
Diversity Phenomenon of the Danaceinae Malachite Beetle Subfamily (Coleoptera: Dasytidae) in Eocene Baltic Amber with a New Description of an Extinct Genus and Species



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
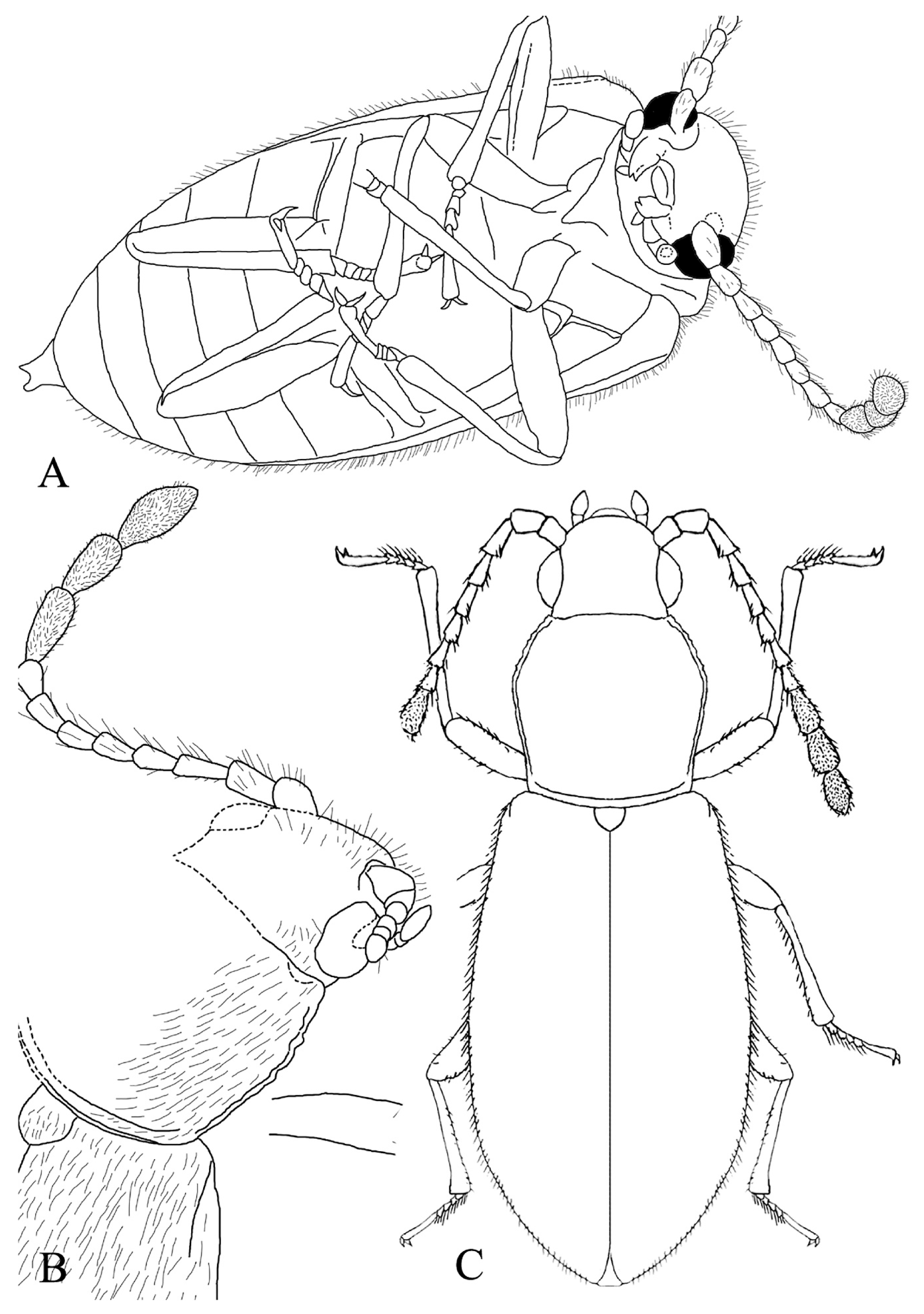
