
Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Examination
2.2. DNA Amplification and Sequencing
2.3. Phylogenetic Analyses
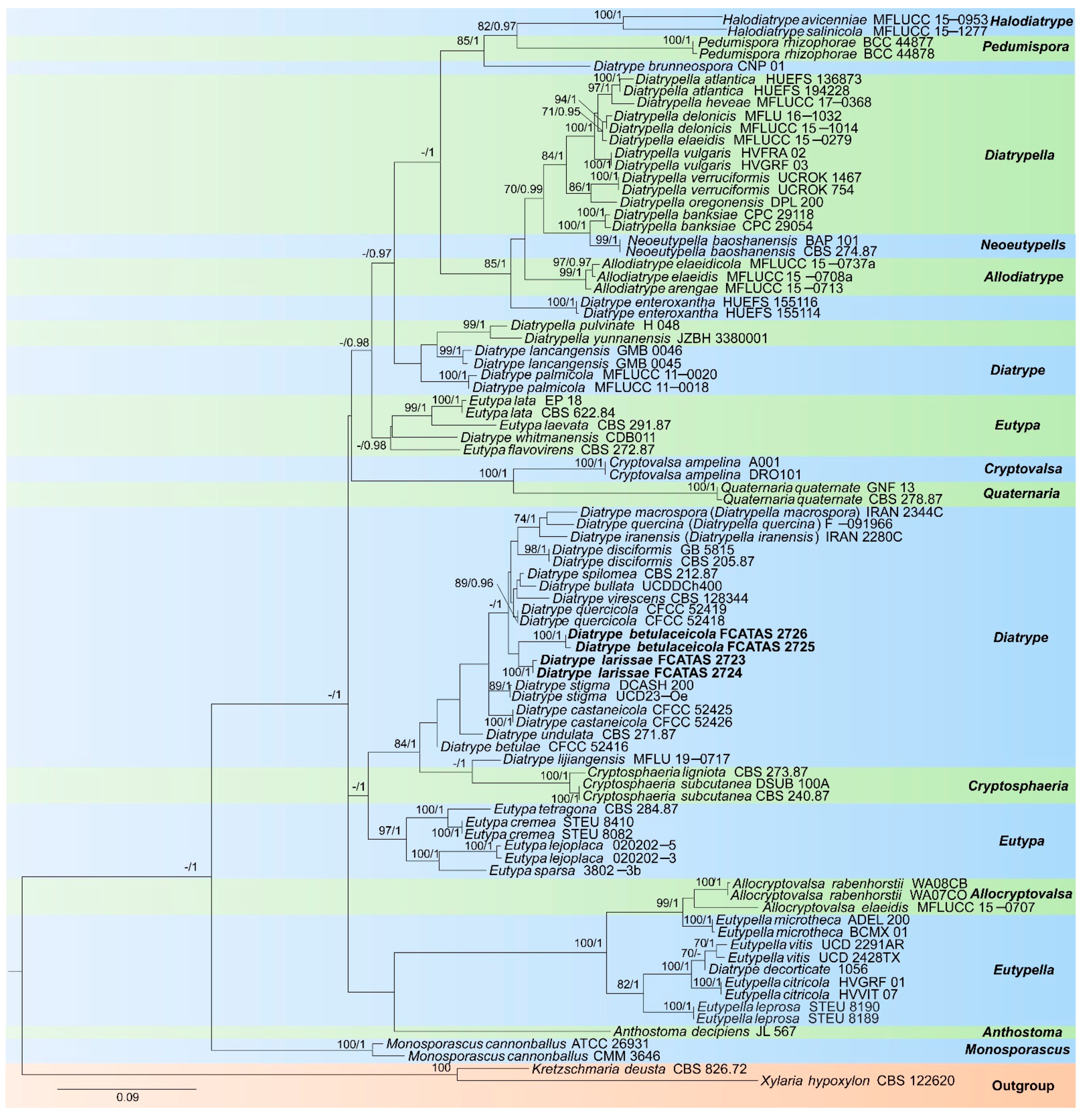
3. Results
3.1. Molecular Phylogeny
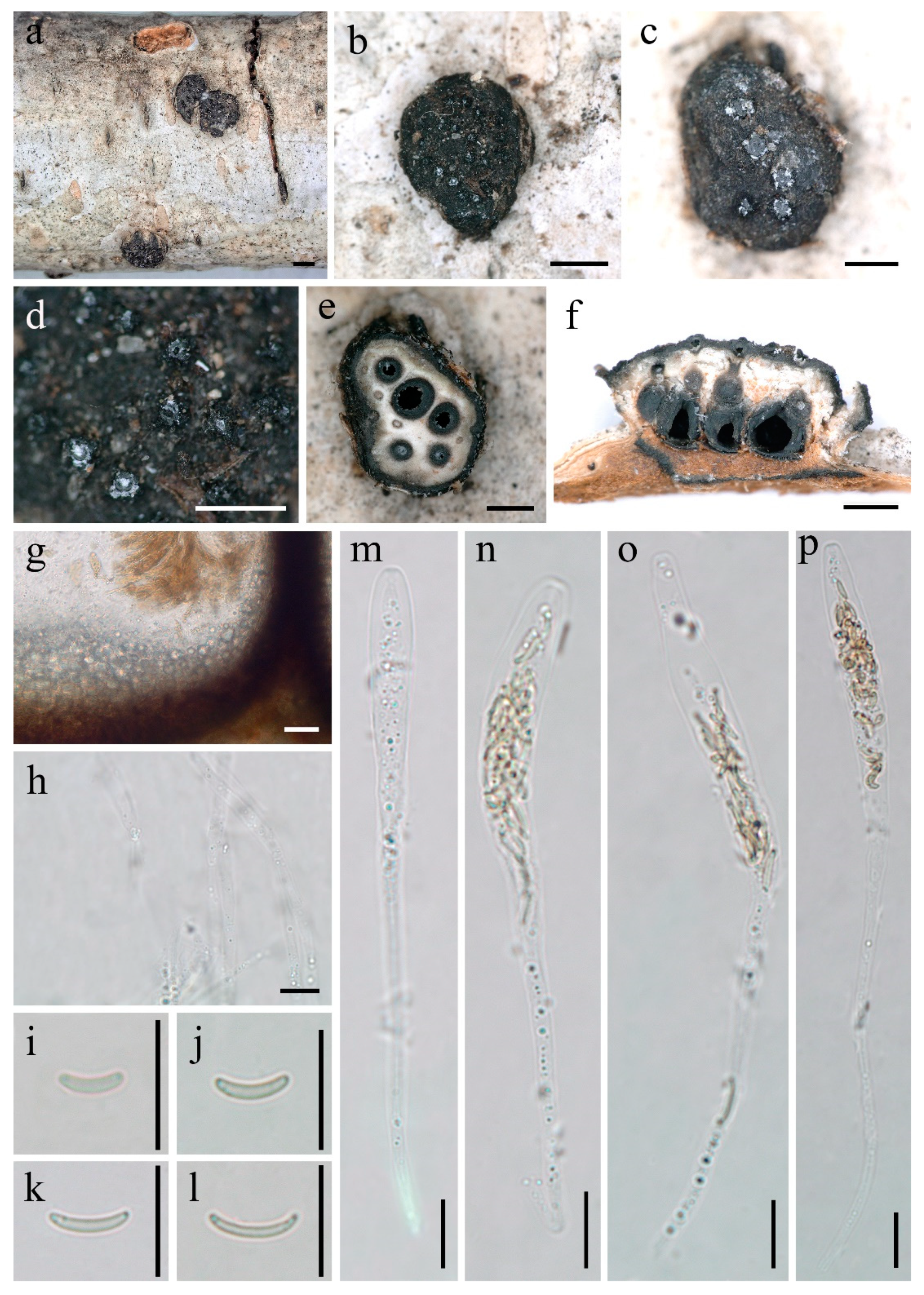
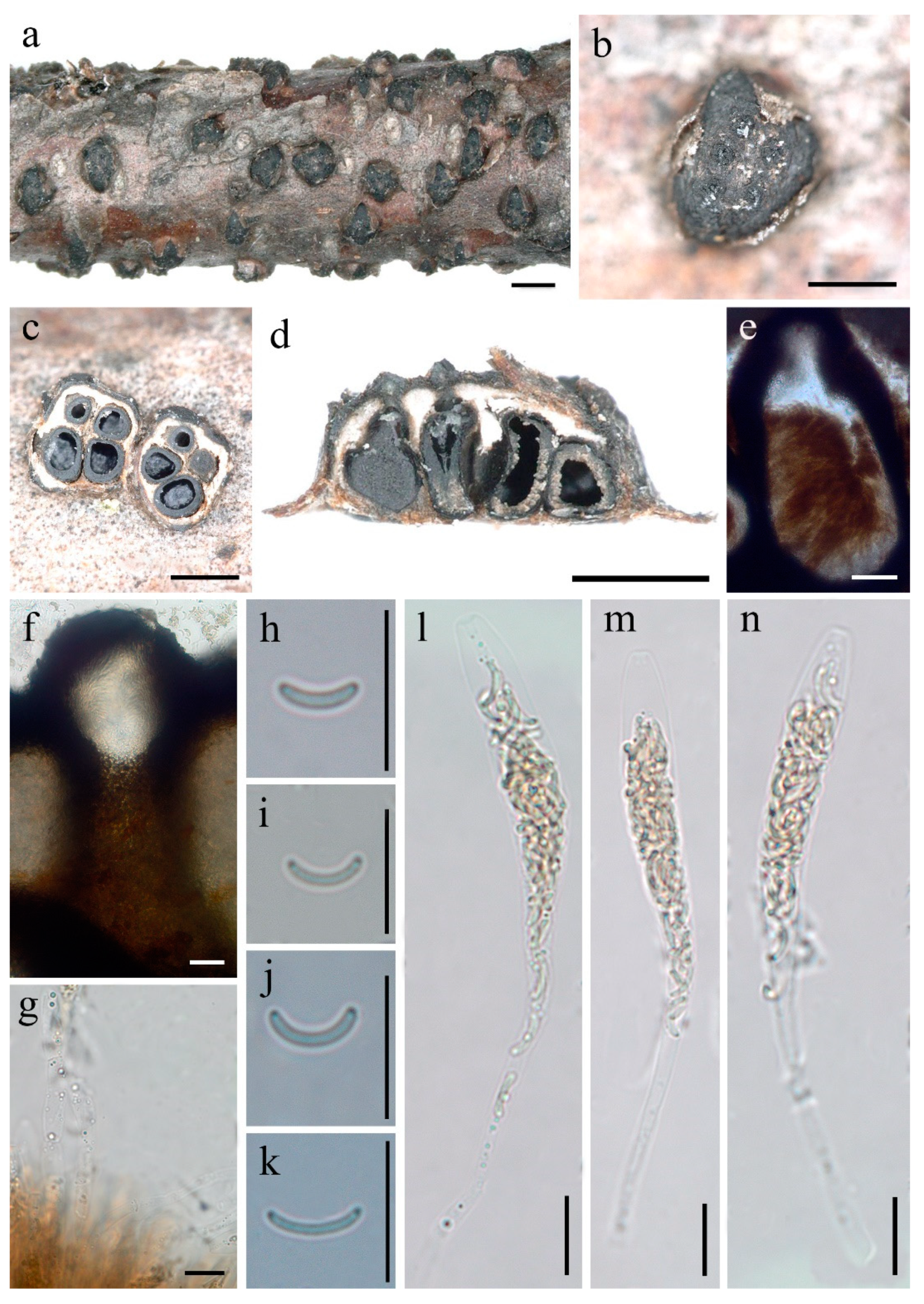
3.2. Taxonomy
4. Discussion
| Key to the speices of Diatrype known from China |
| 1. Sexual morph not known ............................................................................................................. 2 |
| 1. Sexual morph present ................................................................................................................... 3 |
| 2. Conidia 10–13 × 1–2 μm ................................................................................................. D. betulae |
| 2. Conidia 4–6 × 1–1.5 μm .......................................................................................... D. castaneicola |
| 3. Stromata solitary, pustulate, circular or elliptic ........................................................................ 4 |
| 3. Stromata widely and indefinitely effuse .................................................................................. 13 |
| 4. Ascus with more than 8 spores ................................................................................................... 5 |
| 4. Ascus with 8 spores ...................................................................................................................... 7 |
| 5. Ascospores 17–27 × 4–6 μm ...................................................................................... D. quercicola |
| 5. Ascospores length less than 10 μm ............................................................................................. 6 |
| 6. Perithecia less than 6 ...................................................................................................... D. larissae |
| 6. Perithecia more than 6 ........................................................................................... D. betulaceicola |
| 7. Ascus length more than 30 μm .................................................................................................... 8 |
| 7. Ascus length less than 30 μm ..................................................................................................... 11 |
