
Diversity of Wood-Decaying Fungi in Wuliangshan Area, Yunnan Province, P.R. China



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Herbarium Specimen Preparation
2.2. Morphological Studies
2.3. Molecular Procedures and Phylogenetic Analysis



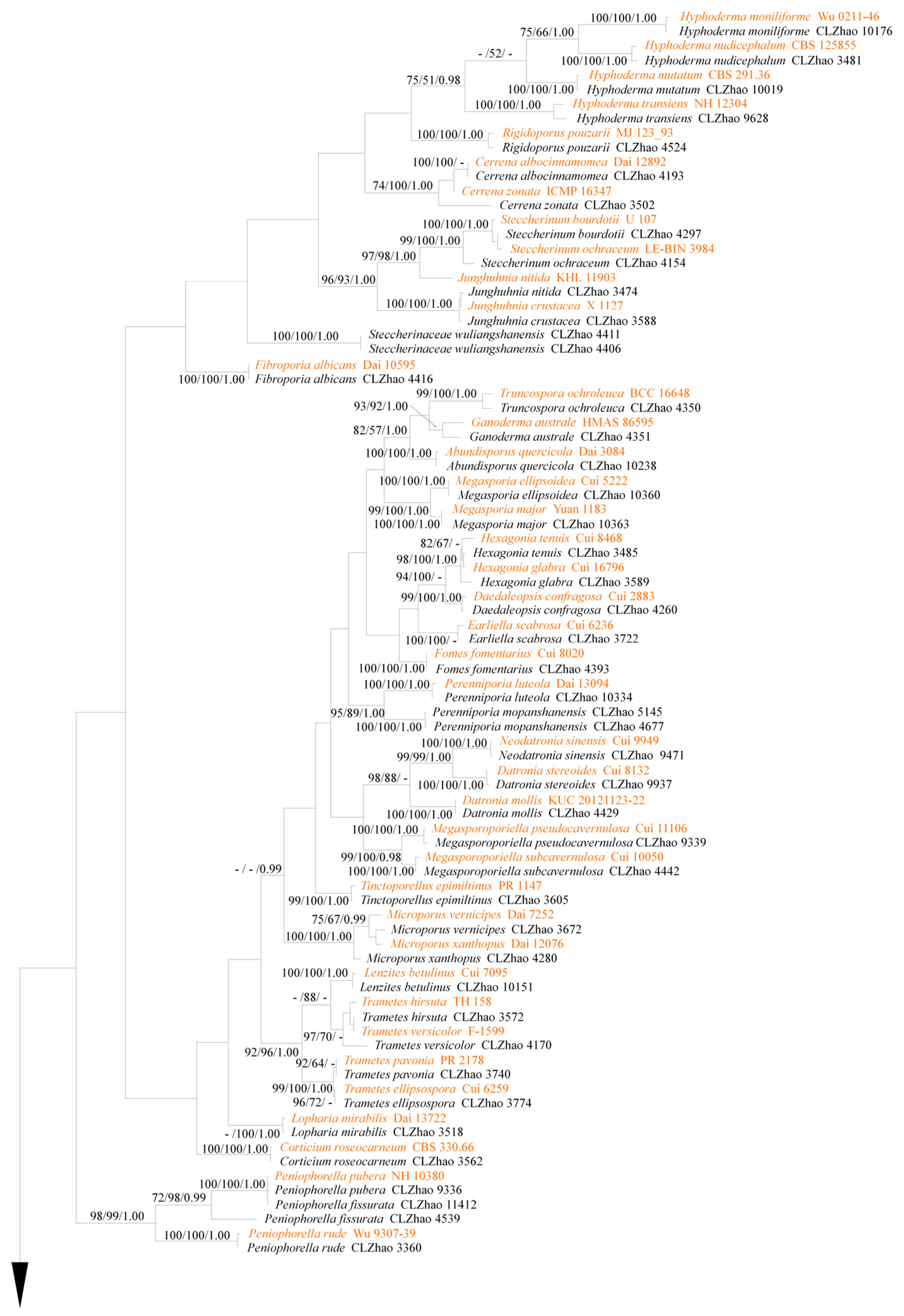
3. Results
3.1. Molecular Phylogeny
3.2. Checklist
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- James, T.Y.; Stajich, J.E.; Hittinger, C.T.; Rokas, A. Toward a fully resolved fungal tree of life. Annu. Rev. Microbiol. 2020, 74, 291–313. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, M. The Fungi: 1, 2, 3 … 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Hollingsworth, T.N.; Mcfarland, J.W. A first comprehensive census of fung i in soil reveals bothhyperdiversity and fine-scale niche partitioning. Ecol. Monogr. 2014, 84, 3–20. [Google Scholar] [CrossRef]
- Hibbett, D. The invisible dimension of fungal diversity. Science 2016, 351, 1150. [Google Scholar] [CrossRef]
- Russell, R.; Paterson, M. Ganoderma—A therapeutic fungal biofactory. Phytochemistry 2006, 67, 1985–2001. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C. Polypore diversity in China with an annotated checklist of Chinese polypores. Mycoscience 2012, 53, 49–80. [Google Scholar] [CrossRef]
- Dai, Y.C.; Cui, B.K.; Si, J.; He, S.H.; Hyde, K.D.; Yuan, H.S.; Liu, X.Y.; Zhou, L.W. Dynamics of the worldwide number of fungi with emphasis on fungal diversity in China. Mycol. Prog. 2015, 14, 62. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumara, S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef]
- M’Barek, H.N.; Arif, S.; Taidi, B.; Hajjaj, H. Consolidated bioethanol production from olive mill waste: Wood-decay fungi from central Morocco as promising decomposition and fermentation biocatalysts. Biotechnol. Rep. 2020, 28, e00541. [Google Scholar] [CrossRef]
- Runnel, K.; Miettinen, O.; Lohmus, A. Polypore fungi as a flagship group to indicate changes in biodiversity—A test case from Estonia. IMA Fungus 2021, 12, 2. [Google Scholar] [CrossRef]
- Yu, Q.G. Wuliangshan National Nature Reserve; Yunnan Science and Technology Press: Kunming, China, 2004; pp. 103–105, 118, 197. [Google Scholar]
- Lu, S.Z.; Yu, Q.G.; Cao, S.W. Measurement of ecosystem diversity in Wuliangshan National Nature Reserve and its adjacent area in Yunnan Province. J. Anhui Agric. Sci. 2008, 36, 2426–2428, 2435. [Google Scholar]
- Luo, Z.H.; Xie, Y.N.; Luo, Y.; Li, X.Y.; Lu, C.R. Population Characteristics of Tsuga dumosa in Wuliangshan National Nature Reserve. J. West China For. Sci. 2013, 42, 94–99. [Google Scholar]
- Gao, L.L.; Yang, H.J.; Liu, G.Z.; Wang, C.H.; Yang, G.P.; Cai, C.T. Diversity of medicinal vascular plant resources in Wuliangshan National Nature Reserve, Yunnan, China. Plant Sci. J. 2018, 36, 203–210. [Google Scholar]
- Peng, H.; Wu, Z.Y. The Preliminary Floristical study on mid-montane humid evergreen broad-leaved forest in Mt. Wuliangshan. Acta Bot. Yunnanica 1998, 20, 12–22. [Google Scholar]
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Wu, G.; Yuan, H.S.; Zhou, L.W.; He, S.H.; Ge, Z.W.; Wu, F.; Wei, Y.L.; et al. Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 2021, 40, 770–805. [Google Scholar] [CrossRef]
- Petersen, J.H. Farvekort. The Danish Mycological Society’s Colour-Chart; Foreningen til Svampekundskabens Fremme: Paris, France, 1996; pp. 1–6. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- Xu, T.M. Study on the Resources and Taxonomy of Wood-Decaying Fungi in Wuliangshan National Nature Reserve, Yunnan Province; Southwest Forestry University: Kunming, China, 2020. [Google Scholar]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Tian, Y.; Ghobad-Nejhad, M.; He, S.H.; Dai, Y.C. Three new species of Aleurodiscus s.l. (Russulales, Basidiomycota) from southern China. MycoKeys 2018, 37, 93–107. [Google Scholar] [CrossRef]
- Asl, A.; Syo, A.; Msp, A.; Kim, T.; Yoo, J.S.; Seelan, J.S.S.; Lim, Y.W. Co-occurrence patterns of wood-decaying fungi and ants in dead pines of South Korea—ScienceDirect. J. Asia Pac. Entomol. 2019, 22, 1154–1160. [Google Scholar] [CrossRef]
- Bandara, A.R.; Karunarathna, S.C.; Phillips, A.J.L.; Mortimer, P.E.; Xu, J.C.; Kakumyan, P.; Hyde, K.D. Diversity of Auricularia (Auriculariaceae, Auriculariales) in Thailand. Phytotaxa 2017, 292, 19. [Google Scholar] [CrossRef]
- Wu, F.; Yuan, Y.; Rivoire, B.; Dai, Y.C. Phylogeny and diversity of the Auricularia mesenterica (Auriculariales, Basidiomycota) complex. Mycol. Prog. 2015, 14, 42. [Google Scholar] [CrossRef]