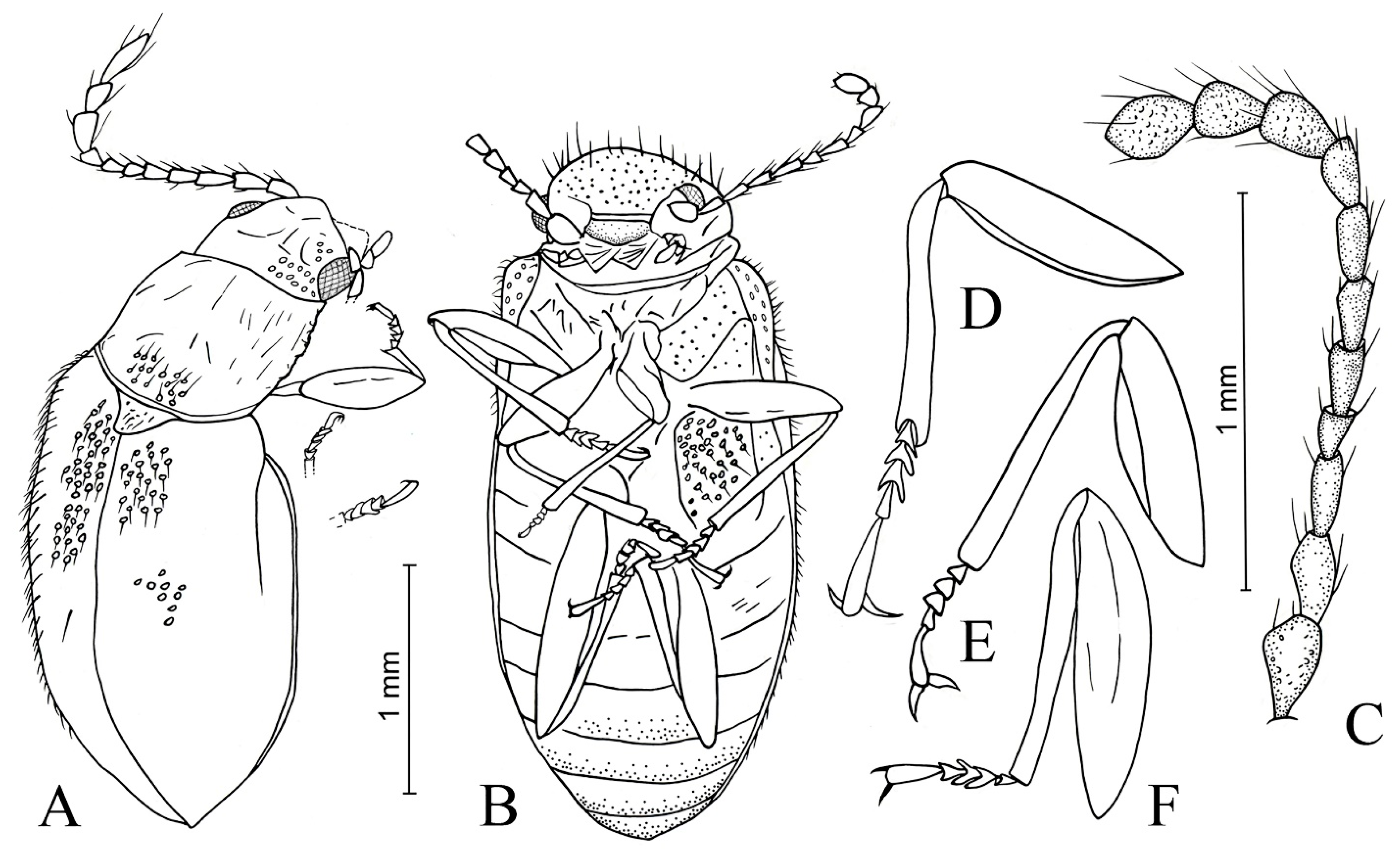
3. Results
- Superfamily Cleroidea Latreille, 1802
- Family Dasytidae Laporte de Castelnau, 1840
- Subfamily Danaceinae C. G. Thompson, 1859
- Tribe Amauroniodini Majer, 1987
- Genus Baltamauroania gen. nov.