| 8. Ascospores 6–8 µm long, size of asci more than 65 μm ....................................... D. lijiangensis |
| 8. Ascospores 8–15 µm long, size of asci less than 65 μm ............................................................ 9 |
| 9. Ascospores brownish ............................................................................................ D. albopruinosa |
| 9. Ascospores subhyaline to light yellow, or light yellow ......................................................... 10 |
| 10. Stromata pulvinate, hemispherical or forming linear stripes .......................... D. oregonensis |
| 10. Stromata circular-polygonal or irregular ......................................................... D. macowaniana |
| 11. Ascospores 4–6 µm long ........................................................................................... D. macounii |
| 11. Ascospores 7–9 µm long ........................................................................................................... 12 |
| 12. Stromata 1–2 × 1–1.5 mm ............................................................................................ D. acericola |
| 12. Stromata 2–7 mm diam ................................................................................................. D. bullata |
| 13. Ascospores 11–19 µm long ................................................................................. D. lancangensis |
| 13. Ascospores 4–9 µm long ........................................................................................................... 14 |
| 14. Ascospores yellowish .......................................................................................... D. subundulata |
| 14. Ascospores hyaline ................................................................................................................... 15 |
| 15. Asci 20–25 μm long, Stromata chocolate-brown ............................................. D. hypoxyloides |
| 15. Asci 25–30 μm long, Stromata different shades of brownish tinges or black .................... 16 |
| 16. Stromata black, perithecia more than 200 µm diam ............................................... D. spilomea |
