering stakeholders to manage stinging jellyfish: A perspective. Coast. Manag. 2018, 46, 1–18. [Google Scholar] [CrossRef]
- Decker, M.B.; Cieciel, K.; Zavolokin, A.; Lauth, R.; Brodeur, R.D.; Coyle, K.O. Population Fluctuations of jellyfish in the Bering Sea and their ecological role in this productive shelf ecosystem. In Jellyfish Blooms; Springer: Cham, Switzerland, 2014; pp. 153–183. [Google Scholar]
- Lynam, C.P.; Gibbons, M.J.; Axelsen, B.E.; Sparks, C.A.; Coetzee, J.; Heywood, B.G.; Brierley, A.S. Jellyfish overtake fish in a heavily fished ecosystem. Curr. Biol. 2006, 16, R492–R493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, W.M.; Martin, D.L.; Felder, D.L.; Asper, V.L.; Perry, H.M. Ecological and economic implications of a tropical jellyfish invader in the Gulf of Mexico. In Marine Bioinvasions: Patterns, Processes and Perspectives; Springer: Cham, Switzerland, 2003; pp. 53–69. [Google Scholar]
- Brodeur, R.D.; Sugisaki, H.; Hunt, G.L., Jr. Increases in jellyfish biomass in the Bering Sea: Implications for the ecosystem. Mar. Ecol. Prog. Ser. 2002, 233, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Uye, S.-I. The giant jellyfish Nemopilema nomurai in East Asian marginal seas. In Jellyfish Blooms; Springer: Cham, Switzerland, 2014; pp. 185–205. [Google Scholar]
- Bosch-Belmar, M.; Milisenda, G.; Basso, L.; Doyle, T.K.; Leone, A.; Piraino, S. Jellyfish impacts on marine aquaculture and fisheries. Rev. Fish. Sci. Aquac. 2020, 29, 242–259. [Google Scholar] [CrossRef]
- Collins, A.G.; Jarms, G. WoRMS Cubozoa: World list of Cubozoa (version 2018-04-01). In Species 2000 & ITIS Catalogue of Life, 2018 Annual Checklist; Roskov, Y., Abucay, L., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W., De Wever, A., et al., Eds.; Naturalis: Leiden, The Netherlands, 2018; Available online: www.catalogueoflife.org/annual-checklist/2018 (accessed on 1 September 2022).
- Crowley-Cyr, L.; Gershwin, L.-A. Protecting the public from hazardous jellyfish: A wicked problem for regulators and operators? The cnidaria: Only a problem or also a resource? In Marine and Freshwater Biology; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2021; pp. 99–122. [Google Scholar]
- Gershwin, L.A.; Richardson, A.J.; Winkel, K.D.; Fenner, P.J.; Lippmann, J.; Hore, R.; Avila-Soria, G.; Brewer, D.; Kloser, R.J.; Steven, A.; et al. Biology and ecology of Irukandji jellyfish (Cnidaria: Cubozoa). In Advances in Marine Biology; Academic Press: Washington, DC, USA, 2013; Volume 66, pp. 1–85. [Google Scholar]
- Kingsford, M.J.; Mooney, C.J. The ecology of box jellyfishes (Cubozoa). In Jellyfish Blooms; Springer: Cham, Switzerland, 2014; pp. 267–302. [Google Scholar]
- Brown, T.W. Chironex Fleckeri: Distribution and Movements Around Magnetic Island, North Queensland; TW Brown: Brooks, GA, USA, 1973. [Google Scholar]
- Pitt, K.; Kingsford, M. Geographic separation of stocks of the edible jellyfish Catostylus mosaicus (Rhizostomeae) in New South Wales, Australia. Mar. Ecol. Prog. Ser. 2000, 196, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Hartwick, R. Distributional ecology and behaviour of the early life stages of the box-jellyfish Chironex fleckeri. In Coelenterate Biology: Recent Research on Cnidaria and Ctenophora; Springer: Cham, Switzerland, 1991; pp. 181–188. [Google Scholar]
- Gordon, M.; Hatcher, C.; Seymour, J. Growth and age determination of the tropical Australian cubozoan Chiropsalmus sp. Hydrobiologia 2004, 530, 339–345. [Google Scholar]