- LSID: urn:lsid:zoobank.org:act:72BB8249-538E-48C4-994A-BE4852BF0C9D
- Type species: Baltamauroania mirabilicornis sp. nov., by present designation.
- Baltamauroania mirabilicornis sp. nov.
- LSID: urn:lsid:zoobank.org:act:C658927F-9C1A-4FBE-A855-24EA8EF11362
4. Discussion
- Tribe Amauroniodini Majer, 1987 (Amauronia Westwood, 1839, Baltamauroania gen. nov., Indamauronia Majer, 1997, Mauroania Majer, 1997, Perisphinctus Schilsky 1898, Protomauroania Tshernyshev, 2021 and Pseudamauronia Pic, 1915);
- Tribe Danacaeomimini Majer, 1987 (Cyrdobaldus Majer, 1989, Cyrdothrix Majer, 1989, Danacaeomimus Champion, 1922, Piechockia Majer, 1986, Schoenia Majer, 1989);
- Tribe Danaceini Kiesenwetter, 1863 (Amecocerus Solier, 1849, Danacea Laporte de Castelnau, 1836, Haplamaurus Kirsch, 1865, Hylodanacaea Pic, 1926, Listrocerus Majer, 1998, Setigerodasytes Pic, 1910, Bilyella Majer, 1988 and Hoppingiana Blaisdell, 1924).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Majer, K. A review of the classification of the Melyridae and related families (Coleoptera, Cleroidea). Entomol. Basilensia 1994, 17, 319–390. [Google Scholar]
- Mayor, A.J. Family Malachiidae Fleming, 1821. In Catalogue of Palaearctic Coleoptera, Volume 4. Elateroidea, Derodontoidea, Bostrichoidea, Lymexyloidea, Cleroidea, Cucujoidea; Lobl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; pp. 415–454. [Google Scholar]
- Tihelka, E.; Huang, D.; Perrichot, V.; Cai, C. A previously missing link in the evolution of dasytine soft-winged flower beetles from Cretaceous Charentese amber (Coleoptera, Melyridae). Pap. Palaeontol. 2021, 7, 1753–1764. [Google Scholar] [CrossRef]
- Lawrence, J.F.; Leschen, R.A.B. 9.11. Melyridae Leach, 1815. In Handbook of Zoology—Coleoptera. Volume 2; Leschen, R., Beutel, R., Eds.; De Gruyter: Berlin, Germany; New York, NY, USA, 2010; pp. 273–280. [Google Scholar]
- Mawdsley, J.R. The importance of species of Dasytinae (Coleoptera: Melyridae) as pollinators in Western North America. Coleopt. Bull. 2003, 57, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Tshernyshev, S.E. A review of oedemerid and dasytid beetle fauna (Coleoptera, Oedemeridae, Melyridae, Dasytidae) of North Asia. Euroasian Entomol. J. 2017, 16, 554–565. [Google Scholar]
- Mirutenko, V.V. The beetles of Malachiidae and Dasytidae families Coleoptera) in the collections of Zoological museum and Department of entomology and biodiversity reserve of Uzhhorod university. Sci. Bull. Uzhgorod Univ. Biol. Ser. 2008, 23, 209–219. [Google Scholar]
- Tshernyshev, S.E. Evolution potential of eocene entomofaunas in formation of contemporary diversity of Coleoptera of the families Malachiidae and Dasytidae sensu latum. In Conceptual and Applied Aspects of Scientific Research and Education in the Field of Invertebrate Zoology: Collection of Articles V International Conference; Bagirov, R.T.-O., Maximova, Y.V., Subbotina, E.Y., Shcherbakov, M.V., Simakova, A.V., Eds.; Tomsk State University Press: Tomsk, Russia, 2020; pp. 178–182. [Google Scholar]
- Néraudeau, D.; Allain, R.; Perrichot, V.; Videt, B.; De Broin, F.D.L.; Guillocheau, F.; Philippe, M.; Rage, J.-C.; Vullo, R. Découverte d’un dépôt paralique à bois fossiles, ambre insectifère et restes d’Iguanodontidae (Dinosauria, Ornithopoda) dans le Cénomanien inférieur de Fouras (Charente-Maritime, Sud-Ouest de la France). Comptes Rendus Palevol 2003, 2, 221–230. [Google Scholar] [CrossRef]