| 16. Stromata various shades of brownish tinges, perithecia less than 200 µm diam.....D. stigma |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nitschke, T.R.J. Grundlage eines Systems der Pyrenomyceten. In Verhandlungen des Naturhistorischen Vereins der Preussischen Rheinlande, Westfalens und des Regierungsbezirks Osnabrück; 1905; Volume 62, pp. 70–77. Available online: https://www.forgottenbooks.com/en/books/VerhandlungendesNaturhistorischenVereinsderPreussischenRheinlandeWestfalensundde_11028671 (accessed on 20 January 2022).
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Acero, F.J.; González, V.; Ballesteros, J.S.; Rubio, V.; Checa, J.; Bills, G.F.; Salazar, O.; Platas, G.; Peláez, F. Molecular phylogenetic studies on the Diatrypaceae based on rDNA-ITS sequences. Mycologia 2004, 96, 249–259. [Google Scholar] [CrossRef]
- Trouillas, F.P.; Sosnowski, M.R.; Gubler, W.D. Two new species of Diatrypaceae from coastal wattle in Coorong National Park, South Australia. Mycosphere 2010, 1, 183–188. [Google Scholar]
- Trouillas, F.P.; Urbez-Torres, J.R.; Gubler, W.D. Diversity of diatrypaceous fungi associated with grapevine canker diseases in California. Mycologia 2010, 102, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Moyo, P.; Damm, U.; Mostert, L.; Halleen, F. Eutypa, Eutypella, and Cryptovalsa Species (Diatrypaceae) associated with Prunus species in South Africa. Plant Disease 2018, 102, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, M.; Asgari, B.; Hemmati, R. Two new species of Eutypella and a new combination in the genus Peroneutypa (Diatrypaceae). Mycol. Progress 2019, 18, 1057–1069. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Wanasinghe, D.N.; Devadatha, B.; Abeywickrama, P.; Jones, E.B.G.; Chomnunti, P.; Sarma, V.V.; Hyde, K.D.; Lumyong, S.; McKenzie, E.H.C. Modern taxonomic approaches to identifying diatrypaceous fungi from marine habitats, with a Novel Genus Halocryptovalsa Dayarathne & K.D.Hyde. Gen. Nov. Cryptogam. Mycol. 2020, 41, 21–67. [Google Scholar] [CrossRef]
- Konta, S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; McKenzie, E.H.C.; Stadler, M.; Boonmee, S.; Phookamsak, R.; Jayawardena, R.S.; Senwanna, C.; Hyde, K.D.; et al. A new genus Allodiatrype, fve new species and a new host record of diatrypaceous fungi from palms (Arecaceae). Mycosphere 2020, 11, 239–268. [Google Scholar] [CrossRef]
- Long, S.H.; Liu, L.L.; Pi, Y.H.; Wu, Y.P.; Lin, Y.; Zhang, X.; Long, Q.D.; Kang, Y.Q.; Kang, J.C.; Wijayawardene, N.N.; et al. New contributions to Diatrypaceae from karst areas in China. Mycokeys 2021, 83, 1–37. [Google Scholar] [CrossRef]