- Bordehore, C.; Fuentes, V.L.; Atienza, D.; Barberá, C.; Fernandez-Jover, D.; Roig, M.; Acevedo-Dudley, M.J.; Canepa, A.J.; Gili, J.M. Detection of an unusual presence of the cubozoan Carybdea marsupialis at shallow beaches located near Denia, Spain (south-western Mediterranean). In Marine Biodiversity Records; Cambridge University Press: Cambridge, UK, 2011; Volume 4. [Google Scholar]
- Kingsford, M.; Seymour, J.; O’Callaghan, M. Abundance patterns of cubozoans on and near the Great Barrier Reef. In Jellyfish Blooms IV; Springer: Cham, Switzerland, 2012; pp. 257–268. [Google Scholar]
- Barnes, J. Studies on Three Venomous cubomedusae. The Cnidaria and Their Evolution: Symposium of the Zoological Society of London; Academic Press: London, UK, 1966. [Google Scholar]
- Llewellyn, L.; Bainbridge, S.; Page, G.; O’Callaghan, M.; Kingsford, M. StingerCam: A tool for ecologists and stakeholders to detect the presence of venomous tropical jellyfish. Limnol. Oceanogr. Methods 2016, 14, 649–657. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, G.; Zhang, M.; Jiang, Y. (Eds.) A method of jellyfish detection based on high resolution multibeam acoustic image. In MATEC Web of Conferences; EDP Sciences: Les Ulis, France, 2019. [Google Scholar]
- Lee, K.-H.; Kim, I.-O.; Yoon, W.-D.; Shin, J.-K.; An, H.-C. A study on vertical distribution observation of giant jellyfish (Nemopilema nomurai) using acoustical and optical methods. J. Korean Soc. Fish. Ocean. Technol. 2007, 43, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Rowley, O.C.; Courtney, R.L.; Browning, S.A.; Seymour, J.E. Bay watch: Using unmanned aerial vehicles (UAV’s) to survey the box jellyfish Chironex fleckeri. PLoS ONE 2020, 15, e0241410. [Google Scholar] [CrossRef]
- Schaub, J.; Hunt, B.P.; Pakhomov, E.A.; Holmes, K.; Lu, Y.; Quayle, L. Using unmanned aerial vehicles (UAVs) to measure jellyfish aggregations. Mar. Ecol. Prog. Ser. 2018, 591, 29–36. [Google Scholar] [CrossRef]
- Rowe, C.E.; Figueira, W.F.; Kelaher, B.P.; Giles, A.; Mamo, L.T.; Ahyong, S.T.; Keable, S.J. Evaluating the effectiveness of drones for quantifying invasive upside-down jellyfish (Cassiopea sp.) in Lake Macquarie, Australia. PLoS ONE 2022, 17, e0262721. [Google Scholar] [CrossRef] [PubMed]
- Beng, K.C.; Corlett, R.T. Applications of environmental DNA (eDNA) in ecology and conservation: Opportunities, challenges and prospects. Biodivers. Conserv. 2020, 29, 2089–2121. [Google Scholar] [CrossRef]
- Rees, H.C.; Maddison, B.C.; Middleditch, D.J.; Patmore, J.R.; Gough, K.C. The detection of aquatic animal species using environmental DNA–a review of eDNA as a survey tool in ecology. J. Appl. Ecol. 2014, 51, 1450–1459. [Google Scholar] [CrossRef]
- Wilcox, T.M.; McKelvey, K.S.; Young, M.K.; Jane, S.F.; Lowe, W.H.; Whiteley, A.R.; Schwartz, M.K. Robust detection of rare species using environmental DNA: The importance of primer specificity. PLoS ONE 2013, 8, e59520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerde, C.L.; Mahon, A.R.; Chadderton, W.L.; Lodge, D.M. “Sight-unseen” detection of rare aquatic species using environmental DNA. Conserv. Lett. 2011, 4, 150–157. [Google Scholar] [CrossRef]
- Shaw, J.L.; Weyrich, L.; Cooper, A. Using environmental (e) DNA sequencing for aquatic biodiversity surveys: A beginner’s guide. Mar. Freshw. Res. 2017, 68, 20–33. [Google Scholar] [CrossRef]