- Perrichot, V.; Néraudeau, D.; Tafforeau, P. Charentese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 192–208. [Google Scholar]
- Kolibač, J.; Perkovsky, E.E. A reclassification of Acanthocnemoides sukatshevae Zherikhin, 1977 from the mid-Cretaceous Taimyr amber (Coleoptera). Cretac. Res. 2020, 115, 104548. [Google Scholar] [CrossRef]
- Zherikhin, V.V. Series of Superfamilies of Cucujiformia. In Mezozoiskie zhestkokrylye; Arnoldi, L.V., Zherikhin, V.V., Nikritin, L.M., Ponomarenko, A.G., Eds.; Trudy Paleontologicheskogo Instituta: Moscow, Russia, 1977; Volume 161, pp. 135–142. (In Russian) [Google Scholar]
- Zherikhin, V.V. Series of superfamilies of Cucujiformia. In Mesozoic Coleoptera; Arnoldi, L.V., Zherikhin, V.V., Nikritin, L.M., Ponomarenko, A.G., Eds.; Oxonian Press Pvt. Ltd.: New Delhi, India, 1992; pp. 182–191, (Complete English translation of the previous reference Zherikhin, 1977). [Google Scholar]
- Schmitz, H.; Schmitz, A.; Trenner, S.; Bleckmann, H. A new type of insect infrared organ of low thermal mass. Naturwissenschaften 2002, 89, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Z.; Yu, Y.; Shih, C.; Ślipiński, A.; Ren, D. First species of Prionoceridae from mid-Cretaceous amber of northern Myanmar (Coleoptera: Cleroidea). Cretac. Res. 2021, 133, 105118. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Legalov, A.A. First record of the family Malachiidae (Coleoptera: Cleroidea) from mid-Cretaceous Burmese amber with a description of Burmalachius acroantennatus gen. et spec. nov. Life 2023, 13, 1938. [Google Scholar] [CrossRef]
- Bocakova, M.; Constantin, R.; Bocak, L. Molecular phylogenetics of the melyrid lineage (Coleoptera: Cleroidea). Cladistics 2012, 28, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Majer, K. Rhadalinae from the Baltic amber (Coleoptera, Dasytidae). Dtsch. Entomol. Z. 1998, 45, 255–264. [Google Scholar] [CrossRef]
- Tshernyshev, S.E. Aploceble (Chalcoaploceble) viridiaeneus Tshernyshev—New subgenus and species of dasytid beetles (Coleopterta, Dasytidae) in Baltic amber. Euroasian Entomol. J. 2012, 11, 213–218. [Google Scholar]
- Tshernyshev, S.E. New species of malachite beetles (Coleoptera, Dasytidae) in Baltic amber. Paleontol. J. 2019, 53, 1024–1035. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Shcherbakov, M.V. A new genus and species of malachite beetles (Coleoptera, Dasytidae) in Baltic amber. Paleontol. J. 2021, 55, 174–178. [Google Scholar] [CrossRef]
- Tshernyshev, S.E.; Perkovsky, E.E. Protomauroania mikhailovi—A new species of malachite beetles (Coleoptera, Dasytidae) in Rovno Amber. Zootaxa 2021, 5006, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.F. A report of some recent collections of fossil Coleoptera from the Miocene shales of Florissant. Iowa City Bull. Lab. Nat. Hist. Univ. Iowa 1912, 6, 3–38. [Google Scholar]
- Wickham, H.F. Twenty new Coleoptera from the Florissant Shales. Trans. Am. Entomol. Soc. 1914, 40, 257–270. [Google Scholar]
- Wickham, H.F. New fossil Coleoptera from the Florissant beds. Bull. Lab. Natl. Hist. State Univ. Iowa 1916, 7, 3–20. [Google Scholar]
- Wickham, H.F. New species of fossil beetles from Florissant, Colorado. Proc. U. S. Natl. Mus. 1917, 52, 463–472. [Google Scholar] [CrossRef]
