
Biological Integrity of Azorean Native Forests Is Better Measured in Cold Season

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Selecting Data Sets
2.3. IBI Calculation and Data Analysis
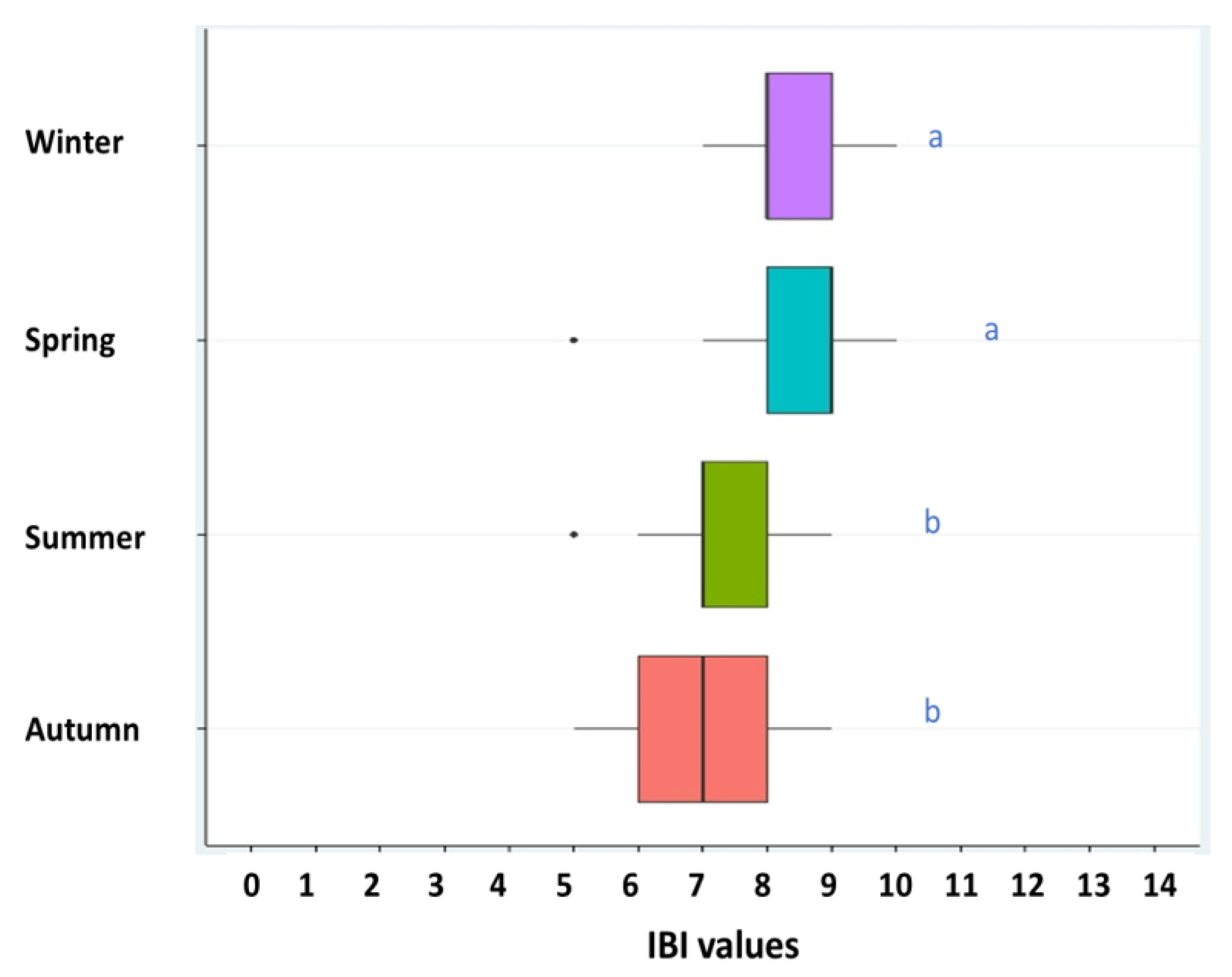
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borges, P.A.V.; Cardoso, P.; Kreft, H.; Whittaker, R.J.; Fattorini, S.; Emerson, B.C.; Gil, A.; Gillespie, R.G.; Matthews, T.J.; Santos, A.M.C.; et al. Global Island Monitoring Scheme (GIMS): A proposal for the long-term coordinated survey and monitoring of native island forest biota. Biodivers. Conserv. 2018, 27, 2567–2586. [Google Scholar] [CrossRef]
- Emerson, B.C.; Borges, P.A.V.; Cardoso, P.; Convey, P.; Dewaard, J.R.; Economo, E.P.; Gillespie, R.G.; Kennedy, S.; Krehenwinkel, H.; Meier, R.; et al. Collective and harmonized high throughput barcoding of insular arthropod biodiversity: Toward a Genomic Observatories Network for islands. Mol. Ecol. 2022, 32, 6161–6176. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/mec.16683 (accessed on 30 May 2023). [CrossRef] [PubMed]
- Fernández-Palacios, J.M. Why islands’? In Islands and Evolution; Pérez-Mellado, V., Ramon, C., Eds.; Institut Menorquí d’Estudis: Mahón, Spain, 2010; Volume 19, pp. 85–109. [Google Scholar]