- Floudas, D.; Hibbett, D.S. Revisiting the taxonomy of Phanerochaete (Polyporales, Basidiomycota) using a four gene dataset and extensive ITS sampling. Fungal Biol. 2015, 119, 679–719. [Google Scholar] [CrossRef]
- Águeda, B.; Parladé, J.; Fernandez-Toiran, L.M.; Cisneros, Ó.; Miguel, A.M.; Modrego, M.P.; Martínez-Peña, F.; Pera, J. Mycorrhizal synthesis between Boletus edulis species complex and rockroses (Cistus sp.). Mycorrhiza 2008, 18, 443–449. [Google Scholar] [CrossRef]
- Deo, G.S.; Khatra, J.; Buttar, S.; Wai, M.L.; Chow, H.L. Anti-proliferative, immuno-stimulatory and anti-inflammatory activities of extracts derived from mushrooms collected in Haida Gwaii, BC (Canada). Int. J. Med. Mushrooms 2019, 21, 629–643. [Google Scholar] [CrossRef]
- Shirouzu, T.; Hosaka, K.; Nam, K.O.; Weir, B.S.; Hosoya, T. Phylogenetic relationships of eight new Dacrymycetes collected from New Zealand. Persoonia 2017, 38, 156–169. [Google Scholar] [CrossRef] [Green Version]
- Shirouzu, T.; Hirose, D.; Oberwinkler, F.; Shimomura, N.; Maekawa, N.; Tokumasu, S. Combined molecular and morphological data for improving phylogenetic hypothesis in Dacrymycetes. Mycologia 2013, 105, 1110–1125. [Google Scholar] [CrossRef]
- Zamora, J.C.; Ekman, S. Phylogeny and character evolution in the Dacrymycetes, and systematics of Unilacrymaceae and Dacryonaemataceae fam. nov. Persoonia 2020, 44, 161–205. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Li, H.J.; Cui, B.K. Molecular phylogeny and taxonomy of Fibroporia (Basidiomycota) in China. Phytotaxa 2015, 203, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Bateman, C.C.; Skelton, J.; Jusino, M.A.; Nolen, Z.J.; Simmons, D.R.; Hulcr, J. Wood decay fungus Flavodon ambrosius (Basidiomycota: Polyporales) is widely farmed by two genera of ambrosia beetles. Fungal Biol. 2017, 121, 984–989. [Google Scholar] [CrossRef]
- Jeppson, M.; Nilsson, R.H.; Larsson, E. European earthstars in Geastraceae (Geastrales, Phallomycetidae)—A systematic approach using morphology and molecular sequence data. Syst. Biodivers. 2013, 11, 437–465. [Google Scholar] [CrossRef]
- Larsson, K.H. Re-thinking the classification of corticioid fungi. Mycol. Res. 2007, 111, 1040–1063. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Chen, J.J.; Korhonen, K.; Martin, F.; Dai, Y.C. An updated global species diversity and phylogeny in the forest pathogenic genus Heterobasidion (Basidiomycota, Russulales). Front. Microbiol. 2021, 11, 596393. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Wu, D.M.; Song, C.G.; Liu, S.; Si, J.; Cui, B.K. Two new Neofomitella species (Polyporaceae, Basidiomycota) based on morphological and molecular evidence. Mycol. Prog. 2019, 18, 593–602. [Google Scholar] [CrossRef]
- Li, H.J.; Cui, B.K.; Dai, Y.C. Taxonomy and multi-gene phylogeny of Datronia (Polyporales, Basidiomycota). Persoonia 2014, 32, 170–182. [Google Scholar] [CrossRef] [Green Version]
- Yurchenko, E.; Wu, S.H. Hyphoderma moniliforme and H. nemorale (Basidiomycota) newly recorded from China. Mycosphere 2015, 6, 113–121. [Google Scholar] [CrossRef]
- Larsson, K.H. Molecular phylogeny of Hyphoderma and the reinstatement of Peniophorella. Mycol. Res. 2007, 111, 186–195. [Google Scholar] [CrossRef]
- Sulzbacher, M.A.; Grebenc, T.; Nouhra, E.R.; Giachini, A.J.; Martín, M.P.; Baseia, L.G. Hysterangium atlanticum sp. nov. forms ectomycorrhizae with Coccoloba species (Polygonaceae) from the Atlantic rainforest of Northeastern Brazil. Symbiosis 2019, 78, 275–286. [Google Scholar] [CrossRef]
- Miettinen, O.; Larsson, E.; Sjokvist, E.; Larsson, K.H. Comprehensive taxon sampling reveals unaccounted diversity and morphological plasticity in a group of dimitic polypores (Polyporales, Basidiomycota). Cladistics Int. J. Willi Hennig Soc. 2012, 28, 251–270. [Google Scholar] [CrossRef] [Green Version]
- Mudalungu, C.M.; Richter, C.; Wittstein, K.; Abdalla, M.A.; Matasyoh, J.C.; Stadler, M.; Süssmuth, R.D. Laxitextines A and B, Cyathane Xylosides from the Tropical Fungus Laxitextum incrustatum. J. Nat. Prod. 2016, 79, 894–898. [Google Scholar] [CrossRef]
- Liu, S.L.; Nakasone, K.K.; Wu, S.H.; He, S.H.; Dai, Y.C. Taxonomy and phylogeny of Lopharia s.s Dendrodontia, Dentocorticium and Fuscocerrena (Basidiomycota, Polyporales). MycoKeys 2018, 32, 25–48. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, O.; Spirin, V.; Josef, V.; Rivoire, B.; Hibbett, D. Polypores and genus concepts in Phanerochaetaceae (Polyporales, Basidiomycota). MycoKeys 2016, 17, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Hallenberg, N.; Nilsson, R.H.; Antonelli, A.; Wu, S.H.; Maekawa, N.; Nordén, B. The Peniophorella praetermissa species complex (Basidiomycota). Mycol. Res. 2008, 111, 1366–1376. [Google Scholar] [CrossRef]