- Clare, E.L.; Economou, C.K.; Faulkes, C.G.; Gilbert, J.D.; Bennett, F.; Drinkwater, R.; Littlefair, J.E. eDNAir: Proof of concept that animal DNA can be collected from air sampling. PeerJ 2021, 9, e11030. [Google Scholar] [CrossRef]
- Thomsen, P.F.; Kielgast, J.; Iversen, L.L.; Møller, P.R.; Rasmussen, M.; Willerslev, E. Detection of a diverse marine fish fauna using environmental DNA from seawater samples. PLoS ONE 2012, 7, e41732. [Google Scholar] [CrossRef]
- Evans, N.T.; Shirey, P.D.; Wieringa, J.G.; Mahon, A.R.; Lamberti, G.A. Comparative cost and effort of fish distribution detection via environmental DNA analysis and electrofishing. Fisheries 2017, 42, 90–99. [Google Scholar]
- Sigsgaard, E.E.; Carl, H.; Møller, P.R.; Thomsen, P.F. Monitoring the near-extinct European weather loach in Denmark based on environmental DNA from water samples. Biol. Conserv. 2015, 183, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Barnes, M.A.; Turner, C.R. The ecology of environmental DNA and implications for conservation genetics. Conserv. Genet. 2016, 17, 1–17. [Google Scholar]
- Smart, A.S.; Weeks, A.R.; van Rooyen, A.R.; Moore, A.; McCarthy, M.A.; Tingley, R. Assessing the cost-efficiency of environmental DNA sampling. Methods Ecol. Evol. 2016, 7, 1291–1298. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.K.; Huerlimann, R.; Edmunds, R.C.; Budd, A.M.; Le Port, A.; Kyne, P.M.; Jerry, D.R.; Simpfendorfer, C.A. Improved detection sensitivity using an optimal eDNA preservation and extraction workflow and its application to threatened sawfishes. Aquat. Conserv. Mar. Freshwat. Ecosyst. 2021, 31, 2131–2148. [Google Scholar]
- Budd, A.M.; Cooper, M.K.; Le Port, A.; Schils, T.; Mills, M.S.; Deinhart, M.E.; Huerlimann, R.; Strugnell, J.M. First detection of critically endangered scalloped hammerhead sharks (Sphyrna lewini) in Guam, Micronesia, in five decades using environmental DNA. Ecol. Indic. 2021, 127, 107649. [Google Scholar] [CrossRef]
- Villacorta-Rath, C.; Espinoza, T.; Cockayne, B.; Schaffer, J.; Burrows, D. Environmental DNA analysis confirms extant populations of the cryptic Irwin’s turtle within its historical range. BMC Ecol. Evol. 2022, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.; Fukuda, Y.; Campbell, H.A. Using environmental DNA to detect estuarine crocodiles, a cryptic-ambush predator of humans. Hum. Wildl. Interact. 2020, 14, 11. [Google Scholar]
- Gaynor, J.J.; Bologna, P.A.; Restaino, D.J.; Barry, C.L. qPCR detection of early life history stage Chrysaora quinquecirrha (sea nettles) in Barnegat Bay, New Jersey. J. Coast. Res. 2017, 78 (Suppl. 1), 184–192. [Google Scholar] [CrossRef]
- Minamoto, T.; Fukuda, M.; Katsuhara, K.R.; Fujiwara, A.; Hidaka, S.; Yamamoto, S.; Takahashi, K.; Masuda, R. Environmental DNA reflects spatial and temporal jellyfish distribution. PLoS ONE 2017, 12, e0173073. [Google Scholar] [CrossRef] [Green Version]
- Bolte, B.; Goldsbury, J.; Huerlimann, R.; Jerry, D.; Kingsford, M. Validation of eDNA as a viable method of detection for dangerous cubozoan jellyfish. Environ. DNA 2021, 3, 769–779. [Google Scholar] [CrossRef]
- Ames, C.L.; Ohdera, A.H.; Colston, S.M.; Collins, A.G.; Fitt, W.K.; Morandini, A.C.; Erickson, J.S.; Vora, G.J. Fieldable environmental DNA sequencing to assess jellyfish biodiversity in Nearshore Waters of the Florida Keys, United States. Front. Mar. Sci. 2021, 8, 369. [Google Scholar] [CrossRef]
