
Population Structure, Distribution, and Spatial Characteristics of Alsophila spinulosa in Chishui, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Square Setting and Community Survey
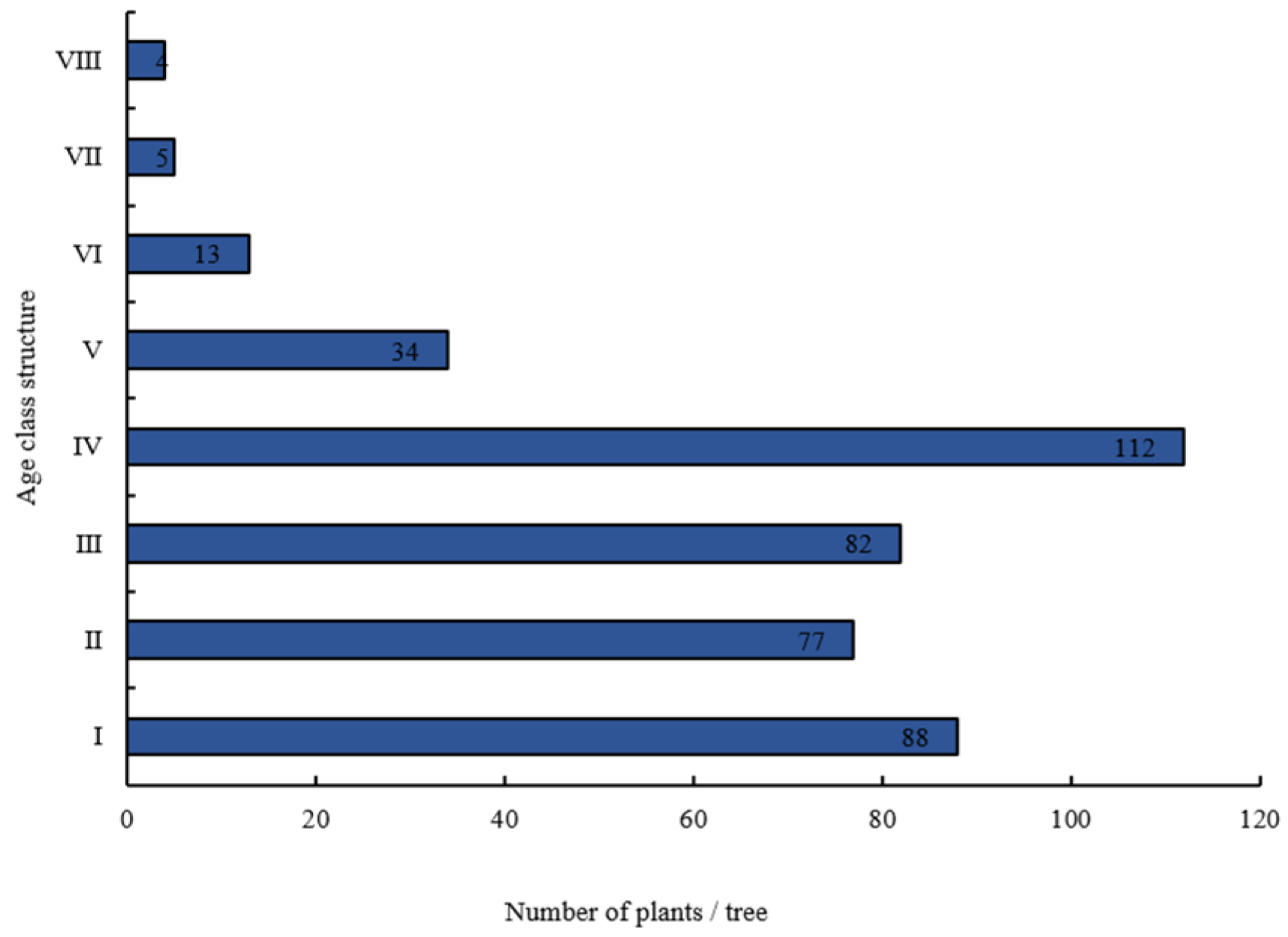