- Cabral, T.S.; Silva, B.D.; María, P.M.; Clement, C.R.; Hosaka, K.; Baseia, I.G. Behind the veil—Exploring the diversity in Phallus indusiatus s.l. (Phallomycetidae, Basidiomycota). MycoKeys 2019, 58, 103–127. [Google Scholar] [CrossRef]
- Volobuev, S.; Okun, M.; Ordynets, A.; Spirin, V. The Phanerochaete sordida group (Polyporales, Basidiomycota) in temperate Eurasia, with a note on Phanerochaete pallida. Mycol. Prog. 2015, 14, 80. [Google Scholar] [CrossRef]
- Melera, S.; Ostellari, C.; Roemer, N.; Avis, P.G.; Tonolla, M.; Barja, F.; Narduzzi-Wicht, B. Analysis of morphological, ecological and molecular characters of Russula pectinatoides Peck and Russula praetervisa Sarnari, with a description of the new taxon Russula recondita Melera & Ostellari. Mycol. Prog. 2016, 16, 117–134. [Google Scholar] [CrossRef]
- Palmer, J.M.; Lindner, D.L.; Volk, T.J. Ectomycorrhizal characterization of an American chestnut (Castanea dentata)-dominated community in Western Wisconsin. Mycorrhiza 2008, 19, 27–36. [Google Scholar] [CrossRef]
- Alshammari, N.; Stephenson, S.L. A preliminary study of wood-decay fungi in forests of northwest Arkansas. Curr. Res. Environ. Appl. Mycol. 2018, 8, 556–563. [Google Scholar] [CrossRef]
- Justo, A.; Miettinen, O.; Floudas, D.; Ortiz-Santana, B.; Sjökvist, E.; Lindner, D.; Nakasone, K.; Niemelä, T.; Larsson, K.H.; Ryvarden, L.; et al. A revised family-level classification of the Polyporales (Basidiomycota). Fungal Biol. 2017, 121, 798–824. [Google Scholar] [CrossRef]
- Zhao, C.L.; Cui, B.K.; Steffen, K.T. Yuchengia, a new polypore genus segregated from Perenniporia (Polyporales) based on morphological and molecular evidence. Nord. J. Bot. 2013, 31, 331–338. [Google Scholar] [CrossRef]
- Milenkovic, I.; Tomsovsky, M.; Karadzic, D.; Veselinovic, M. Decline of Paulownia tomentosa caused by Trametes hirsuta in Serbia. For. Pathol. 2018, 48, e12438. [Google Scholar] [CrossRef]
- Justo, A.; Hibbett, D.S. Phylogenetic classification of Trametes (Basidiomycota, Polyporales) based on a five–marker dataset. Taxon 2011, 60, 1567–1583. [Google Scholar] [CrossRef]
- Telleria, M.T.; Melo, I.; Duenas, M.; Larsson, K.H.; Martín, M.P.P. Molecular analyses confirm Brevicellicium in Trechisporales. IMA Fungus 2013, 4, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. Bioedit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Zhao, C.L.; Wu, Z.Q. Ceriporiopsis kunmingensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analysis. Mycol. Prog. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling High-Impact Science for Phylogenetics Researchers with Limited Resources. Assoc. Comput. Mach. 2012, 39, 1–8. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C. A revised checklist of corticioid and hydnoid fungi in China for 2010. Mycoscience 2011, 52, 69–79. [Google Scholar] [CrossRef]
- Zhao, C.L. Wood Inhabiting Fungi Diversity in Wuliangshan Area; China Forestry Publishing House: Beijing, China, 2021. [Google Scholar]
- Willis, K.J. State of the World’s Fungi 2018; Report; Royal Botanic Gardens: Kew, UK, 2018. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Ayesha, P.; Maham, A.; Ariza, A.; Husnain, H. “Ganoderma lucidum sensu lato”—A sacred mushroom for immortality. Stud. Fungi 2020, 5, 508–516. [Google Scholar] [CrossRef]
- Huang, R.X.; Luo, K.Y.; Zhao, C.L. Phlebia nigrodontea sp. nov. in Meruliaceae (Polyporales) with a black hymenial surface. Phytotaxa 2020, 458, 195–206. [Google Scholar] [CrossRef]
- Himani, S.; Krishnappa, M. Xylaria oxyacanthae (Xylariaceae), a new record on Diospyros melanoxylon from India. Stud. Fungi 2020, 5, 485–490. [Google Scholar] [CrossRef]
- Blanco-Dios, J.B. Notes on the genus Entoloma s.l. in the Northwest of the Iberian Peninsula (XI): A new species in subgenus Entoloma. Stud. Fungi 2021, 6, 64–70. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, R. Global diversity and distribution of distoseptosporic micromycete Corynespora Güssow (Corynesporascaceae): An updated checklist with current status. Stud. Fungi 2021, 6, 1–63. [Google Scholar] [CrossRef]
- Wang, H.; Zhao, C.L. Hyphodermella aurantiaca sp. nova. (Polyporales, Basidiomycota) as evidenced by morphological characters and phylogenetic analyses. Ann. Bot. Fenn. 2021, 58, 61–68. [Google Scholar] [CrossRef]