- Mychek-Londer, J.G.; Balasingham, K.D.; Heath, D.D. Using environmental DNA metabarcoding to map invasive and native invertebrates in two Great Lakes tributaries. Environ. DNA 2020, 2, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Kingsford, M.J.; Schlaefer, J.A.; Morrissey, S.J. Population structures and levels of connectivity for scyphozoan and cubozoan jellyfish. Diversity 2021, 13, 174. [Google Scholar] [CrossRef]
- Klymus, K.E.; Dannise, V.R.R.; Thompson, N.L.; Richter, C.A. Development and testing of species-specific quantitative PCR assays for environmental DNA applications. J. Vis. Exp. JOVE 2020, 165, e61825. [Google Scholar] [CrossRef]
- Coster, S.S.; Dillon, M.N.; Moore, W.; Merovich, G.T., Jr. The update and optimization of an eDNA assay to detect the invasive rusty crayfish (Faxonius rusticus). PLoS ONE 2021, 16, e0259084. [Google Scholar] [CrossRef]
- ThermoFisher. Design and Optimization of SYBR Green Assays; ThermoFisher: Waltham, MA, USA, 2018. [Google Scholar]
- Furlan, E.M.; Gleeson, D. Improving reliability in environmental DNA detection surveys through enhanced quality control. Mar. Freshw. Res. 2016, 68, 388–395. [Google Scholar] [CrossRef]
- Darling, J.A.; Mahon, A.R. From molecules to management: Adopting DNA-based methods for monitoring biological invasions in aquatic environments. Environ. Res. 2011, 111, 978–988. [Google Scholar] [CrossRef]
- Vuong, N.-M.; Villemur, R.; Payment, P.; Brousseau, R.; Topp, E.; Masson, L. Fecal source tracking in water using a mitochondrial DNA microarray. Water Res. 2013, 47, 16–30. [Google Scholar] [CrossRef]
- Harrison, J.B.; Sunday, J.M.; Rogers, S.M. Predicting the fate of eDNA in the environment and implications for studying biodiversity. Proc. R. Soc. B 2019, 286, 20191409. [Google Scholar] [CrossRef]
- Collins, R.A.; Wangensteen, O.S.; O’Gorman, E.J.; Mariani, S.; Sims, D.W.; Genner, M.J. Persistence of environmental DNA in marine systems. Commun. Biol. 2018, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Seymour, M.; Durance, I.; Cosby, B.J.; Ransom-Jones, E.; Deiner, K.; Ormerod, S.J.; Colbourne, J.; Wilgar, G.; Carvalho, G.R.; de Bruyn, M.; et al. Acidity promotes degradation of multi-species environmental DNA in lotic mesocosms. Commun. Biol. 2018, 1, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ely, T.; Barber, P.H.; Man, L.; Gold, Z. Short-lived detection of an introduced vertebrate eDNA signal in a nearshore rocky reef environment. PLoS ONE 2021, 16, e0245314. [Google Scholar] [CrossRef] [PubMed]
- Strickler, K.M.; Fremier, A.K.; Goldberg, C.S. Quantifying effects of UV-B, temperature, and pH on eDNA degradation in aquatic microcosms. Biol. Conserv. 2015, 183, 85–92. [Google Scholar] [CrossRef]
- Andruszkiewicz Allan, E.; Zhang, W.G.; CLavery, A.; FGovindarajan, A. Environmental DNA shedding and decay rates from diverse animal forms and thermal regimes. Environ. DNA 2021, 3, 492–514. [Google Scholar] [CrossRef]
- Barnes, M.A.; Turner, C.R.; Jerde, C.L.; Renshaw, M.A.; Chadderton, W.L.; Lodge, D.M. Environmental conditions influence eDNA persistence in aquatic systems. Environ. Sci. Technol. 2014, 48, 1819–1827. [Google Scholar] [CrossRef]
- Lamb, P.D.; Fonseca, V.G.; Maxwell, D.L.; Nnanatu, C.C. Systematic review and meta-analysis: Water type and temperature affect environmental DNA decay. Mol. Ecol. Resour. 2022, 22, 2494–2505. [Google Scholar] [CrossRef] [PubMed]
- Coordinators, N.R. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2015, 44, D7–D19. [Google Scholar]