- Doidge, E.M. The South African fungi and lichens to the end of 1945. Bothalia 1950, 5, 1–1094. [Google Scholar]
- Munk, A. Danish pyrenomycetes. A preliminary flora. Dan. Bot. Arkiv 1957, 17, 1–491. [Google Scholar]
- Conners, I.L. An Annotated Index of Plant Diseases in Canada and Fungi Tecorded on Plants in Alaska; Canadian Department of Agriculture: Ottawa, ON, Canada, 1967.
- Rappaz, F. Taxonomie et nomenclature des Diatrypaceae à asques octospores. Mycol. Helv. 1987, 2, 285–648. [Google Scholar]
- Mulenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A Preliminary Checklist of Micromycetes in Poland. Kraków: W. Szafer Institute of Botany; Polish Academy of Sciences: Warsaw, Poland, 2008. [Google Scholar]
- Vasilyeva, L.N.; Ma, H.X. Diatrypaceous fungi in north-eastern China. 1. Cryptosphaeria and Diatrype. Phytotaxa 2014, 186, 261–270. [Google Scholar] [CrossRef]
- Peng, M.K.; Zhang, B.; Qu, Z.; Li, Y.; Ma, H.X. New record genus and a new species of Allodiatrype from China based on morphological and molecular characters. Phytotaxa 2021, 500, 275–284. [Google Scholar] [CrossRef]
- Glawe, D.A.; Rogers, J.D. Diatrypaceae in the Pacific Northwest. Mycotaxon 1984, 20, 401–460. [Google Scholar]
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. I. Diatrype Fr. (Diatrypaceae). Fungal Divers. 2004, 17, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Vasilyeva, L.N.; Stephenson, S.L. The genus Diatrype (ascomycota, diatrypaceae) in arkansas and texas (USA). Mycotaxon 2009, 107, 307–313. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Dissanayake, L.S.; Wijayawardene, N.N.; Dayarathne, M.C.; Samarakoon, M.C.; Dai, D.Q.; Hyde, K.D.; Kang, J.C. Paraeutypella guizhouensis gen. et sp. nov. and Diatrypella longiasca sp. nov. (Diatrypaceae) from China. Biodivers. Data J. 2021, 9, e63864. [Google Scholar] [CrossRef]
- Zhu, H.; Pan, M.; Wijayawardene, N.N.; Jiang, N.; Ma, R.; Dai, D.; Tian, C.; Fan, X. The Hidden Diversity of Diatrypaceous Fungi in China. Front. Microbiol. 2021, 12, 646262. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, E.; Fournier, J.; Per, D.; Luangsaard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium Are Nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop 2010 (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tiffany, L.H.; Gilman, J.C. Iowa ascomycetes 4: Diatrypaceae. Iowa State J. Sci. 1965, 40, 121–161. [Google Scholar]