- Zong, T.K.; Liu, C.M.; Wu, J.R.; Zhao, C.L. Trechispora daweishanensis and T. xantha spp. nov. (Hydnodontaceae, Trechisporales) found in Yunnan Province of China. Phytotaxa 2021, 479, 147–159. [Google Scholar] [CrossRef]
- Boonyuen, N.; Chuaseeharonnachai, C.; Nuankaew, S.; Kwantong, P.; Pornputtapong, N.; Suwannarach, N.; Jones, E.B.G.; Somrithipol, S. Novelties in Fuscosporellaceae (Fuscosporellales): Two New Parafuscosporella from Thailand Revealed by Morphology and Phylogenetic Analyses. Diversity 2021, 13, 517. [Google Scholar] [CrossRef]
- Cui, B.K.; Jia, B.S. Wood-rotting fungi in eastern China 7. Polypores from Huangshan Mountain, Anhui Province. Mycosystema 2011, 30, 529–535. [Google Scholar] [CrossRef]
- Cui, B.K.; Yu, C.J. Fungal flora and population structure of polypores in the Great Hingan Mountains. Acta Ecol. Sin. 2011, 31, 3700–3709. [Google Scholar]
- Cui, B.K.; Dai, Y.C. Wood-decaying fungi in eastern Himalayas 3. Polypores from Laojunshan Mountains, Yunnan Province. Mycosystema 2012, 31, 486–492. [Google Scholar]
- He, X.; Chen, J.Z.; Zhao, C.L. Diversity of wood-decaying fungi in Haikou Forestry Farm, Yunnan Province, P.R. China. Stud. Fungi 2021, 6, 365–377. [Google Scholar] [CrossRef]
- Yuan, H.S.; Dai, Y.C. Polypores from northern and central Yunnan Province, Southwestern China. Sydowia 2008, 60, 147–159. [Google Scholar]
- Wu, Z.Q.; Shen, S.; Luo, K.Y.; Wang, Z.H.; Zhao, C.L. Morphological and molecular identification of a new species of Atraporiella (Polyporales, Basidiomycota) in China. Phytotaxa 2017, 332, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Ma, X.; Xu, T.M.; Zhao, C.L. Phlebia ailaoshanensis sp. nov. (Polyporales, Basidiomycota) evidenced by morphological characters and phylogenetic analyses. Phytotaxa 2018, 373, 184–196. [Google Scholar] [CrossRef]
- Wu, Z.Q.; Xu, T.M.; Shen, S.; Liu, X.F.; Luo, K.Y.; Zhoa, C.L. Elaphroporia ailaoshanensis gen. et sp nov in Polyporales (Basidiomycota). MycoKeys 2018, 29, 81–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.F.; Shen, S.; Zhao, C.L. Morphological and molecular identification of a new species of Eichleriella (Auriculariales, Basidiomycota) in China. Phytotaxa 2019, 404, 245–254. [Google Scholar] [CrossRef]
- Luo, K.Y.; Ma, X.; Zhao, C.L. Neofavolus yunnanensis sp. nov. (Polyporales, Basidiomycota) from China: Evidence from morphology and DNA sequence data. Phytotaxa 2019, 408, 109–116. [Google Scholar] [CrossRef]
- Xu, T.M.; Liu, X.F.; Chen, Y.H.; Zhao, C.L. Rhomboidia wuliangshanensis gen. et sp. nov. from southwestern China. Mycotaxon 2019, 134, 649–662. [Google Scholar] [CrossRef]
- Chen, J.Z.; Zhao, C.L. Morphological and molecular identification of four new resupinate species of Lyomyces (Hymenochaetales) from southern China. MycoKeys 2020, 65, 101–118. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.Z.; Yuan, Z.Y.; Han, C.Z.; Zhao, C.L. Morphological and molecular identification of a new species of Cinereomyces (Polyporales, Basidiomycota) in southern China. Phytotaxa 2020, 459, 51–60. [Google Scholar] [CrossRef]
- Gu, Z.R.; Zhao, C.L. The hidden wood-decaying fungal diversity: Rhizochaete from east Asia. Diversity 2021, 3, 503. [Google Scholar] [CrossRef]
- Luo, K.Y.; Qu, M.H.; Zhao, C.L. Additions to the knowledge of corticioid Xylodon (Schizoporaceae, Hymenochaetales): Introducing three new Xylodon species from southern China. Diversity 2021, 13, 581. [Google Scholar] [CrossRef]
- Wang, X.H.; Das, K.; Bera, I.; Chen, Y.H.; Bhatt, R.P.; Ghosh, A.; Hembrom, M.E.; Hofstetter, V.; Parihar, A.; Vizzini, A.; et al. Fungal Biodiversity Profiles 81–90. Cryptogam. Mycol. 2019, 40, 57–95. [Google Scholar] [CrossRef]
- Zhao, C.L.; Ma, X. Perenniporia mopanshanensis sp. nov. from China. Mycotaxon 2019, 134, 125–137. [Google Scholar] [CrossRef]
- Chen, L.; Shi, Z.J.; Wu, C.H.; Zhao, C.L. Gloeodontia yunnanensis sp. nov. (Russulales, Basidiomycota) from China, evidenced by morphological characters and phylogenetic analyses. Phytotaxa 2020, 432, 111–118. [Google Scholar] [CrossRef]
- Guan, Q.X.; Zhao, T.J.; Zhao, C.L. Morphological characters and phylogenetic analyses reveal two new species of Peniophorella from southern China. Mycol. Prog. 2020, 19, 397–404. [Google Scholar] [CrossRef]
- Cui, B.K.; Wei, Y.L.; Dai, Y.C. Polypores from Zijin Mountain, Jiangsu Province. Mycosystema 2006, 25, 9–14. [Google Scholar]
- Wei, Y.L. Species diversity, fungal flora and distribution characters of polypores in Changbai Mountains, China. Chin. J. Appl. Ecol. 2011, 22, 2711–2717. [Google Scholar]