- Deagle, B.E.; Jarman, S.N.; Coissac, E.; Pompanon, F.; Taberlet, P. DNA metabarcoding and the cytochrome c oxidase subunit I marker: Not a perfect match. Biol. Lett. 2014, 10, 20140562. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Vinh, L.S.; Flouri, T.; Stamatakis, A.; von Haeseler, A.; Minh, B.Q. MPBoot: Fast phylogenetic maximum parsimony tree inference and bootstrap approximation. BMC Evol. Biol. 2018, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bentlage, B.; Cartwright, P.; Yanagihara, A.A.; Lewis, C.; Richards, G.S.; Collins, A.G. Evolution of box jellyfish (Cnidaria: Cubozoa), a group of highly toxic invertebrates. Proc. R. Soc. B Biol. Sci. 2010, 277, 493–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mooney, C.; Kingsford, M. Sources and movements of Chironex fleckeri medusae using statolith elemental chemistry. In Jellyfish Blooms IV; Springer: Cham, Switzerland, 2012; pp. 269–277. [Google Scholar]
- Acevedo, M.J.; Straehler-Pohl, I.; Morandini, A.C.; Stampar, S.N.; Bentlage, B.; Matsumoto, G.I.; Yanagihara, A.; Toshino, S.; Bordehore, C.; Fuentes, V.L. Revision of the genus Carybdea (Cnidaria: Cubozoa: Carybdeidae): Clarifying the identity of its type species Carybdea marsupialis. Zootaxa 2019, 4543, 515–548. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.G.; Bentlage, B.; Gillan, W.; Lynn, T.H.; Morandini, A.C.; Marques, A.C. Naming the Bonaire banded box jelly, Tamoya ohboya, n. sp. (Cnidaria: Cubozoa: Carybdeida: Tamoyidae). Zootaxa 2011, 2753, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Prediger, E. How to Design Primers and Probes for PCR and qPCR. Integrated DNA Technologies. 2013. Available online: https://www.idtdna.com/pages/education/decoded/article/designing-pcr-primers-and-probes (accessed on 1 June 2020).
- Owczarzy, R.; Tataurov, A.V.; Wu, Y.; Manthey, J.A.; McQuisten, K.A.; Almabrazi, H.G.; Pedersen, K.F.; Lin, Y.; Garretson, J.; McEntaggart, N.O.; et al. IDT SciTools: A suite for analysis and design of nucleic acid oligomers. Nucleic Acids Res. 2008, 36 (Suppl. 2), W163–W169. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The sequence manipulation suite: JavaScript programs for analyzing and formatting protein and DNA sequences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Klymus, K.E.; Merkes, C.M.; Allison, M.J.; Goldberg, C.S.; Helbing, C.C.; Hunter, M.E.; Jackson, C.A.; Lance, R.F.; Mangan, A.M.; Monroe, E.M.; et al. Reporting the limits of detection and quantification for environmental DNA assays. Environ. DNA 2020, 2, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Roussel, J.M.; Paillisson, J.M.; Treguier, A.; Petit, E. The downside of eDNA as a survey tool in water bodies. J. Appl. Ecol. 2015, 52, 823–826. [Google Scholar] [CrossRef]
- Xiong, W.; Li, H.; Zhan, A. Early detection of invasive species in marine ecosystems using high-throughput sequencing: Technical challenges and possible solutions. Mar. Biol. 2016, 163, 139. [Google Scholar] [CrossRef]
- Edmunds, R.C.; Burrows, D. Got glycogen?: Development and multispecies validation of the novel preserve, precipitate, lyse, precipitate, purify (PPLPP) workflow for environmental DNA extraction from Longmire’s preserved water samples. J. Biomol. Tech. JBT 2020, 31, 125. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.K.; Villacorta-Rath, C.; Burrows, D.; Jerry, D.R.; Carr, L.; Barnett, A.; Huveneers, C.; Simpfendorfer, C.A. Practical eDNA sampling methods inferred from particle size distribution and comparison of capture techniques for a Critically Endangered elasmobranch. Environ. DNA 2022, 4, 1011–1023. [Google Scholar] [CrossRef]