- Trouillas, F.P.; Wayne, M.P.; Sosnowski, M.R.; Huang, R.; Peduto, F.; Loschiavo, A.; Savocchia, S.; Scott, E.S.; Gubler, W.D. Taxonomy and DNA phylogeny of Diatrypaceae associated with Vitis vinifera and other woody plants in Australia. Fungal Divers. 2011, 49, 203–223. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, M.; Hemmati, R.; Vasilyeva, L.N.; Trouillas, F.P. A new species and a new record of Diatrypaceae from Iran. Mycosphere 2015, 6, 60–68. [Google Scholar] [CrossRef]
- Mehrabi, M.; Hemmati, R.; Vasilyeva, L.N.; Trouillas, F.P. Diatrypella macrospora sp. nov. and new records of diatrypaceous fungi from Iran. Phytotaxa 2016, 252, 43–55. [Google Scholar] [CrossRef]
- De Almeida, D.A.C.; Gusmão, L.F.P.; Miller, A.N. Taxonomy and molecular phylogeny of Diatrypaceae (Ascomycota, Xylariales) species from the Brazilian semi-arid region, including four new species. Mycol. Prog. 2016, 15, 1–27. [Google Scholar] [CrossRef]
- Shang, Q.J.; Hyde, K.D.; Phookamsak, R.; Doilom, M.; Bhat, D.J.; Maharachchikumbura, S.S.; Promputtha, I. Diatrypella tectonae and Peroneutypa mackenziei spp. nov. (Diatrypaceae) from northern Thailand. Mycol. Prog. 2017, 16, 463–476. [Google Scholar] [CrossRef]
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. II. Cryptovalsa Ces. et De Not. And Diatrypaceae (Ces. et De Not.) Nitschke (Diatrypaceae). Fungal Divers. 2005, 19, 189–200. [Google Scholar]
- Liu, J.K.; Hyde, K.D.; Gareth, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.; Mckenzie, E.H.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Host/Substrate | Origin | GenBank Accession Numbers | |
|---|---|---|---|---|---|
| ITS | TUB2 | ||||
| Allocryptovalsa elaeidis | MFLUCC 15-0707 | Elaeis guineensis | Thailand | MN308410 | MN340296 |
| A. rabenhorstii | WA07CO | Vitis vinifera | Australia | HQ692620 | HQ692522 |
| A. rabenhorstii | WA08CB | Vitis vinifera | Australia | HQ692619 | HQ692523 |
| Allodiatrype arengaeTS | MFLUCC 15-0713 | Arenga pinnata | Thailand | MN308411 | MN340297 |
| A. elaeidicola | MFLUCC 15-0737a | Elaeis guineensis | Thailand | MN308415 | MN340299 |
| A. elaeidis | MFLUCC 15-0708a | Elaeis guineensis | Thailand | MN308412 | MN340298 |
| Anthostoma decipiensTS | JL567 | Vitis vinifera | Spain | JN975370 | JN975407 |