- Gafforov, Y.; Ordynets, A.; Langer, E.; Yarasheva, M.; de Mello Gugliotta, A.; Schigel, D.; Pecoraro, L.; Zhou, Y.; Cai, L.; Zhou, L.W. Species diversity with comprehensive annotations of wood-inhabiting poroid and corticioid fungi in Uzbekistan. Front. Microbiol. 2020, 11, 598321. [Google Scholar] [CrossRef]
- Dai, Y.C.; Zhou, L.W.; Yang, Z.L.; Wen, H.A.; Bau, T.; Li, T.H. A revised checklist of edible fungi in China. Mycosystema 2010, 29, 1–21. [Google Scholar] [CrossRef]
- Dai, Y.C.; Yang, Z.L. A revised checklist of medicinal fungi in China. Mycosystema 2008, 27, 801–824. [Google Scholar] [CrossRef]
- Mao, X.L. The Economic Fungi in China; Science Press: Beijing, China, 1998. [Google Scholar]
- Ying, J.Z.; Mao, X.L.; Ma, Q.M. Atlas of Chinese Medicinal Fungi; Science Press: Beijing, China, 1987. [Google Scholar]
- Park, W.H.; Lee, H.D. Illustrated Book of Korean Medicinal Mushrooms. Kyo-Hak Publishing Co., Ltd.: Seoul, Korea, 1999. [Google Scholar]
- Saoudi, O.; Ghaouar, N. Biocatalytic characterization of free and immobilized laccase from Trametes versicolor in its activation zone. Biol. Macromolocules 2019, 128, 681–691. [Google Scholar] [CrossRef]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C. Pathogenic wood-decaying fungi on woody plants in China. Mycosystema 2012, 31, 493–509. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspe, O.; Kakishima, M.; Sanchez-Ramırez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.C. Illustrations of Pathogenic Wood-Decaying Fungi in China; Science Press: Beijing, China, 2005. [Google Scholar]
- Dai, Y.C. Illustrations of Wood-Decaying Fungi on Stored Wood or Structural Timber in China; Science Press: Beijing, China, 2009. [Google Scholar]
- Dai, Y.C.; Cui, B.K. Biodiversity of Wood-Inhabiting Fungi in Hainan; Science Press: Beijing, China, 2010. [Google Scholar]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central China Farmer’s Publishing House: Zhengzhou, China, 2015. [Google Scholar]
- Liu, T.Z.; Li, G.L. Atlas of Macrofungi in Saihanwula National Nature Reserve of Inner Mongolia; Inner Mongolia Science and Technology Press: Chifeng, Mongolia, 2019. [Google Scholar]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol Spectr. 2017, 5, 4. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Specimen No. | GenBank Accession No. | References | Country |
|---|---|---|---|---|
| ITS | ||||
| Abundisporus quercicola | CLZhao 10238 | MT159978 | Present study | China, Yunnan |
| A. quercicola | Dai 3084 | KC415907 | [19] | China, Yunnan |
| Agaricus campestris | MA Fungi 80998 | NR_151745 | Unpublished | USA, Bethesda |
| Aleurodiscus botryosus | CBS 336.66 | MH858812 | [20] | Netherlands, Upsalalaan |
| A. botryosus | CLZhao 4333 | MK343481 | Present study | China, Yunnan |
| A. cerussatus | CLZhao 4715 | MK404310 | Present study | China, Yunnan |
| A. cerussatus | SPG 3004 | MT831058 | [19] | Argentina, Esquel |
| A. isabellinus | CLZhao 4405 | MK343482 | Present study | China, Yunnan |
| A. isabellinus | He 5294 | MH109053 | [21] | China, Beijing |
| A. mirabilis | CLZhao 4621 | MK343488 | Present study | China, Yunnan |
| A. mirabilis | JMB 2152 | MT568513 | [19] | Argentina, Esquel |
| A. subroseus | CLZhao 4722 | MK343489 | Present study | China, Yunnan |
| A. subroseus | He 4814 | MH109055 | [21] | China, Beijing |
| Antrodia malicola | CLZhao 3832 | MH114612 | Unpublished | China, Yunnan |
| Athelia epiphylla | SFC 20180314-01 | MK992816 | [22] | Korea, Seoul |
| Auricularia asiatica | CLZhao 3652 | MK268844 | Present study | China, Yunnan |
| A. asiatica | OM 13932 | MZ618931 | [19] | China, Beijing |
| A. cornea | CLZhao 4245 | MK343502 | Present study | China, Yunnan |
| A. cornea | PDD 94825 | KX621146 | [23] | Thailand, Chiang Rai |
| A. delicata | CLZhao 4403 | MK343505 | Present study | China, Yunnan |
| A. delicata | MFLU162112 | KX621149 | [23] | Thailand, Chiang Rai |
| A. mesenterica | BRNM 648573 | KP729279 | [24] | China, Beijing |
| Bjerkandera adusta | CLZhao 4310 | MK343508 | Present study | China, Yunnan |
| B. adusta | HHB-12826-Sp | KP134983 | [25] | USA, Worcester |
| Boletus edulis | 375 | EU554664 | [26] | Spain, Cabrils |
| Byssomerulius corium | CLZhao 9318 | MT177289 | Present study | China, Yunnan |
| B. corium | FP-102382 | KP135007 | [25] | USA, Worcester |
| Cantharellus cibarius | CL-67 | MK281467 | [27] | Canada, Prince George |
| Ceriporia lacerata | CLZhao 3693 | MK268856 | Present study | China, Yunnan |
| C. lacerata | FP-55521T | KP135024 | [25] | USA, Worcester |