- Langlois, V.S.; Allison, M.J.; Bergman, L.C.; To, T.A.; Helbing, C.C. The need for robust qPCR-based eDNA detection assays in environmental monitoring and species inventories. Environ. DNA 2021, 3, 519–527. [Google Scholar] [CrossRef]
- Sassoubre, L.M.; Yamahara, K.M.; Gardner, L.D.; Block, B.A.; Boehm, A.B. Quantification of environmental DNA (eDNA) shedding and decay rates for three marine fish. Environ. Sci. Technol. 2016, 50, 10456–10464. [Google Scholar] [CrossRef] [PubMed]
- Kwong, S.L.; Villacorta-Rath, C.; Doyle, J.; Uthicke, S. Quantifying shedding and degradation rates of environmental DNA (eDNA) from Pacific crown-of-thorns seastar (Acanthaster cf. solaris). Mar. Biol. 2021, 168, 1–10. [Google Scholar] [CrossRef]
- Wood, S.A.; Biessy, L.; Latchford, J.L.; Zaiko, A.; von Ammon, U.; Audrezet, F.; Cristescu, M.E.; Pochon, X. Release and degradation of environmental DNA and RNA in a marine system. Sci. Total Environ. 2020, 704, 135314. [Google Scholar] [CrossRef] [PubMed]
- Schlaefer, J.A.; Wolanski, E.; Kingsford, M.J. Swimming behaviour can maintain localised jellyfish (Chironex fleckeri: Cubozoa) populations. Mar. Ecol. Prog. Ser. 2018, 591, 287–302. [Google Scholar] [CrossRef]
- Shogren, A.J.; Tank, J.L.; Andruszkiewicz, E.A.; Olds, B.; Jerde, C.; Bolster, D. Modelling the transport of environmental DNA through a porous substrate using continuous flow-through column experiments. J. R. Soc. Interface 2016, 13, 20160290. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Huang, Q.; Zhang, X.; Chen, H. Adsorption of DNA on clay minerals and various colloidal particles from an Alfisol. Soil. Biol. Biochem. 2006, 38, 471–476. [Google Scholar] [CrossRef]
- Jo, T.; Murakami, H.; Yamamoto, S.; Masuda, R.; Minamoto, T. Effect of water temperature and fish biomass on environmental DNA shedding, degradation, and size distribution. Ecol. Evol. 2019, 9, 1135–1146. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, S.; Ushio, M.; Sakurai, S.; Minamoto, T.; Yamanaka, H. Water temperature-dependent degradation of environmental DNA and its relation to bacterial abundance. PLoS ONE 2017, 12, e0176608. [Google Scholar] [CrossRef] [PubMed]
- Australian Institute of Marine Science (AIMS). AIMS Sea Water Temperature Observing System (AIMS Temperature Logger Program). 2017. Available online: https://apps.aims.gov.au/metadata/view/4a12a8c0-c573-11dc-b99b-00008a07204e (accessed on 1 June 2020).
- Eichmiller, J.J.; Best, S.E.; Sorensen, P.W. Effects of temperature and trophic state on degradation of environmental DNA in lake water. Environ. Sci. Technol. 2016, 50, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Deiner, K.; Bik, H.M.; Mächler, E.; Seymour, M.; Lacoursière-Roussel, A.; Altermatt, F.; Creer, S.; Bista, I.; Lodge, D.M.; De Vere, N.; et al. Environmental DNA metabarcoding: Transforming how we survey animal and plant communities. Mol. Ecol. 2017, 26, 5872–5895. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.R.; Clark, Z.; Treml, E.A.; Brown, M.S.; Matthews, T.G.; Pocklington, J.B.; Stafford-Bell, R.E.; Bott, N.J.; Nai, Y.H.; Miller, A.D.; et al. Detecting marine pests using environmental DNA and biophysical models. Sci. Total Environ. 2022, 816, 151666. [Google Scholar] [CrossRef]
- Mooney, C.J.; Kingsford, M.J. Discriminating populations of medusae (Chironex fleckeri, Cubozoa) using statolith microchemistry. Mar. Freshw. Res. 2017, 68, 1144–1152. [Google Scholar] [CrossRef]