| Cryptosphaeria ligniota | CBS 273.87 | Populus tremula | Switzerland | KT425233 | KT425168 |
| C. subcutanea | CBS 240.87 | NA | Norway | KT425232 | KT425167 |
| C. subcutanea | DSUB100A | NA | Norway | KT425189 | KT425124 |
| Cryptovalsa ampelina | A001 | NA | Australia | GQ293901 | GQ293972 |
| C. ampelina | DRO101 | NA | America | GQ293902 | GQ293982 |
| Diatrype betulae | CFCC 52416 | Betula davurica | China | MW632943 | NA |
| D. betulaceicola TS | FCATAS 2725 | Betula sp. | China | OM040386 | OM240966 |
| D. betulaceicola TS | FCATAS 2726 | Betula sp. | China | OM040387 | OM240967 |
| D. decorticata | 1056 | NA | NA | KU320621 | NA |
| D. brunneospora | CNP01 | Acacia longifolia subsp. sophorae | Australia | HM581946 | HQ692478 |
| D. bullata | UCDDCh400 | NA | America | DQ006946 | DQ007002 |
| D. castaneicola | CFCC 52425 | Castanea mollissima | China | MW632941 | NA |
| D. castaneicola | CFCC 52426 | Castanea mollissima | China | MW632942 | NA |
| D. disciformisTS | CBS 205.87 | Fagus sylvatica | Switzerland | AJ302437 | NA |
| D. disciformisTS | GB 5815 | Fagus grandifolia | America | KR605644.1 | KY352434.1 |
| D. enteroxantha | HUEFS155114 | NA | Brazil | KM396617 | KT003700 |
| D. enteroxantha | HUEFS155116 | NA | Brazil | KM396618 | KT022236 |
| D. iranensis (Diatrypellairanensis) | IRAN 2280C | Quercus brantii | Iran | KM245033 | NA |
| D. lancangensis | GMB0045 | NA | China | MW797113 | MW814885 |
| D. lancangensis | GMB0046 | NA | China | MW797114 | MW814886 |
| D. larissae TS | FCATAS 2723 | NA | China | OM040384 | OM240964 |
| D. larissae TS | FCATAS 2724 | NA | China | OM040385 | OM240965 |
| D. lijiangensis | MFLU 19-0717 | NA | China | MK852582 | MK852583 |
| D. macrospora (Diatrypellamacrospora) | IRAN 2344C | Quercus brantii | Iran | KR605648 | NA |
| D. oregonensis | DPL200 | Quercus kelloggii | America | GQ293940 | GQ293999 |
| D. palmicola | MFLUCC 11-0018 | Caryota urens | Thailand | KP744438 | NA |
| D. palmicola | MFLUCC 11-0020 | Caryota urens | Thailand | KP744439 | NA |
| D. quercicola | CFCC 52418 | Quercus mongolica | China | MW632938 | NA |
| D. quercicola | CFCC 52419 | Quercus mongolica | China | MW632939 | NA |
| D. quercina (Diatrypellaquercina) | F-091966 | Quercus faginea | Spain | AJ302444 | NA |
| D. spilomea | CBS 212.87 | Acer campestre | Switzerland | AJ302433 | NA |