| Cerrena albocinnamomea | CLZhao 4193 | MK268860 | Present study | China, Yunnan |
| C. albocinnamomea | Dai 12892 | KC485522 | [19] | China, Beijing |
| C. zonata | CLZhao 3502 | MK268838 | Present study | China, Yunnan |
| C. zonata | ICMP 16347 | MW862786 | [19] | New Zealand, Auckland |
| Climacodon pulcherrimus | CBS 130.40 | MH856063 | [20] | Netherlands, Upsalalaan |
| C. pulcherrimus | CLZhao 4379 | MK343524 | Present study | China, Yunnan |
| Corticium roseocarneum | CBS 330.66 | MH858810 | [20] | Netherlands, Upsalalaan |
| C. roseocarneum | CLZhao 3562 | MK268877 | Present study | China, Yunnan |
| C. roseum | CBS 104.52 | MH856944 | [20] | Netherlands, Upsalalaan |
| Crustodontia chrysocreas | CLZhao 4143 | MK269228 | Present study | China, Yunnan |
| C. chrysocreas | KMRB 18041009 | MK120528 | [19] | Korea, Seoul |
| Dacrymyces cyrtosporus | PDD 107980 | NR_148190 | [28] | USA, Bethesda |
| D. flabelliformis | PDD 76696 | NR_166790 | [29] | USA, Bethesda |
| D. stillatus | UPS F-939814 | MN595677 | [30] | Sweden, Uppsala |
| Daedaleopsis confragosa | CLZhao 4260 | MK343522 | Present study | China, Yunnan |
| D. confragosa | Cui 2883 | KU900526 | [19] | China, Beijing |
| Datronia mollis | CLZhao 4429 | MK343523 | Present study | China, Yunnan |
| D. mollis | KUC 20121123-22 | KJ668554 | [19] | Korea, Seoul |
| D. stereoides | CLZhao 9937 | MT193539 | Present study | China, Yunnan |
| D. stereoides | Cui 8132 | JX559270 | [19] | China, Beijing |
| Earliella scabrosa | CLZhao 3722 | MK268896 | Present study | China, Yunnan |
| E. scabrosa | Cui 6236 | KC867366 | [19] | China, Beijing |
| Fibroporia albicans | CLZhao 4416 | MK343526 | Present study | China, Yunnan |
| F. albicans | Dai 10595 | NR_153980 | [31] | China, Beijing |
| Flavodon ambrosius | CLZhao 3341 | MH114652 | Present study | China, Yunnan |
| F. ambrosius | Hulcr 12546 | LC216231 | [32] | USA, Gainesville |
| Fomes fomentarius | CLZhao 4393 | MK343528 | Present study | China, Yunnan |
| F. fomentarius | Cui 8020 | JX290073 | [19] | China, Beijing |
| Ganoderma australe | CLZhao 4351 | MK343538 | Present study | China, Yunnan |
| G. australe | HMAS 86595 | AY884184 | [19] | China, Beijing |
| Geastrum pectinatum | MJ 9030 | KC581962 | [33] | Sweden, Goteborg |
| Gloeodontia yunnanensis | CLZhao 10504 | MN908252 | Present study | China, Yunnan |
| G. yunnanensis | CLZhao 11058 | MN908253 | Present study | China, Yunnan |
| Gloeophyllum sepiarium | CBS 293.51 | MH856865 | [20] | Netherlands, Upsalalaan |
| G. sepiarium | CLZhao 3473 | MK268934 | Present study | China, Yunnan |
| Gomphus clavatus | EL 64/03 | EU118628 | [34] | Sweden, Goteborg |
| Heterobasidion insulare | CLZhao 4071 | MK268945 | Present study | China, Yunnan |
| H. insulare | Dai 13933 | MT146489 | [35] | China, Beijing |
| Hexagonia glabra | CLZhao 3589 | MK268955 | Present study | China, Yunnan |
| H. glabra | Cui 16796 | MK192431 | [36] | China, Beijing |
| H. tenuis | CLZhao 3485 | MK268956 | Present study | China, Yunnan |
| H. tenuis | Cui 8468 | JX559277 | [37] | China, Beijing |
| Hymenochaete innexa | CLZhao 4433 | MK343546 | Present study | China, Yunnan |
| H. innexa | OUT 855 | MT596387 | [19] | Italy, Torino |
| H. xerantica | CLZhao 2789 | MK343628 | Unpublished | China, Yunnan |
| Hyphoderma moniliforme | CLZhao 10176 | MT246965 | Present study | China, Yunnan |
| H. moniliforme | Wu 0211-46 | KC928284 | [38] | China, Taiwan |
| H. mutatum | CBS 291.36 | MH855803 | [20] | Netherlands, Upsalalaan |
| H. mutatum | CLZhao 10019 | MT246969 | Present study | China, Yunnan |
| H. nudicephalum | CBS 125855 | MH863811 | [20] | Netherlands, Upsalalaan |
| H. nudicephalum | CLZhao 3481 | MK269020 | Present study | China, Yunnan |
| H. transiens | CLZhao 9628 | MT246975 | Present study | China, Yunnan |
| H. transiens | NH 12304 | DQ677504 | [39] | Sweden, Goteborg |
| Hysterangium atlanticum | Fungos 2115 | NR_165211 | [40] | Slovenia, Ljubljana |
| Irpex lacteus | CLZhao 4338 | MK343588 | Present study | China, Yunnan |
| I. lacteus | FD-9 | KP135026 | [25] | USA, Worcester |
| Junghuhnia crustacea | CLZhao 3588 | MK269088 | Present study | China, Yunnan |
| J. crustacea | X 1127 | JN710554 | [41] | Finland, Helsinki |
| J. nitida | CLZhao 3474 | MK269086 | Present study | China, Yunnan |
| J. nitida | KHL 11903 | EU118638 | [34] | Sweden, Goteborg |
| Laxitextum incrustatum | CLZhao 9273 | MT159974 | Present study | China, Yunnan |
| L. incrustatum | STMA 14285 | KT722621 | [42] | Germany, Braunschweig |
| Lenzites betulinus | CLZhao 10151 | MT193546 | Present study | China, Yunnan |