- Elias, R.B. Ecologia das florestas de Juniperus dos Açores. Ph.D. Dissertation, Universidade dos Açores, Angra do Heroísmo, Portugal, 2007. [Google Scholar]
- Elias, R.B.; Gil, A.; Fernández-Palacios, J.M.; Azevedo, E.B.; Reis, F. Natural zonal vegetation of the Azores Islands: Characterization and potential distribution. Phytocoenologia 2016, 46, 107–123. [Google Scholar] [CrossRef]
- Nunes, R.; Gabriel, R.; Elias, R.B.; Rigal, F.; Borges, P.A. Arthropods and other Biota associated with the Azorean Trees and Shrubs: Juniperus brevifolia. Arquipélago-Life Mar. Sci. 2015, 32, 19–48. [Google Scholar]
- Tsafack, N.; Gabriel, R.; Elias, R.; Boieiro, M.; Ferreira, M.T.; Borges, P. Arthropods and other biota associated with the Azorean trees and shrubs: Laurus azorica (Seub) Franco (Magnoliophyta, Magnoliopsida, Laurales, Lauraceae). Biodivers. Data J. 2022, 10, e80088. [Google Scholar] [CrossRef]
- Borges, P. A list of the terrestrial and marine biota from the Azores. In Princípia; Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente, M., Raposeiro, P., Rodrigues, P., et al., Eds.; Cascais, Portugal, 2010; Available online: https://agris.fao.org/search/en/providers/123417/records/6474616b79cbb2c2c1acaf6e (accessed on 27 November 2023).