| D. stigma | DCASH200 | Quercus sp. | America | GQ293947 | GQ294003 |
| D. stigma | UCD23-Oe | NA | NA | JX515704 | JX515670 |
| D. undulata | CBS 271.87 | Betula sp. | Switzerland | AJ302436 | NA |
| D. virescens | CBS 128344 | NA | NA | MH864890 | NA |
| D. whitmanensis | CDB011 | Vitis vinifera | America | GQ293954 | GQ294010 |
| Diatrypella atlantica | HUEFS 136873 | unidentified plant | Brazil | KM396614 | KR259647 |
| D. atlantica | HUEFS 194228 | unidentified plant | Brazil | KM396615 | KR363998 |
| D. banksiae | CPC 29054 | Banksia coccinia | Australia | KY173401 | NA |
| D. banksiae | CPC 29118 | Banksia formosa | Australia | KY173402 | NA |
| D. delonicis | MFLU 16-1032 | Delonix regia | Thailand | MH812995 | MH847791 |
| D. delonicis | MFLUCC 15-1014 | Delonix regia | Thailand | MH812994 | MH847790 |
| D. elaeidis | MFLUCC 15-0279 | Elaeis guineensis | Thailand | MN308417 | MN340300 |
| D. heveae | MFLUCC 17-0368 | Hevea brasiliensis | Thailand | MF959501 | MG334557 |
| D. pulvinata | H048 | Salix alba | Czech Republic | FR715523 | FR715495 |
| D. verruciformisTSQ | UCROK1467 | Quercus agrifolia | America | JX144793 | JX174093 |
| D. verruciformisTSQ | UCROK754 | Quercus agrifolia | America | JX144783 | JX174083 |
| D. vulgaris | HVFRA02 | Fraxinus angustifolia | Australia | HQ692591 | HQ692503 |
| D. vulgaris | HVGRF03 | Citrus paradisi | Australia | HQ692590 | HQ692502 |
| D. yunnanensis | JZBH3380001 | unidentified plant | China | MN653008 | MN887112 |
| Eutypa cremea | STEU 8082 | Vitis vinifera | South Africa | KY111656 | KY111598 |
| E. cremea | STEU 8410 | Prunus armeniaca | South Africa | KY752765 | KY752789 |
| E. flavovirens | CBS 272.87 | Quercus ilex | France | AJ302457 | DQ006959 |
| E. laevata | CBS 291.87 | Salix sp. | Switzerland | HM164737 | HM164771 |
| E. lataTS | EP18 | Vitis vinifera | NSW, Australia | HQ692611 | HQ692501 |
| E. lataTS (Eutypa armeniacae) | CBS 622.84 | Vitis vinifera | Italy | AJ302446 | DQ006964 |
| E. lejoplaca | 020202-3 | Acer pseudoplatanus | Switzerland | AY684238 | AY684197 |
| E. lejoplaca | 020202-5 | Acer pseudoplatanus | Switzerland | AY684221 | AY684196 |
| E. sparsa | 3802-3b | Populus sp. | Switzerland | AY684220 | AY684201 |
| E. tetragona | CBS 284.87 | Sarothamnus scoparius | France | DQ006923 | DQ006960 |