- Majer, K. Subfamilial classification of the family Malachiidae (Coleoptera, Cleroidea). Entomol. Basiliensia 2002, 24, 179–244. [Google Scholar]
- Constantin, R. Malachiidae Fleming, 1821. In Catalogue des Coleopteres de France; Tronquet, M., Ed.; Association Roussillonnaise d’Entomologie: Perpignan, France, 2021; Volume 487, 492p. [Google Scholar]
- Gimmel, M.L.; Bocakova, M.; Gunter, N.L.; Leschen, R.A.B. Comprehensive phylogeny of the Cleroidea (Coleoptera: Cucujiformia). Syst. Entomol. 2019, 44, 527–558. [Google Scholar] [CrossRef]
- Hoffeins, H.W. On the preparation and conservation of amber inclusions in artificial resin. Pol. Pismo Entomol. 2001, 70, 215–219. [Google Scholar]
- Majer, K. Comparative morphology and proposed major taxonomy of the Family Melyridae (Insecta, Coleoptera). Pol. Pismo Entomol. 1987, 56, 719–859. [Google Scholar]
- Sadowski, E.-M.; Seyfullah, L.J.; Schmidt, A.R.; Kunzmann, L. Conifers of the ‘Baltic amber forest’ and their palaeoeco-logical significance. Stapfia 2017, 106, 1–73. [Google Scholar]
- Sadowski, E.-M.; Schmidt, A.R.; Denk, T. Staminate inflorescences with in situ pollen from Eocene Baltic amber reveal high diversity in Fagaceae (oak family). Willdenowia 2020, 50, 405–517. [Google Scholar] [CrossRef]
- Seyfullah, L.J.; Beimforde, C.; Dal Corso, J.; Perrichot, V.; Rikkinen, J.; Schmidt, A.R. Production and preservation of resins—Past and present. Biol. Rev. 2018, 93, 1684–1714. [Google Scholar] [CrossRef] [PubMed]
- Bukejs, A.; Alekseev, V.I.; Pollock, D.A. Waidelotinae, a new subfamily of Pyrochroidae (Coleoptera: Tenebrionoidea) from Baltic amber of the Sambian peninsula and the interpretation of Sambian amber stratigraphy, age and location. Zootaxa 2019, 4664, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Kasiński, J.R.; Kramarska, R.; Słodkowska, B.; Sivkov, V.; Piwocki, M. Paleocene and Eocene deposits on the eastern margin of the Gulf of Gdańsk (Yantarny P-1 bore hole, Kaliningrad region, Russia). Geol. Q. 2020, 64, 29–53. [Google Scholar] [CrossRef]
- Bukejs, A.; Alekseev, V.I.; Legalov, A.A. A new Eocene genus of the subtribe Tylodina (Coleoptera: Curculionidae) and notes concerning local differences of Baltic amber in the Kaliningrad Region. Foss. Rec. 2020, 23, 75–81. [Google Scholar] [CrossRef]
- Arillo, A. Paleoethology: Fossilized behaviours in amber. Geol. Acta 2007, 5, 159–166. [Google Scholar]
- Berendt, G.C. Die organischen Bernstein-Einschlüsse im Allgemeinen. In Der Bernstein und Die in Ihm Befindlichen Pflanzenreste der Vorwelt. Bd.1; Goeppert, H.R., Berendt, G.C., Eds.; Nicolai: Berlin, Germany, 1845; pp. 41–60. [Google Scholar]
- Klebs, R. Über Bernsteineinschlüsse im allgemeinen und die Coleopteren meiner Bersteinsammlung. Schr. Phys.-Okon. Ges. Königsberg 1910, 51, 217–242. [Google Scholar]
- Spahr, U. Bibliographie der Bernstein- und Kopal-Käfer (Coleoptera). Stutt. Beitr. Natukd. Ser. B 1981, 72, 1–22. [Google Scholar]
- Spahr, U. Systematischer Katalog der Bernstein- und Kopal-Käfer (Coleoptera). Stutt. Beitr. Natukd. Ser. B 1981, 80, 1–107. [Google Scholar]
- Tshernyshev, S.E. New Taxa of Soft-winged Flower Beetles (Coleoptera, Malachiidae) in Baltic and Rovno Amber. Paleontol. J. 2016, 50, 953–962. [Google Scholar] [CrossRef]