- Coelho, M.C.M.; Gabriel, R.; Hespanhol, H.; Borges, P.A.V.; Ah-Peng, C. Bryophyte Diversity along an Elevational Gradient on Pico Island (Azores, Portugal). Diversity 2021, 13, 162. [Google Scholar] [CrossRef]
- Gabriel, R.; Bates, J.W. Bryophyte community composition and habitat specificity in the natural forests of Terceira, Azores. Plant Ecol. 2005, 177, 125–144. [Google Scholar] [CrossRef]
- Ferreira, M.T.; Cardoso, P.; Borges, P.A.; Gabriel, R.; de Azevedo, E.B.; Reis, F.; Araújo, M.B.; Elias, R.B. Effects of climate change on the distribution of indigenous species in oceanic islands (Azores). Clim. Chang. 2016, 138, 603–615. [Google Scholar] [CrossRef]
- Santos, F.D.; Valente, M.A.; Miranda PM, A.; Aguiar, A.; Azevedo, E.B.; Tomé, A.R.; Coelho, F. Climate change scenarios in the Azores and Madeira islands. World Resour. Rev. 2004, 16, 473–491. [Google Scholar]
- Wallon, S.; Melo, C.; Tsafack, N.; Elias, R.; Borges, P. Assessing the effects of climate change on arthropod abundance in Azorean pastures: PASTURCLIM project’s baseline monitoring data. Biodivers. Data J. 2023, 11, e103723. [Google Scholar] [CrossRef]
- Borges, P.A.V.; Rigal, F.; Ros-Prieto, A.; Cardoso, P. Increase of insular exotic arthropod diversity is a fundamental dimension of the current biodiversity crisis. Insect Conserv. Divers. 2020, 13, 508–518. [Google Scholar] [CrossRef]
- Pozsgai, G.; Cardoso, P.; Rigal, F.; Boieiro, M.; Gabriel, R.; de Azevedo, E.B.; Borges, P.A.V. Arthropod co-occurrence networks indicate environmental differences between islands and signal introduced species in Azorean native forest remnants. Front. Ecol. Evol. 2023, 11, 262. [Google Scholar] [CrossRef]
- Costa, R.; Cardoso, P.; Rigal, F.; Borges, P.A.V. Island spider origins show complex vertical stratification patterns in Macaronesia. Insect Conserv. Divers. 2023, 16, 886–895. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/icad.12686 (accessed on 30 September 2023). [CrossRef]
- Brush, M.; Matthews, T.J.; Borges, P.A.V.; Harte, J. Land use change through the lens of macroecology: Insights from Azorean arthropods and the maximum entropy theory of ecology. Ecography 2022, 2022, e06141. [Google Scholar] [CrossRef]
- Gaston, K.J.; Borges, P.A.V.; He, F.; Gaspar, C. Abundance, spatial variance and occupancy: Arthropod species distribution in the Azores. J. Anim. Ecol. 2006, 75, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, H.; Moura, M.; Maciel, M.G.B.; Silva, L.; Rumsey, F.J.; Carine, M.A. The Linnean shortfall in oceanic island biogeography: A case study in the Azores. J. Biogeogr. 2011, 38, 1345–1355. [Google Scholar] [CrossRef]
- Tsafack, N.; Lhoumeau, S.; Ros-Prieto, A.; Navarro, L.; Kocsis, T.; Manso, S.; Figueiredo, T.; Ferreira, M.T.; Borges, P.A.V. Arthropod-based biotic integrity indices: A novel tool for evaluating the ecological condition of native forests in the Azores archipelago. Ecol. Indic. 2023, 154, 110592. [Google Scholar] [CrossRef]
- Cardoso, P.; Borges, P.A.V.; Gaspar, C. Biotic integrity of the arthropod communities in the natural forests of Azores. Biodivers. Conserv. 2007, 16, 2883–2901. [Google Scholar] [CrossRef]
- Karr, J.R.; Fausch, K.D.; Angermeier, P.L.; Yant, P.R.; Schlosser, I.J. Assessing Biological Integrity in Running Waters, a Method and Its Rationale. Illinois Natural History Survey, Champaign. 1986. Available online: https://semspub.epa.gov/work/01/554353.pdf (accessed on 27 March 2023).