| Eutypella citricola | HVGRF01 | Citrus sinensis | Australia | HQ692589 | HQ692521 |
| E. citricola | HVVIT07 | Vitis vinifera | Australia | HQ692579 | HQ692512 |
| E. leprosa | STEU 8189 | NA | South Africa | MF359637 | MF359672 |
| E. leprosa | STEU 8190 | NA | South Africa | MF359638 | MF359673 |
| E. microtheca | ADEL200 | Ulmus procera | Australia | HQ692559 | HQ692527 |
| E. microtheca | BCMX01 | Vitis vinifera | Mexico | KC405563 | KC405560 |
| E. vitis | UCD2291AR | Vitis vinifera | America | HQ288224 | HQ288303 |
| E. vitis | UCD2428TX | Vitis vinifera | TX, America | FJ790851 | GU294726 |
| Halodiatrype avicenniae | MFLUCC 15-0953 | Avicennia sp. | Thailand | KX573916 | KX573931 |
| H. salinicolaTS | MFLUCC 15-1277 | submerged marine wood | Thailand | KX573915 | KX573932 |
| Kretzschmaria deusta | CBS 826.72 | NA | NA | KU683767 | KU684190 |
| Monosporascus cannonballusTS | ATCC 26931 | NA | NA | FJ430598 | NA |
| M. cannonballusTS | CMM 3646 | Boerhavia sp. | Brazil | JX971617 | NA |
| Neoeutypella baoshanensisTS | BAP101 | Pinus armandii | China | MH822887 | MH822888 |
| N. baoshanensisTS | CBS 274.87 | Ficus carica | France | AJ302460 | NA |
| Pedumispora rhizophoraeTS | BCC44877 | Rhizophora apiculata | Thailand | KJ888853 | NA |
| P. rhizophoraeTS | BCC44878 | Rhizophora apiculata | Thailand | KJ888854 | NA |
| Quaternaria quaternata | GNF13 | Fagus sp. | Iran | KR605645 | KY352464 |
| Q. quaternata | CBS 278.87 | Fagus sulvatica | Switzerland | AJ302469 | NA |
| Xylaria hypoxylon | CBS 122620 | NA | Sweden | AM993141 | KX271279 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Zhang, B.; Qu, Z.; Song, Z.; Pan, X.; Zhao, C.; Ma, H. Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China. Diversity 2022, 14, 149. https://doi.org/10.3390/d14020149
Yang Z, Zhang B, Qu Z, Song Z, Pan X, Zhao C, Ma H. Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China. Diversity. 2022; 14(2):149. https://doi.org/10.3390/d14020149
Chicago/Turabian StyleYang, Zhanen, Bo Zhang, Zhi Qu, Zikun Song, Xiaoyan Pan, Changlin Zhao, and Haixia Ma. 2022. "Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China" Diversity 14, no. 2: 149. https://doi.org/10.3390/d14020149
APA StyleYang, Z., Zhang, B., Qu, Z., Song, Z., Pan, X., Zhao, C., & Ma, H. (2022). Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China. Diversity, 14(2), 149. https://doi.org/10.3390/d14020149