
Ant Diversity Declines with Increasing Elevation along the Udzungwa Mountains, Tanzania

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description
2.2. Ant Sampling
2.3. Data Analysis
3. Results
3.1. Ant Diversity across Three Main Forest Types along the Elevational Gradient
3.2. Sampling Completeness
3.3. Species Diversity Patterns
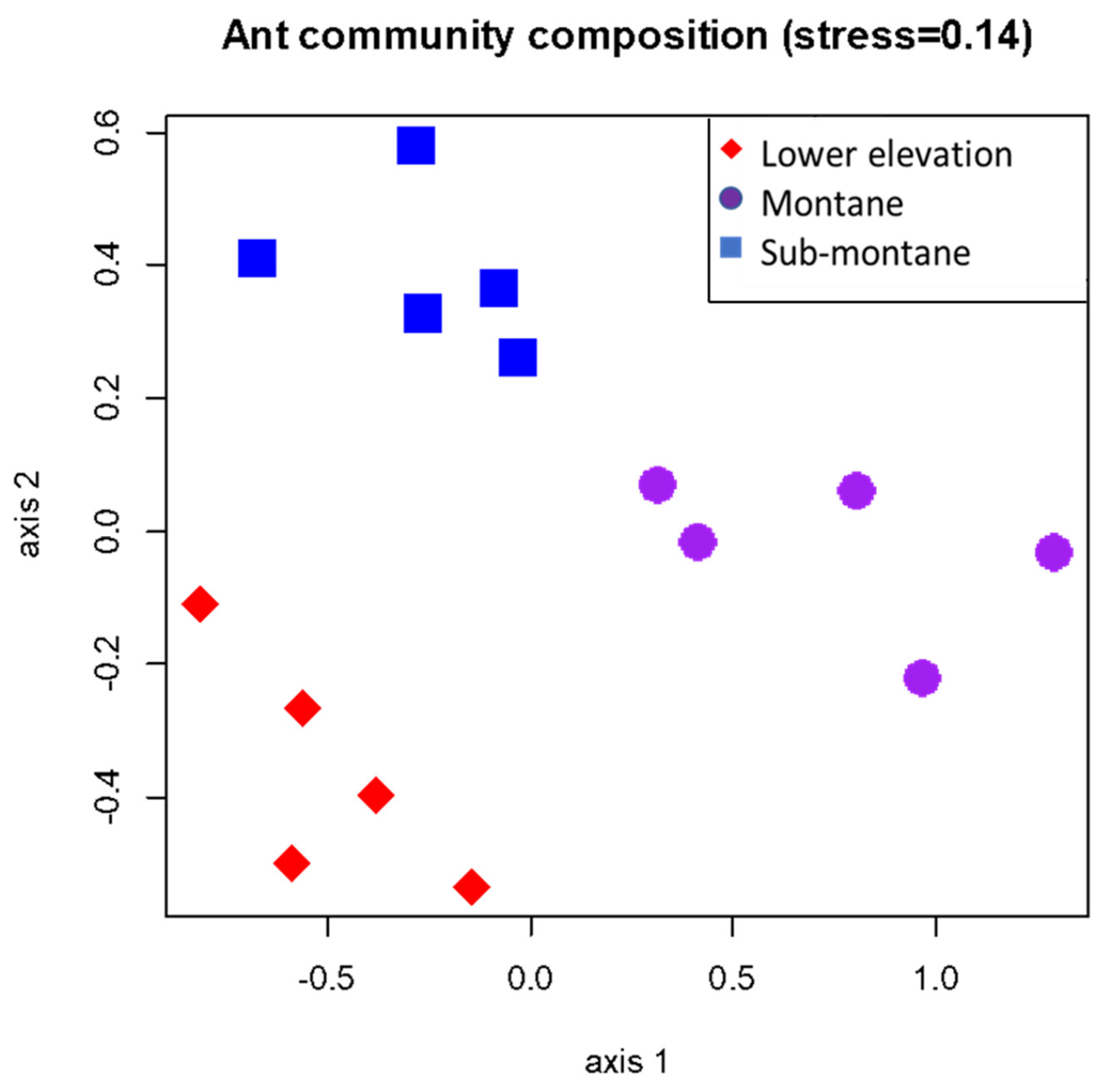
3.4. Ant Assemblage Composition across Gradients
3.5. Indicators Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dickinson, K.J.M.; Mark, A.F.; Lee, W.G. Long-term monitoring of nonforest communities for biological conservation. New Zealand J. Bot. 1992, 30, 163–179. [Google Scholar] [CrossRef]
- Jost, L.; DeVries, P.; Walla, T.; Greeney, H.; Chao, A.; Ricotta, C. Partitioning diversity for conservation analyses. Divers. Distrib. 2010, 16, 65–76. [Google Scholar] [CrossRef]
- Malumbres-Olarte, J.; Crespo, L.; Cardoso, P.; Szuts, T.; Fannes, W.; Pape, T.; Scharff, N. The same but different: Equally megadiverse but taxonomically variant spider communities along an elevational gradient. Acta Oecologica-Int. J. Ecol. 2018, 88, 19–28. [Google Scholar] [CrossRef]
- McCoy, E.D. The distribution of insects along elevational gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Brown, J.H. Mammals on mountainsides: Elevational patterns of diversity. Glob. Ecol. Biogeogr. 2001, 10, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Sanders, N.J.; Moss, J.; Wagner, D. Patterns of ant species richness along elevational gradients in an arid ecosystem. Glob. Ecol. Biogeogr. 2003, 12, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal gradients of biodiversity: Pattern, process, scale, and synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Kaspari, M.; Ward, P.S.; Yuan, M. Energy gradients and the geographic distribution of local ant diversity. Oecologia 2004, 140, 407–413. [Google Scholar] [CrossRef]
- Arnan, X.; Cerda, X.; Retana, J. Ant functional responses along environmental gradients. J. Anim. Ecol. 2014, 83, 1398–1408. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C. The elevational gradient of species richness—A uniform pattern. Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Worm, B. A neutral-metabolic theory of latitudinal biodiversity. Glob. Ecol. Biogeogr. 2016, 25, 630–641. [Google Scholar] [CrossRef]
- Munyai, T.C.; Foord, S.H. Ants on a mountain: Spatial, environmental and habitat associations along an altitudinal transect in a centre of endemism. J. Insect Conserv. 2012, 16, 677–695. [Google Scholar] [CrossRef]