- Suggitt, A.J.; Gillingham, P.K.; Hill, J.K.; Huntley, B.; Kunin, W.E.; Roy, D.B.; Thomas, C.D. Habitat microclimates drive fine-scale variation in extreme temperatures. Oikos 2011, 120, 1–8. [Google Scholar] [CrossRef]
- Borges, P.; Lamelas-Lopez, L.; Andrade, R.; Lhoumeau, S.; Vieira, V.; Soares, A.; Borges, I.; Boieiro, M.; Cardoso, P.; Crespo, L.C.; et al. An updated checklist of Azorean arthropods (Arthropoda). Biodivers. Data J. 2022, 10, e97682. [Google Scholar] [CrossRef]
- Borges, P.A.; Gabriel, R.; Arroz, A.M.; Costa, A.; Cunha, R.T.; Silva, L.; Mendonça, E.; Martins, A.M.F.; Reis, F.; Cardoso, P. The Azorean Biodiversity Portal: An internet database for regional biodiversity outreach. System. Biodivers. 2010, 8, 423–434. [Google Scholar] [CrossRef]
- Costa, R.; Borges, P. SLAM Project—Long Term Ecological Study of the Impacts of Climate Change in the natural forest of Azores: I—The spiders from native forests of Terceira and Pico Islands (2012–2019). BDJ 2021, 9, e69924. [Google Scholar] [CrossRef] [PubMed]
- Lhoumeau, S.; Cardoso, P.; Boieiro, M.; Ros-Prieto, A.; Costa, R.; Lamelas-Lopez, L.; Leite, A.; Rosário, I.A.D.; Gabriel, R.; Malumbres-Olarte, J.; et al. SLAM Project—Long Term Ecological Study of the Impacts of Climate Change in the natural forests of Azores: V—New records of terrestrial arthropods after ten years of SLAM sampling. Biodivers. Data J. 2022, 10, e97952. [Google Scholar] [CrossRef]
- Lhoumeau, S.; Cardoso, P.; Costa, R.; Boieiro, M.; Malumbres-Olarte, J.; Amorim, I.; Rigal, F.; Santos, A.; Gabriel, R.; Borges, P. SLAM Project—Long Term Ecological Study of the Impacts of Climate Change in the natural forest of Azores: IV—The spiders of Terceira and Pico Islands (2019–2021) and general diversity patterns after ten years of sampling. BDJ 2022, 10, e96442. [Google Scholar] [CrossRef]
- Tsafack, N.; Fattorini, S.; Boieiro, M.; Rigal, F.; Ros-Prieto, A.; Ferreira, M.T.; Borges, P.A.V. The Role of Small Lowland Patches of Exotic Forests as Refuges of Rare Endemic Azorean Arthropods. Diversity 2021, 13, 443. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2023. Available online: https://www.R-project.org (accessed on 1 May 2023).
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests’. 2023. Available online: https://cran.r-project.org/web/packages/rstatix/index.html (accessed on 1 May 2023).
- Lhoumeau, S.; Borges, P.A.V. Assessing the Impact of Insect Decline in Islands: Exploring the Diversity and Community Patterns of Indigenous and Non-Indigenous Arthropods in the Azores Native Forest over 10 Years. Diversity 2023, 15, 753. [Google Scholar] [CrossRef]
- Alexandrino, E.R.; Buechley, E.R.; Karr, J.R.; Ferraz, K.M.P.M.d.B.; Ferraz, S.F.d.B.; Couto, H.T.Z.D.; Şekercioğlu, H. Bird based Index of Biotic Integrity: Assessing the ecological condition of Atlantic Forest patches in human-modified landscape. Ecol. Indic. 2017, 73, 662–675. [Google Scholar] [CrossRef]