| L. betulinus | Cui 7095 | JX290075 | [19] | China, Beijing |
| Lopharia mirabilis | CLZhao 3518 | MK269105 | Present study | China, Yunnan |
| L. mirabilis | Dai 13722 | MF626346 | [43] | China, Beijing |
| Lyomyces bambusinus | CLZhao 3675 | MN945969 | Present study | China, Yunnan |
| L. bambusinus | CLZhao 4840 | MN945971 | Present study | China, Yunnan |
| L. cremeus | CLZhao 4138 | MN945974 | Present study | China, Yunnan |
| L. cremeus | CLZhao 8295 | MN945972 | Present study | China, Yunnan |
| L. macrosporus | CLZhao 4516 | MN945977 | Present study | China, Yunnan |
| L. macrosporus | CLZhao 8605 | MN945975 | Present study | China, Yunnan |
| L. wuliangshanensis | CLZhao 4475 | MN945983 | Present study | China, Yunnan |
| L. wuliangshanensisd | CLZhao 4108 | MN945980 | Present study | China, Yunnan |
| Megasporia ellipsoidea | CLZhao 10360 | MT193554 | Present study | China, Yunnan |
| M. ellipsoidea | Cui 5222 | JQ314367 | [19] | China, Beijing |
| M. major | CLZhao 10363 | MT193555 | Present study | China, Yunnan |
| M. major | Yuan 1183 | JQ314365 | [19] | China, Beijing |
| Megasporoporiella pseudocavernulosa | CLZhao 9339 | MK894044 | Present study | China, Yunnan |
| M. pseudocavernulosa | Cui 11106 | KX900657 | [19] | China, Beijing |
| M. subcavernulosa | CLZhao 4442 | MK343599 | Present study | China, Yunnan |
| M. subcavernulosa | Cui 10050 | JQ314357 | [19] | China, Beijing |
| Microporus vernicipes | CLZhao 3672 | MK269132 | Present study | China, Yunnan |
| M. vernicipes | Dai 7252 | KX880619 | [19] | China, Beijing |
| M. xanthopus | CLZhao 4280 | MT159985 | Present study | China, Yunnan |
| M. xanthopus | Dai 12076 | KX880620 | [19] | China, Beijing |
| Neodatronia sinensis | CLZhao 9471 | MT193563 | Present study | China, Yunnan |
| N. sinensis | Cui 9949 | KX900663 | [19] | China, Beijing |
| Oxychaete cervinogilva | CLZhao 3462 | MK269143 | Present study | China, Yunnan |
| O. cervinogilva | Schigel 5216 | KX752596 | [44] | Finland, Helsinki |
| Peniophora cinerea | 0911 TES27A5 | LN808982 | [19] | Spain, Sevilla |
| P. cinerea | CLZhao 9254 | MT159975 | Present study | China, Yunnan |
| P. incarnata | CBS 430.72 | MH860518 | [20] | Netherlands, Upsalalaan |
| P. incarnata | CLZhao 9859 | MT159976 | Present study | China, Yunnan |
| P. pesudoversicolor | CLZhao 9415 | MT159977 | Present study | China, Yunnan |
| P. pseudoversicolor | CBS 338.66 | MH858814 | [20] | Netherlands, Upsalalaan |
| Peniophorella fissurata | CLZhao 11412 | MN864261 | Present study | China, Yunnan |
| P. fissurata | CLZhao 4539 | MN864259 | Present study | China, Yunnan |
| P. pubera | CLZhao 9336 | MK894055 | Present study | China, Yunnan |
| P. pubera | NH 10380 | DQ647504 | [45] | Sweden, Goteborg |
| P. rude | CLZhao 3360 | MH114830 | Present study | China, Yunnan |
| P. rude | Wu 9307-39 | DQ647499 | [45] | Sweden, Goteborg |
| Perenniporia luteola | CLZhao 10334 | MT193564 | Present study | China, Yunnan |
| P. luteola | Dai 13094 | KX900679 | [19] | China, Beijing |
| P. mopanshanensis | CLZhao 4677 | MK404422 | Present study | China, Yunnan |
| P. mopanshanensis | CLZhao 5145 | MH784912 | Present study | China, Yunnan |
| Phaeophlebiopsis peniophoroides | CLZhao 3637 | MK269159 | Present study | China, Yunnan |
| P. peniophoroides | FP-150577 | KP135417 | [25] | USA, Worcester |
| Phallus impudicus | KHTW 08001 | MG678529 | [46] | Brazil, Manaus |
| Phanerochaete angustocystidiata | CLZhao 9280 | MT177305 | Present study | China, Yunnan |
| P. angustocystidiata | KUC20121102-15 | KJ668492 | [19] | Korea, Seoul |
| P. australis | CLZhao 3571 | MK269240 | Present study | China, Yunnan |
| P. australis | HHB-7105-Sp | KP135081 | [25] | USA, Worcester |
| P. concrescens | CLZhao 4367 | MK404430 | Present study | China, Yunnan |
| P. concrescens | Spirin 6111 | KP994352 | [47] | Russia, Saint-Petersburg |
| P. sanguineocarnosa | CLZhao 4447 | MK343613 | Present study | China, Yunnan |
| P. sanguineocarnosa | HHB-2189 | KP135124 | [25] | USA, Worcester |
| P. sordida | CLZhao 3407 | MK269174 | Present study | China, Yunnan |
| P. sordida | FD-241 | KP135136 | [25] | USA, Worcester |
| Phlebiopsis castanea | CLZhao 3501 | MK269230 | Present study | China, Yunnan |
| P. castanea | Spirin 5295 | KX752610 | [44] | Finland, Helsinki |
| P. crassa | CLZhao 3339 | MT177325 | Present study | China, Yunnan |
| P. crassa | KKN-86 | KP135394 | [25] | USA, Worcester |
| Rhomboidia wuliangshanensis | CLZhao 4406 | MK860715 | Present study | China, Yunnan |