- Munyai, T.C.; Foord, S.H. Temporal Patterns of Ant Diversity across a Mountain with Climatically Contrasting Aspects in the Tropics of Africa. PLoS ONE 2015, 10, e0122035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, M.K.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F.; et al. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and Ecosystem Responses to Elevational Gradients: Processes, Mechanisms, and Insights for Global Change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Bishop, T.R.; Robertson, M.P.; van Rensburg, B.J.; Parr, C.L. Elevation-diversity patterns through space and time: Ant communities of the Maloti-Drakensberg Mountains of southern Africa. J. Biogeogr. 2014, 41, 2256–2268. [Google Scholar] [CrossRef] [Green Version]
- Sanders, N.J.; Lessard, J.P.; Fitzpatrick, M.C.; Dunn, R.R. Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains. Glob. Ecol. Biogeogr. 2007, 16, 640–649. [Google Scholar] [CrossRef]
- Korner, C. The use of ’altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Meier, S.; Soininen, J.; Casamayor, E.O.; Pan, F.Y.; Tang, X.M.; Yang, X.D.; Zhang, Y.L.; Wu, Q.L.; Zhou, J.Z.; et al. Regional and global elevational patterns of microbial species richness and evenness. Ecography 2017, 40, 393–402. [Google Scholar] [CrossRef]
- Chinn, W.G.H.; Chinn, T.J.H. Tracking the snow line: Responses to climate change by New Zealand alpine invertebrates. Arct. Antarct. Alp. Res. 2020, 52, 361–389. [Google Scholar] [CrossRef]
- Bruno, D.; Belmar, O.; Maire, A.; Morel, A.; Dumont, B.; Datry, T. Structural and functional responses of invertebrate communities to climate change and flow regulation in alpine catchments. Glob. Change Biol. 2019, 25, 1612–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunetti, M.; Magoga, G.; Iannella, M.; Biondi, M.; Montagna, M. Phylogeography and species distribution modelling of Cryptocephalus barii (Coleoptera: Chrysomelidae): Is this alpine endemic species close to extinction? Zookeys 2019, 865, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecl, G.T.; Araujo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengard, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, 6332. [Google Scholar] [CrossRef]
- Lovett, J.C.; Marshall, A.R.; Carr, J. Changes in tropical forest vegetation along an altitudinal gradient in the Udzungwa Mountains National Park, Tanzania. Afr. J. Ecol. 2006, 44, 478–490. [Google Scholar] [CrossRef]
- Rovero, F.; Marshall, A.R.; Jones, T.; Perkin, A. The primates of the Udzungwa Mountains: Diversity, ecology and conservation. J. Anthropol. Sci. 2009, 87, 93–126. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Dinesen, L.; Lehmberg, T.; Rahner, M.C.; Fjeldsa, J. Conservation priorities for the forests of the Udzungwa Mountains, Tanzania, based on primates, duikers and birds. Biol. Conserv. 2001, 99, 223–236. [Google Scholar] [CrossRef]