- Kane, D.D.; Gordon, S.I.; Munawar, M.; Charlton, M.N.; Culver, D.A. The Planktonic Index of Biotic Integrity (P-IBI): An approach for assessing lake ecosystem health. Ecol. Indic. 2009, 9, 1234–1247. [Google Scholar] [CrossRef]
- Zhu, H.; Hu, X.-D.; Wu, P.-P.; Chen, W.-M.; Wu, S.-S.; Li, Z.-Q.; Zhu, L.; Xi, Y.-L.; Huang, R. Development and testing of the phytoplankton biological integrity index (P-IBI) in dry and wet seasons for Lake Gehu. Ecol. Indic. 2021, 129, 107882. [Google Scholar] [CrossRef]
- Jiménez-García, E.; Andújar, C.; López, H.; Emerson, B.C. Towards understanding insect species introduction and establishment: A community-level barcoding approach using island beetles. Mol. Ecol. 2023, 32, 3778–3792. [Google Scholar] [CrossRef]
- Cardoso, P.; Arnedo, M.A.; Triantis, K.A.; Borges, P.A.V. Drivers of diversity in Macaronesian spiders and the role of species extinctions. J. Biogeogr. 2010, 37, 1034–1046. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Sites Code | Island | Site Quality | Longitude | Latitude |
|---|---|---|---|---|
| FLO-NFFR-T-06 | Flores | Disturbed | −31.2235 | 39.4074 |
| FLO-NFFR-T-07 | Flores | Disturbed | −31.2175 | 39.4032 |
| FLO-NFMA-T-08 | Flores | Preserved | −31.2094 | 39.4600 |
| PIC-ML-400 | Pico | Disturbed | −28.4311 | 38.5207 |
| PIC-NFCA-T-08 | Pico | Preserved | −28.2000 | 38.4408 |
| PIC-NFMP-T-10 | Pico | Disturbed | −28.2759 | 38.4630 |
| TER-0M | Terceira | Disturbed | −27.3748 | 38.7666 |
| TER-NFBF-T-01 | Terceira | Preserved | −27.2193 | 38.7618 |
| TER-NFPG-T-33 | Terceira | Disturbed | −27.2271 | 38.7334 |
| TER-NFSB-T164 | Terceira | Preserved | −27.3074 | 38.7355 |
| TER-NFTB-T-15 | Terceira | Preserved | −27.2006 | 38.7364 |
| TER-NFTB-T-18-O | Terceira | Preserved | −27.1980 | 38.7323 |
| Parameters | Score 0 | Score 1 | Score 2 |
|---|---|---|---|
| sEnd | <18 | 18–30 | >30 |
| nEnd | <35 | 35–63 | >63 |
| sNat | <38 | 38–46 | >46 |
| nNat | >47 | 33–47 | <33 |
| sSap | >25 | 20–25 | <20 |
| nSap | >33 | 14–33 | <14 |
| nInt | >19 | 5–19 | <5 |
| END | NAT | INT | χ2 | p | ||
|---|---|---|---|---|---|---|
| Number of individuals | ||||||
| Cold semester | Winter | 3233 | 4131 | 463 | 2802.3 | <0.001 |
| Spring | 10,863 | 12,179 | 1131 | 9039.1 | <0.001 | |
| Warm semester | Summer | 13,737 | 25,846 | 2608 | 19,210 | <0.001 |
| Autumn | 5155 | 8250 | 1885 | 3975.5 | <0.001 | |
| Total | 32,988 | 50,406 | 6087 | 33,429 | <0.001 | |
| Number of morpho (species) | ||||||
| Cold semester | Winter | 32 | 43 | 58 | 7.6842 | 0.021 |
| Spring | 50 | 59 | 76 | 5.6541 | 0.059 | |
| Warm semester | Summer | 48 | 72 | 85 | 10.312 | <0.01 |
| Autumn | 41 | 62 | 82 | 13.632 | <0.01 | |
| Total | 171 | 236 | 301 | 35.805 | <0.001 | |
| Islands | Site Quality | Cold (Winter/Spring) | Warm (Summer/Autumn) | U | p | ||
|---|---|---|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | ||||
| Flores | Disturbed | 4.3 ± 1.03 | [3–6] | 3.6 ±1.06 | [1–4] | 5.04 | 0.283 |
| Flores | Disturbed | 4.0 ± 1.41 | [3–5] | 1.7 ± 0.58 | [1–2] | 4.00 | 0.261 |
| Pico | Disturbed | 1.13 ± 1.46 | [0–4] | 0.9 ± 1.36 | [0–4] | 0.68 | 0.878 |