| R. wuliangshanensis | CLZhao 4411 | MK860716 | Present study | China, Yunnan |
| Rigidoporus pouzarii | CLZhao 4524 | MK404499 | Present study | China, Yunnan |
| R. pouzarii | MJ 123_93 | JQ409464 | [19] | Czech Republic, South Bohemia |
| Russula recondita | LUG 19058 | NR_147635 | [48] | USA, Bethesda |
| Sebacina incrustans | JMP 0084 | EU819442 | [49] | USA, Wisconsin |
| Serpula dendrocalami | CLZhao 3136 | MK863399 | Present study | China, Yunnan |
| S. dendrocalami | CLZhao 3607 | MK863404 | Present study | China, Yunnan |
| Sistotrema brinkmannii | ATCC 36164 | DQ899095 | [19] | USA, Stillwater |
| S. brinkmannii | CLZhao 9324 | MT159987 | Present study | China, Yunnan |
| Sistotremastrum guttuliferum | BHI-F755a | MW488061 | [19] | USA, West Lafayette |
| S. guttuliferum | CLZhao 9771 | MT159990 | Present study | China, Yunnan |
| Steccherinum bourdotii | CLZhao 4297 | MK343648 | Present study | China, Yunnan |
| S. bourdotii | U 107 | MN749593 | [50] | Arabia, Hail |
| S. ochraceum | CLZhao 4154 | MK404504 | Present study | China, Yunnan |
| S. ochraceum | LE-BIN 3984 | MK795064 | [19] | Russia, Saint Petersburg |
| Terana coerulea | CLZhao 9803 | MT177329 | Present study | China, Yunnan |
| T. coerulea | FP-104073 | KP134980 | [25] | USA, Worcester |
| Thelephora terrestris | JMP 0086 | EU819444 | [49] | USA, La Crosse |
| Tinctoporellus epimiltinus | CLZhao 3605 | MK269225 | Present study | China, Yunnan |
| T. epimiltinus | PR1147 | KY948721 | [51] | USA, Massachusetts |
| Trametes ellipsosopia | CLZhao 3774 | MK269226 | Present study | China, Yunnan |
| T. ellipsospora | Cui 6259 | JN048767 | [52] | China, Beijing |
| T. hirsuta | CLZhao 3572 | MK269220 | Present study | China, Yunnan |
| T. hirsuta | TH 158 | MF960866 | [53] | Czech Republic, Brno |
| T. pavonia | CLZhao 3740 | MK269222 | Present study | China, Yunnan |
| T. pavonia | PR2178 | JN164959 | [54] | USA, Worcester |
| T. versicolor | CLZhao 4170 | MK269115 | Present study | China, Yunnan |
| T. versicolor | F-1599 | MN749366 | [19] | USA, West Lafayette |
| Trametopsis cervina | CLZhao 9449 | MW418032 | Present study | China, Yunnan |
| T. cervina | TJV 93_216T | JN165020 | [54] | USA, Worcester |
| Trechispora echinospora | MA-Fungi 82485 | JX392847 | [55] | Spain, Madrid |
| T. nivea | A 1683 | MT458536 | [19] | Greece, Athens |
| T. nivea | CLZhao 10154 | MT159996 | Present study | China, Yunnan |
| Truncospora ochroleuca | CLZhao 4350 | MK343685 | Present study | China, Yunnan |
| T. ochroleuca | BCC 16648 | KP059107 | [19] | Thailand, Khlong |
| Orders | Families | Genera | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Order | Family | Genera | Spp. | % | Family | Genera | Spp. | % | Genera | Spp. | % |
| Polyporales | 9 | 40 | 67 | 70.53 | Polyporaceae | 18 | 28 | 29.47 | Aleurodiscus | 5 | 5.26 |
| Russulales | 5 | 6 | 12 | 12.63 | Phanerochaetaceae | 6 | 11 | 11.58 | Phanerochaete | 5 | 5.26 |
| Hymenochaetales | 2 | 3 | 6 | 6.32 | Steccherinaceae | 5 | 8 | 8.42 | Hyphoderma, | 4 | 4.21 |
| Auriculariales | 1 | 1 | 3 | 3.16 | Hyphodermataceae | 2 | 8 | 8.42 | Lyomyces | 4 | 4.21 |
| Boletales | 2 | 4 | 2 | 2.11 | Stereaceae | 2 | 6 | 6.32 | Trametes | 4 | 4.21 |
| Trechisporales | 1 | 2 | 2 | 2.11 | Irpicaceae | 5 | 5 | 5.26 | Auricularia | 3 | 3.16 |
| Cantharellales | 1 | 1 | 1 | 1.05 | Cerrenaceae | 2 | 3 | 3.16 | Peniophora | 3 | 3.16 |
| Corticiales | 1 | 1 | 1 | 1.05 | Hymenochaetaceae | 2 | 3 | 3.16 | Peniophorella | 3 | 3.16 |
| Gloeophyllales | 1 | 1 | 1 | 1.05 | Rickenellaceae | 1 | 3 | 3.16 | Steccherinum | 3 | 3.16 |
| Subtotal | 43 | 75 | 78.95 | Subtotal | 34 | 35.79 | |||||
| Other families (14) | 16 | 20 | 21.05 | Other genera (50) | 61 | 64.21 | |||||
| Total | 23 | 59 | 95 | 100% | 23 | 59 | 95 | 100% | 59 | 95 | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Zhao, C.-L. Diversity of Wood-Decaying Fungi in Wuliangshan Area, Yunnan Province, P.R. China. Diversity 2022, 14, 131. https://doi.org/10.3390/d14020131
He X, Zhao C-L. Diversity of Wood-Decaying Fungi in Wuliangshan Area, Yunnan Province, P.R. China. Diversity. 2022; 14(2):131. https://doi.org/10.3390/d14020131
Chicago/Turabian StyleHe, Xiao, and Chang-Lin Zhao. 2022. "Diversity of Wood-Decaying Fungi in Wuliangshan Area, Yunnan Province, P.R. China" Diversity 14, no. 2: 131. https://doi.org/10.3390/d14020131
APA StyleHe, X., & Zhao, C. -L. (2022). Diversity of Wood-Decaying Fungi in Wuliangshan Area, Yunnan Province, P.R. China. Diversity, 14(2), 131. https://doi.org/10.3390/d14020131