- Rovero, F.; De Luca, D.W. Checklist of mammals of the Udzungwa mountains of Tanzania. Mammalia 2007, 71, 47–55. [Google Scholar] [CrossRef]
- Cordeiro, N.J.; Lovett, J.C.; Mulungu, E.; Maina, G.G.; Gerstle, J.H. Initial trends of bird assemblages before and after river diversion in an endemic-rich African forest. Biodivers. Conserv. 2006, 15, 971–983. [Google Scholar] [CrossRef]
- Romdal, T.S.; Rahbek, C. Elevational zonation of afrotropical forest bird communities along a homogeneous forest gradient. J. Biogeogr. 2009, 36, 327–336. [Google Scholar] [CrossRef]
- Marshall, A.R.; Topp-Jorgensen, J.E.; Brink, H.; Fanning, E. Monkey abundance and social structure in two high-elevation forest reserves in the Udzungwa Mountains of Tanzania. Int. J. Primatol. 2005, 26, 127–145. [Google Scholar] [CrossRef]
- Marshall, A.R.; Lovett, J.C.; White, P.C.L. Selection of line-transect methods for estimating the density of group-living animals: Lessons from the primates. Am. J. Primatol. 2008, 70, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Rowson, B.; Paustian, M.; Van Goethem, J. New species and records of terrestrial slugs from East Africa (Gastropoda, Urocyclidae, Veronicellidae, Agriolimacidae). Zookeys 2017, 723, 11–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.A.; Rosenmejer, T.; Enghoff, H. A mountain of millipedes IX: Species of the family Gomphodesmidae from the Udzungwa Mountains, Tanzania (Diplopoda, Polydesmida). Eur. J. Taxon. 2020, 675, 1–35. [Google Scholar] [CrossRef]
- Clausnitzer, V.; Dijkstra, K.D.B.; Kipping, J. Globally threatened dragonflies (Odonata) in Eastern Africa and implications for conservation. J. East Afr. Nat. Hist. 2011, 100, 89–111. [Google Scholar] [CrossRef]
- de Jong, R.; Congdon, T.C.E. The montane butterflies of the eastern African forests. In Biogeography and Ecology of the Rain Forests of Eastern Africa; Lovett, J.C., Wasser, S.K., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 133–173. [Google Scholar]
- Zilihona, I.J.E.; Nummelin, M. Coleopteran diversity and abundance in different habitats near Kihansi waterfall, in the Udzungwa Mountains, Tanzania. Biodivers. Conserv. 2001, 10, 769–777. [Google Scholar] [CrossRef]
- Bruhl, C.A.; Mohamed, V.; Linsenmair, K.E. Altitudinal distribution of leaf litter ants along a transect in primary forests on Mount Kinabalu, Sabah, Malaysia. J. Trop. Ecol. 1999, 15, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Folgarait, P.J. Ant biodiversity and its relationship to ecosystem functioning: A review. Biodivers. Conserv. 1998, 7, 1221–1244. [Google Scholar] [CrossRef]