- Mooney, C.J.; Kingsford, M.J. Statolith morphometrics can discriminate among taxa of cubozoan jellyfishes. PLoS ONE 2016, 11, e0155719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Port, J.A.; O’Donnell, J.L.; Romero-Maraccini, O.C.; Leary, P.R.; Litvin, S.Y.; Nickols, K.J.; Yamahara, K.M.; Kelly, R.P. Assessing vertebrate biodiversity in a kelp forest ecosystem using environmental DNA. Mol. Ecol. 2016, 25, 527–541. [Google Scholar] [CrossRef]
- Kelly, R.P.; Gallego, R.; Jacobs-Palmer, E. The effect of tides on nearshore environmental DNA. PeerJ 2018, 6, e4521. [Google Scholar] [CrossRef] [Green Version]
- Takahara, T.; Minamoto, T.; Yamanaka, H.; Doi, H.; Kawabata, Z. Estimation of fish biomass using environmental DNA. PLoS ONE 2012, 7, e35868. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, P.F.; Kielgast, J.; Iversen, L.L.; Wiuf, C.; Rasmussen, M.; Gilbert, M.T.P.; Orlando, L.; Willerslev, E. Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 2012, 21, 2565–2573. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Estimating occupancy and abundance of stream amphibians using environmental DNA from filtered water samples. Can. J. Fish. Aquat. Sci. 2013, 70, 1123–1130. [Google Scholar] [CrossRef]
- Lacoursière-Roussel, A.; Côté, G.; Leclerc, V.; Bernatchez, L. Quantifying relative fish abundance with eDNA: A promising tool for fisheries management. J. Appl. Ecol. 2016, 53, 1148–1157. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, T.M.; McKelvey, K.S.; Young, M.K.; Sepulveda, A.J.; Shepard, B.B.; Jane, S.F.; Whiteley, A.R.; Lowe, W.H.; Schwartz, M.K. Understanding environmental DNA detection probabilities: A case study using a stream-dwelling char Salvelinus fontinalis. Biol. Conserv. 2016, 194, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Minami, K.; Fukaya, K.; Takahashi, K.; Sawada, H.; Murakami, H.; Tsuji, S.; Hashizume, H.; Kubonaga, S.; Horiuchi, T.; et al. Environmental DNA as a ‘snapshot’of fish distribution: A case study of Japanese jack mackerel in Maizuru Bay, Sea of Japan. PLoS ONE 2016, 11, e0149786. [Google Scholar]
- Richardson, A.J.; Bakun, A.; Hays, G.C.; Gibbons, M.J. The jellyfish joyride: Causes, consequences and management responses to a more gelatinous future. Trends Ecol. Evol. 2009, 24, 312–322. [Google Scholar] [CrossRef]
- Madin, E.M.; Ban, N.C.; Doubleday, Z.A.; Holmes, T.H.; Pecl, G.T.; Smith, F. Socio-economic and management implications of range-shifting species in marine systems. Glob. Environ. Change 2012, 22, 137–146. [Google Scholar] [CrossRef]
- Orellana, E.R.; Collins, A.G. First report of the box jellyfish Tripedalia cystophora (Cubozoa: Tripedaliidae) in the continental USA, from Lake Wyman, Boca Raton, Florida. Mar. Biodivers. Rec. 2011, 4, e54. [Google Scholar] [CrossRef] [Green Version]
- Sathirapongsasuti, N.; Khonchom, K.; Poonsawat, T.; Pransilpa, M.; Ongsara, S.; Detsri, U.; Bungbai, S.; Lawanangkoon, S.-A.; Pattanaporkrattana, P.W.; Trakulsrichai, S. Rapid and accurate species-specific PCR for the identification of lethal chironex box jellyfish in Thailand. Int. J. Environ. Res. Public Health 2021, 18, 219. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession No. | Country of Origin | Source |
|---|---|---|---|
| Chironex fleckeri | GQ849101 (E) | Darwin, Australia | [66] |
| GQ849102 (E) | Weipa, Australia | [66] | |
| GQ849103 (E) | Weipa, Australia | [66] | |
| OP877024 (N) | Townsville, Australia | SLSQ | |
| OP877025 (N) | Townsville, Australia | SLSQ | |