- Tshernyshev, S.E. New taxa of soft-winged flower beetles (Coleoptera, Malachiidae) in Baltic Amber. Paleontol. J. 2020, 54, 67–75. [Google Scholar] [CrossRef]
- Kubisz, D. Beetles in the collection of the Museum of Amber Inclusions, University of Gdansk, with description of Colotes sambicus sp. n. (Coleoptera: Melyridae). Pol. J. Entomol. 2001, 70, 259–265. [Google Scholar]
- Kirejtshuk, A.G.; Nel, A. New beetles of the suborder Polyphaga from the lowermost Eocene French amber (Insecta: Coleoptera). Ann. Soc. Entomol. Fr. 2008, 44, 419–442. [Google Scholar] [CrossRef]
- Mawdsley, J.R. Review of the extant and fossil Dasytinae (Coleoptera: Melyridae) of Colorado, U.S.A. Trans. Am. Entomol. Soc. Phila. 1999, 125, 251–267. [Google Scholar]
- Rasnitsyn, A.P.; Ross, A.J. A preliminary list of arthropod families present in the Burmese amber collection at The Natural History Museum, London. Bull.-Nat. Hist. Mus. Geol. Ser. 2000, 56, 21–24. [Google Scholar]
- Ross, A.; Mellish, C.; York, P.; Craighton, B. Chapter 12: Burmese amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 116–136. [Google Scholar]
- Ross, A.J. Burmese (Myanmar) amber checklist and bibliography 2018. Palaeoentomology 2019, 2, 22–84. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Ponomarenko, A.G.; Prokin, A.A.; Chang, H.; Nikolajev, G.V.; Ren, D. Current knowledge on Mesozoic Coleoptera from Daohugou and Liaoning (North East China). Acta Geol. Sin. 2010, 84, 783–792. [Google Scholar] [CrossRef]
- Handlirsch, A. IV. Abschnitt. Tertiäre Insekten. In Die Fossilen Insekten und die Phylogenie der Rezenten Formen: Ein Handbuch für Paläontologen und Zoologen; Handlirsch, A., Ed.; Wilhelm Engelman: Leipzig, Germany, 1907; pp. 673–1120. [Google Scholar]
- Bachofen-Echt, A. Der Bernstein und Seine Einschlüsse; Springer: Berlin/Heidelberg, Germany, 1949. [Google Scholar]
- Larsson, S.G. Baltic Amber—A Palaeobiological Study. Entomonograph 1; Scandinavian Science Press Ltd.: Klampenborg, Denmark, 1978. [Google Scholar]
- Perkovsky, E.E.; Vasilenko, D.V. A summary of recent results in the study of Taimyr amber. Paleontol. J. 2019, 53, 984–993. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tshernyshev, S.E.; Lyubarsky, G.Y.; Alekseev, V.; Bukejs, A. Diversity Phenomenon of the Danaceinae Malachite Beetle Subfamily (Coleoptera: Dasytidae) in Eocene Baltic Amber with a New Description of an Extinct Genus and Species. Diversity 2023, 15, 1077. https://doi.org/10.3390/d15101077
Tshernyshev SE, Lyubarsky GY, Alekseev V, Bukejs A. Diversity Phenomenon of the Danaceinae Malachite Beetle Subfamily (Coleoptera: Dasytidae) in Eocene Baltic Amber with a New Description of an Extinct Genus and Species. Diversity. 2023; 15(10):1077. https://doi.org/10.3390/d15101077
Chicago/Turabian StyleTshernyshev, Sergei E., Georgy Yu. Lyubarsky, Vitalii Alekseev, and Andris Bukejs. 2023. "Diversity Phenomenon of the Danaceinae Malachite Beetle Subfamily (Coleoptera: Dasytidae) in Eocene Baltic Amber with a New Description of an Extinct Genus and Species" Diversity 15, no. 10: 1077. https://doi.org/10.3390/d15101077
APA StyleTshernyshev, S. E., Lyubarsky, G. Y., Alekseev, V., & Bukejs, A. (2023). Diversity Phenomenon of the Danaceinae Malachite Beetle Subfamily (Coleoptera: Dasytidae) in Eocene Baltic Amber with a New Description of an Extinct Genus and Species. Diversity, 15(10), 1077. https://doi.org/10.3390/d15101077