- Andersen, A.N.; Majer, J.D. Ants show the way Down Under: Invertebrates as bioindicators in land management. Front. Ecol. Environ. 2004, 2, 291–298. [Google Scholar] [CrossRef]
- Wiescher, P.T.; Pearce-Duvet, J.M.C.; Feener, D.H. Assembling an ant community: Species functional traits reflect environmental filtering. Oecologia 2012, 169, 1063–1074. [Google Scholar] [CrossRef] [Green Version]
- Kaspari, M.; Weiser, M.D. Ant activity along moisture gradients in a neotropical forest. Biotropica 2000, 32, 703–711. [Google Scholar] [CrossRef]
- Botes, A.; McGeoch, M.A.; Robertson, H.G.; van Niekerk, A.; Davids, H.P.; Chown, S.L. Ants, altitude and change in the northern Cape Floristic Region. J. Biogeogr. 2006, 33, 71–90. [Google Scholar] [CrossRef]
- Burgess, N.D.; Butynski, T.M.; Cordeiro, N.J.; Doggart, N.H.; Fjeldsa, J.; Howell, K.M.; Kilahama, F.B.; Loader, S.P.; Lovett, J.C.; Mbilinyi, B.; et al. The biological importance of the Eastern Arc Mountains of Tanzania and Kenya. Biol. Conserv. 2007, 134, 209–231. [Google Scholar] [CrossRef]
- Shangali, C.F.; Mabula, C.K.; Mmari, C. Biodiversity and human activities in the Udzungwa Mountain forests, Tanzania. 1. Ethnobotanical survey in the Uzungwa scarp forest Reserve. J. East Afr. Nat. Hist. 1998, 87, 291–318. [Google Scholar] [CrossRef] [Green Version]
- Rovero, F.; Owen, N.; Jones, T.; Canteri, E.; Iemma, A.; Tattoni, C. Camera trapping surveys of forest mammal communities in the Eastern Arc Mountains reveal generalized habitat and human disturbance responses. Biodivers. Conserv. 2017, 26, 1103–1119. [Google Scholar] [CrossRef]
- Lovett, J.C. Tanzanian forest tree plot diversity and elevation. J. Trop. Ecol. 1999, 15, 689–694. [Google Scholar] [CrossRef] [Green Version]
- Malumbres-Olarte, J.; Scharff, N.; Pape, T.; Coddington, J.A.; Cardoso, P. Gauging megadiversity with optimized and standardized sampling protocols: A case for tropical forest spiders. Ecol. Evol. 2017, 7, 494–506. [Google Scholar] [CrossRef] [Green Version]
- Fisher, B.L.; Bolton, B. Ants of Africa and Madagascar: A Guide to the Genera; University of California Press: Orland, CA, USA, 2016. [Google Scholar]
- Chao, A.; Ma, K.H.; Hsieh, T.C. iNEXT (iNterpolation and EXTrapolation) Online: Software for Interpolation and Extrapolation of Species Diversity. 2016. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download (accessed on 25 March 2019).
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 9—User’s Guide and Application. Available online: http://purl.oclc.org/estimates (accessed on 25 March 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 25 March 2019).