| Pico | Disturbed | 8.5 ± 0.71 | [8–9] | 6.7 ± 0.58 | [6–7] | 4.00 | 0.261 |
| Terceira | Disturbed | 2.2 ± 1.09 | [1–5] | 1.8 ± 1.23 | [1–4] | 8.83 | 0.032 |
| Terceira | Disturbed | 5.7 ± 1.64 | [4–8] | 4.1 ± 1.81 | [1–7] | 9.82 | 0.133 |
| Flores | Preserved | 8.5 ± 0.71 | [8–9] | 7.0 ± 1 | [6–8] | 2.33 | 0.506 |
| Pico | Preserved | 6.5 ± 2.12 | [5–8] | 8.7 ± 0.58 | [8–9] | 2.33 | 0.311 |
| Terceira | Preserved | 8.5 ± 0.53 | [8–9] | 6.3 ± 1.29 | [4–8] | 14.55 | 0.013 |
| Terceira | Preserved | 9.6 ± 0.53 | [9–10] | 7.9 ± 0.74 | [7–9] | 12.93 | p < 0.01 |
| Terceira | Preserved | 8.5 ± 0.53 | [8–9] | 7.2 ± 0.75 | [6–8] | 11.52 | p < 0.01 |
| Terceira | Preserved | 8.4 ± 0.79 | [8–10] | 7.0 ± 1.12 | [5–8] | 6.53 | 0.258 |
| All Disturbed sites | 3.7 ± 2.5 | [0–9] | 2.8 ± 2.1 | [0–7] | 648 | 0.112 | |
| All Preserved sites | 8.6 ± 0.9 | [5–10] | 7.2 ± 1.1 | [4–9] | 289 | p < 0.001 | |
| Total | 6.3 ± 3.0 | [0–10] | 5.1 ± 2.7 | [0–9] | 2471.5 | p < 0.001 | |
| Site Quality | Season 1 (n) | Season 2 (n) | H | p |
|---|---|---|---|---|
| Disturbed | Winter (29) | Spring (35) | 385.5 | 1 |
| Disturbed | Winter (29) | Summer (41) | 560 | 1 |
| Disturbed | Winter (29) | Autumn (35) | 521.5 | 1 |
| Disturbed | Spring (35) | Summer (41) | 899 | 0.9 |
| Disturbed | Spring (35) | Autumn (35) | 779.5 | 0.7 |
| Disturbed | Summer (41) | Autumn (35) | 778.5 | 1 |
| Preserved | Winter (31) | Spring (34) | 465.5 | 1 |
| Preserved | Winter (31) | Summer (44) | 1030 | 0.001 |
| Preserved | Winter (31) | Autumn (42) | 1062.5 | p < 0.001 |
| Preserved | Spring (34) | Summer (44) | 1165 | p < 0.001 |
| Preserved | Spring (34) | Autumn (42) | 1185.5 | p < 0.001 |
| Preserved | Summer (44) | Autumn (42) | 1079.5 | 1 |
| Disturbed | Winter/Spring (37) | Summer/Autumn (44) | 980 | 1 |
| Preserved | Winter/Spring (40) | Summer/Autumn (47) | 1591 | p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsafack, N.; Lhoumeau, S.; Ros-Prieto, A.; Navarro, L.; Kocsis, T.; Manso, S.; Figueiredo, T.; Ferreira, M.T.; Borges, P.A.V. Biological Integrity of Azorean Native Forests Is Better Measured in Cold Season. Diversity 2023, 15, 1189. https://doi.org/10.3390/d15121189
Tsafack N, Lhoumeau S, Ros-Prieto A, Navarro L, Kocsis T, Manso S, Figueiredo T, Ferreira MT, Borges PAV. Biological Integrity of Azorean Native Forests Is Better Measured in Cold Season. Diversity. 2023; 15(12):1189. https://doi.org/10.3390/d15121189
Chicago/Turabian StyleTsafack, Noelline, Sébastien Lhoumeau, Alejandra Ros-Prieto, Loic Navarro, Timea Kocsis, Sónia Manso, Telma Figueiredo, Maria Teresa Ferreira, and Paulo A. V. Borges. 2023. "Biological Integrity of Azorean Native Forests Is Better Measured in Cold Season" Diversity 15, no. 12: 1189. https://doi.org/10.3390/d15121189
APA StyleTsafack, N., Lhoumeau, S., Ros-Prieto, A., Navarro, L., Kocsis, T., Manso, S., Figueiredo, T., Ferreira, M. T., & Borges, P. A. V. (2023). Biological Integrity of Azorean Native Forests Is Better Measured in Cold Season. Diversity, 15(12), 1189. https://doi.org/10.3390/d15121189