| OP877026 (N) | Weipa, Australia | [67] | |
| OP877027 (N) | Weipa, Australia | [67] | |
| OP877028 (N) | Weipa, Australia | SLSQ | |
| OP877029 (N) | Mackay, Australia | [67] | |
| OP877030 (N) | Mackay, Australia | [67] | |
| OP877031 (N) | Darwin, Australia | [67] | |
| Carukia barnesi | GQ849097 (E) | Cairns, Australia | [66] |
| GQ849098 (E) | Cairns, Australia | [66] | |
| OP877033 (N) | Palm Cove, Australia | Sample provided by Jamie Seymour to ROEL | |
| Alatina alata | GQ506980 (E) | Osprey Reef, Australia | [66] |
| OP877035 (N) | Waikiki, Hawaii | Sample provided by Lisa Gershwin to ROEL | |
| Carybdea xaymacana | KT288254 (E) | Puerto Rico, Caribbean | [68] |
| GQ849114 (E) | Panama, Caribbean | [66] | |
| GQ849115 (E) | Panama, Caribbean | [66] | |
| GQ849118 (E) | Panama, Caribbean | [66] | |
| OP877034 (N) | Townsville, Australia | ROEL | |
| Copula sivickisi | GQ849113 (E) | Cairns, Australia | [66] |
| OP877032 (N) | Townsville, Australia | ROEL | |
| Tamoya ohboya | HQ824528 (E) | Bonaire, Caribbean Netherlands | [69] |
| Assay | Label | Sequence (5′-3′) | Target Sequence Length (bp) |
|---|---|---|---|
| Species-specific Assay—C. fleckeri | Chironex_16S_F | ATCTTCCACTGTCTCAGCTTTACC | 147 |
| Chironex_16S_R | CCTCAGTACTCGTGTCTCCCTA | ||
| Chironex_16S_P | (FAM)-CTCGTCCTTCCAAGTATAAG-(MGB) | ||
| Endogenous Control Assay—Generic Fish | Fish_16S_F | GACCTCGATGTTGGATCA | 87–88 |
| Fish_16S_R | CTCAGATCACGTAGGACTTTA | ||
| Fish_16S_probe | (VIC)-ACATCCTAWTGGTGC-(MGB) |
| Location | Site | C. fleckeri Assay | Endogenous Control Assay |
|---|---|---|---|
| Weipa | 1 | 7/12 | 12/12 |
| Weipa | 2 | 6/12 | 12/12 |
| Horseshoe Bay | 1 | 2/12 | 12/12 |
| Horseshoe Bay | 2 | 1/12 | 12/12 |
| Horseshoe Bay | 3 | 5/12 | 12/12 |
| Horseshoe Bay | 4 | 3/12 | 12/12 |
| Maud Bay | 5 | 0/12 | 12/12 |
| Maud Bay | 6 | 0/12 | 12/12 |
| Species | Decay Constant | Time to Decay | Experimental Temperature | Source |
|---|---|---|---|---|
| Chironex fleckeri | 0.060 h−1 | 27 h (99% decay) | 26–28 °C | Present study |
| Chrysaora pacifica | 0.033 h−1 | 4 days (90% decay) | 17.1–20 °C | [43] |
| Copula sivickisi | 0.034 h−1 | 9 days (99% decay) | 20 °C | [44] |
| Engraulis mordax | 0.101 h−1 | 3–4 days (detection limit) | 22 °C | [81] |
| Sardinops sagax | 0.068 h−1 | 3–4 days (detection limit) | 18.7 °C | [81] |
| Scomber japonicas | 0.070 h−1 | 3–4 days (detection limit) | 18.7 °C | [81] |
| Acanthaster spp. | 0.048 h−1 | 96 h (99% decay) | 24–28 °C | [82] |
| Styela clava | 0.104 h−1 | 94 h (detection limit) | 19 °C | [83] |
| Sabella spallanzanii | 0.248 h−1 | 42 h (detection limit) | 19 °C | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrissey, S.J.; Jerry, D.R.; Kingsford, M.J. Genetic Detection and a Method to Study the Ecology of Deadly Cubozoan Jellyfish. Diversity 2022, 14, 1139. https://doi.org/10.3390/d14121139
Morrissey SJ, Jerry DR, Kingsford MJ. Genetic Detection and a Method to Study the Ecology of Deadly Cubozoan Jellyfish. Diversity. 2022; 14(12):1139. https://doi.org/10.3390/d14121139
Chicago/Turabian StyleMorrissey, Scott J., Dean R. Jerry, and Michael J. Kingsford. 2022. "Genetic Detection and a Method to Study the Ecology of Deadly Cubozoan Jellyfish" Diversity 14, no. 12: 1139. https://doi.org/10.3390/d14121139
APA StyleMorrissey, S. J., Jerry, D. R., & Kingsford, M. J. (2022). Genetic Detection and a Method to Study the Ecology of Deadly Cubozoan Jellyfish. Diversity, 14(12), 1139. https://doi.org/10.3390/d14121139