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Grytnes, J.A.; Vetaas, O.R. Species richness and altitude: A comparison between null models and interpolated plant species richness along the Himalayan altitudinal gradient, Nepal. Am. Nat. 2002, 159, 294–304. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range—An extension of Rapoport latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef]
- Stanley, W.T.; Hutterer, R. Differences in abundance and species richness between shrews and rodents along an elevational gradient in the Udzungwa Mountains, Tanzania. Acta Theriol. 2007, 52, 261–275. [Google Scholar] [CrossRef]
- Dunn, R.R.; Guénard, B.; Weiser, M.D.; Sanders, N.J. Geographic Gradients. Ant Ecol. 2010, 91, 38–58. [Google Scholar]
- Rosenzweig, M.L.; Abramsky, Z. How are diversity and productivity related? Species Divers. Ecol. Communities 1993, 52–65. [Google Scholar]
- Wiens, J.J.; Graham, C.H. Niche conservatism: Integrating evolution, ecology, and conservation biology. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Davies, T.J.; Grytnes, J.A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Ghalambor, C.K.; Huey, R.B.; Martin, P.R. Why mountain passes are higher in the tropics—Revisited. Integr. Comp. Biol. 2004, 44, 558. [Google Scholar]
- Hua, X. The impact of seasonality on niche breadth, distribution range and species richness: A theoretical exploration of Janzen’s hypothesis. Proc. R. Soc. B-Biol. Sci. 2016, 283, 20160349. [Google Scholar] [CrossRef] [Green Version]
- McCain, C.M. Global analysis of bird elevational diversity. Glob. Ecol. Biogeogr. 2009, 18, 346–360. [Google Scholar] [CrossRef]
- Garcia, F.H.; Wiesel, E.; Fischer, G. The ants of Kenya (Hymenoptera: Formicidae): Faunal overview, first species checklist, bibliography, accounts for all genera, and discussion on taxonomy and zoogeography. J. East Afr. Nat. Hist. 2013, 101, 127–222. [Google Scholar] [CrossRef]
- Oyen, K.J.; Giri, S.; Dillon, M.E. Altitudinal variation in bumble bee (Bombus) critical thermal limits. J. Therm. Biol. 2016, 59, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N. Functional groups and patterns of organization in North American ant communities: A comparison with Australia. J. Biogeogr. 1997, 24, 433–460. [Google Scholar] [CrossRef]
- Gathalkar, G.; Sen, A. Foraging and predatory activities of ants. In The Complex World of Ants; IntechOpen: London, UK, 2018; pp. 51–70. [Google Scholar] [CrossRef]
- Ward, P.S. Army ants—The biology of social predation—Gotwald, W.H. Science 1995, 270, 319–320. [Google Scholar] [CrossRef]
- Kenne, M.; Schatz, B.; Durand, J.L.; Dejean, A. Hunting strategy of a generalist ant species proposed as a biological control agent against termites. Entomol. Exp. Et Appl. 2000, 94, 31–40. [Google Scholar] [CrossRef]
- Schoning, C. Driver ants invading a termite nest: Why do the most catholic predators of all seldom take this abundant prey? Biotropica 2007, 39, 663–667. [Google Scholar] [CrossRef]


{kind=link}
| Habitat Types | Altitudinal Range | Description |
|---|---|---|
| Lower elevation forest (lowest elevation) | 650–800 | Forest with deciduous and semi-deciduous trees, canopy 15–25 m with emergents reaching 50 m |
| Sub-montane forest (mid elevation) | 800–1400 | Moist forest with mainly evergreen species, canopy 25–40 m with emergents reaching 50 m |
| Montane forest (high elevation) | 1400–2600 | Evergreen moist forest, with canopy height progressively lower with increasing altitude |
| Subfamily | Species | Individuals (Activities) |
|---|---|---|
| Dolichoderinae | ||
| Technomyrmex | 3 | 5 |
| Dorylinae | ||
| Aenictus | 1 | 2 |
| Dorylus | 1 | 13,260 |
| Parasyscia | 2 | 4 |
| Formicinae | ||
| Camponotus | 5 | 68 |
| Lepisiota | 5 | 209 |
| Nylanderia | 1 | 156 |
| Plagiolepis | 3 | 8 |
| Polyrhachis | 2 | 15 |
| Tapinolepsis | 3 | 10 |
| Myrmicinae | ||
| Calyptomyrmex | 1 | 1 |
| Cardiocondyla | 1 | 6 |
| Carebara | 2 | 126 |
| Catalaucus | 1 | 1 |
| Crematogaster | 5 | 50 |
| Melissotarsus | 1 | 1 |
| Meranoplus | 1 | 7 |
| Microdaceton | 1 | 1 |
| Monomorium | 2 | 49 |
| Myrmicaria | 1 | 10,512 |
| Pheidole | 8 | 4751 |
| Solenopsis | 2 | 308 |
| Strumigenys | 6 | 83 |
| Tetramorium | 19 | 1261 |
| Ponerinae | ||
| Anochectus | 3 | 5 |
| Bothroponera | 4 | 79 |
| Cryptopone | 1 | 1 |
| Hypoponera | 4 | 11 |
| Leptogenys | 4 | 449 |
| Megaponera | 1 | 210 |
| Mesoponera | 3 | 35 |
| Odontomachus | 1 | 10 |
| Plectroctena | 3 | 82 |
| Habitat Type | Observed Species per Habitat (Mean SD) | Plot | Altitude m a.s.l | Observed Species Richness per Plot | Simpson’s Diversity Index | Sample Coverage |
|---|---|---|---|---|---|---|
| Lower elevation | 71 (30.6 ± 8.1) | 1 | 650 | 31 | 0.500 | 0.9963 |
| 2 | 650 | 38 | 0.375 | 0.996 | ||
| 7 | 708 | 40 | 0.862 | 0.993 | ||
| 10 | 674 | 22 | 0.405 | 0.989 | ||
| 13 | 659 | 24 | 0.825 | 0.974 | ||
| Sub-montane | 44 (19.6 ± 5.3) | 3 | 1005 | 24 | 0.297 | 0.998 |
| 4 | 993 | 21 | 0.447 | 0.995 | ||
| 8 | 978 | 25 | 0.623 | 0.998 | ||
| 11 | 1006 | 15 | 0.666 | 0.996 | ||
| 14 | 908 | 13 | 0.813 | 0.911 | ||
| Montane | 33 (15.2 ± 6.5) | 5 | 1448 | 23 | 0.747 | 0.994 |
| 6 | 1482 | 18 | 0.271 | 1 | ||
| 9 | 1527 | 15 | 0.820 | 0.995 | ||
| 12 | 1552 | 15 | 0.466 | 0.986 | ||
| 15 | 1531 | 5 | 0.261 | 0.986 |
| Habitat Type | Species | Indicator Value (%) |
|---|---|---|
| Lower elevation | Pheidole sp.05 | 100 |
| Megaponera analis rapax | 99.52 | |
| Camponotus sp.02. (etiolipes gp.) | 94.12 | |
| Tetramorium cf. yarthiellum | 80 | |
| Nylanderia sp.01 | 70.77 | |
| Sub-montane | Bothroponera sp.01 | 85.71 |
| Myrmicaria rustica angustior | 81.23 | |
| Montane | Tetramorium sp.14 | 100 |
| Mesoponera sp.02 | 82.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunene, C.; Foord, S.H.; Scharff, N.; Pape, T.; Malumbres-Olarte, J.; Munyai, T.C. Ant Diversity Declines with Increasing Elevation along the Udzungwa Mountains, Tanzania. Diversity 2022, 14, 260. https://doi.org/10.3390/d14040260
Kunene C, Foord SH, Scharff N, Pape T, Malumbres-Olarte J, Munyai TC. Ant Diversity Declines with Increasing Elevation along the Udzungwa Mountains, Tanzania. Diversity. 2022; 14(4):260. https://doi.org/10.3390/d14040260
Chicago/Turabian StyleKunene, Caroline, Stefan H. Foord, Nikolaj Scharff, Thomas Pape, Jagoba Malumbres-Olarte, and Thinandavha C. Munyai. 2022. "Ant Diversity Declines with Increasing Elevation along the Udzungwa Mountains, Tanzania" Diversity 14, no. 4: 260. https://doi.org/10.3390/d14040260
APA StyleKunene, C., Foord, S. H., Scharff, N., Pape, T., Malumbres-Olarte, J., & Munyai, T. C. (2022). Ant Diversity Declines with Increasing Elevation along the Udzungwa Mountains, Tanzania. Diversity, 14(4), 260. https://doi.org/10.3390/d14040260