2.3. Age Class Structure Classification
2.4. Point Pattern Analysis
2.5. Data Processing and Analysis
3. Results
3.1. Age Structure of A. spinulosa Populations
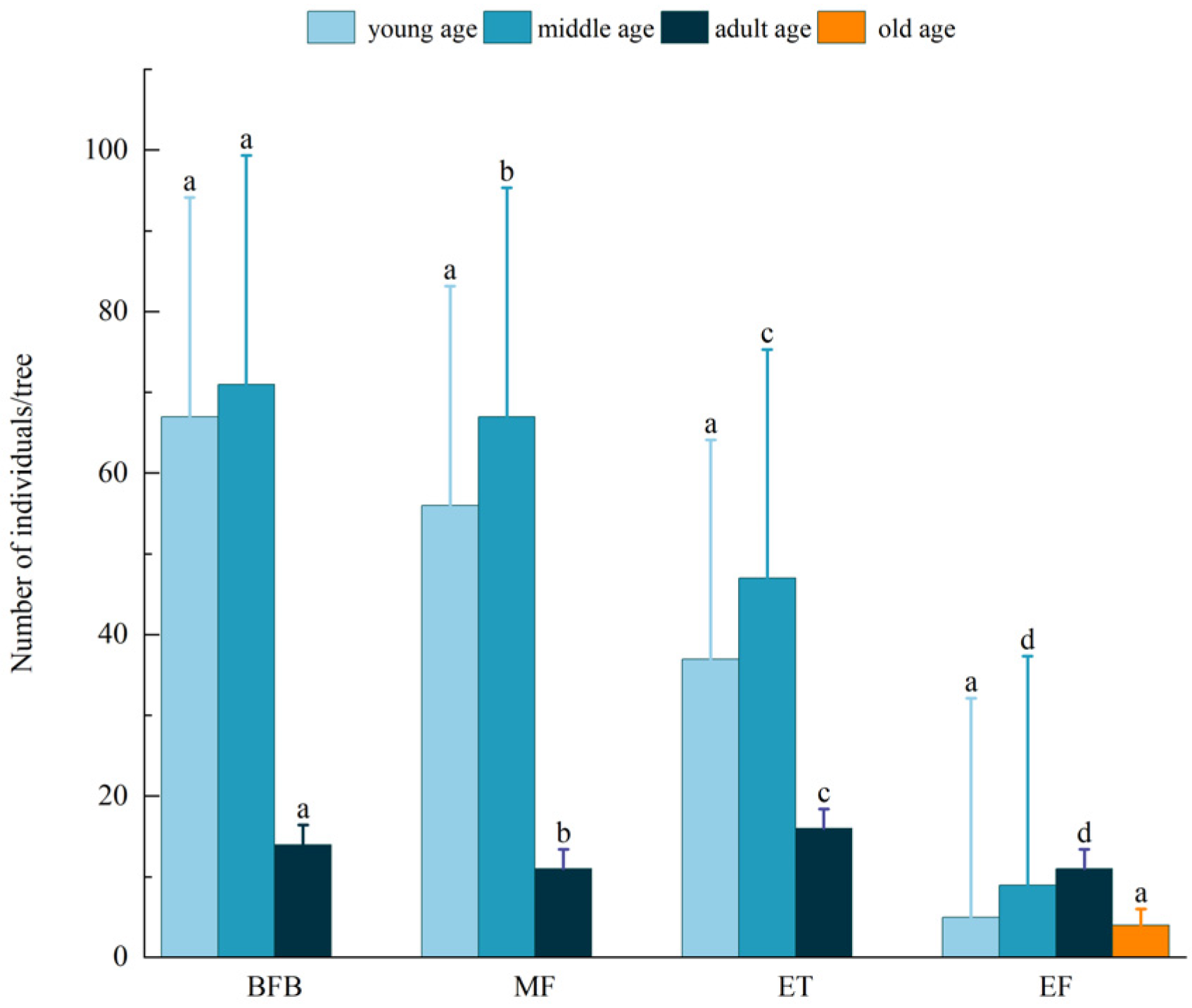
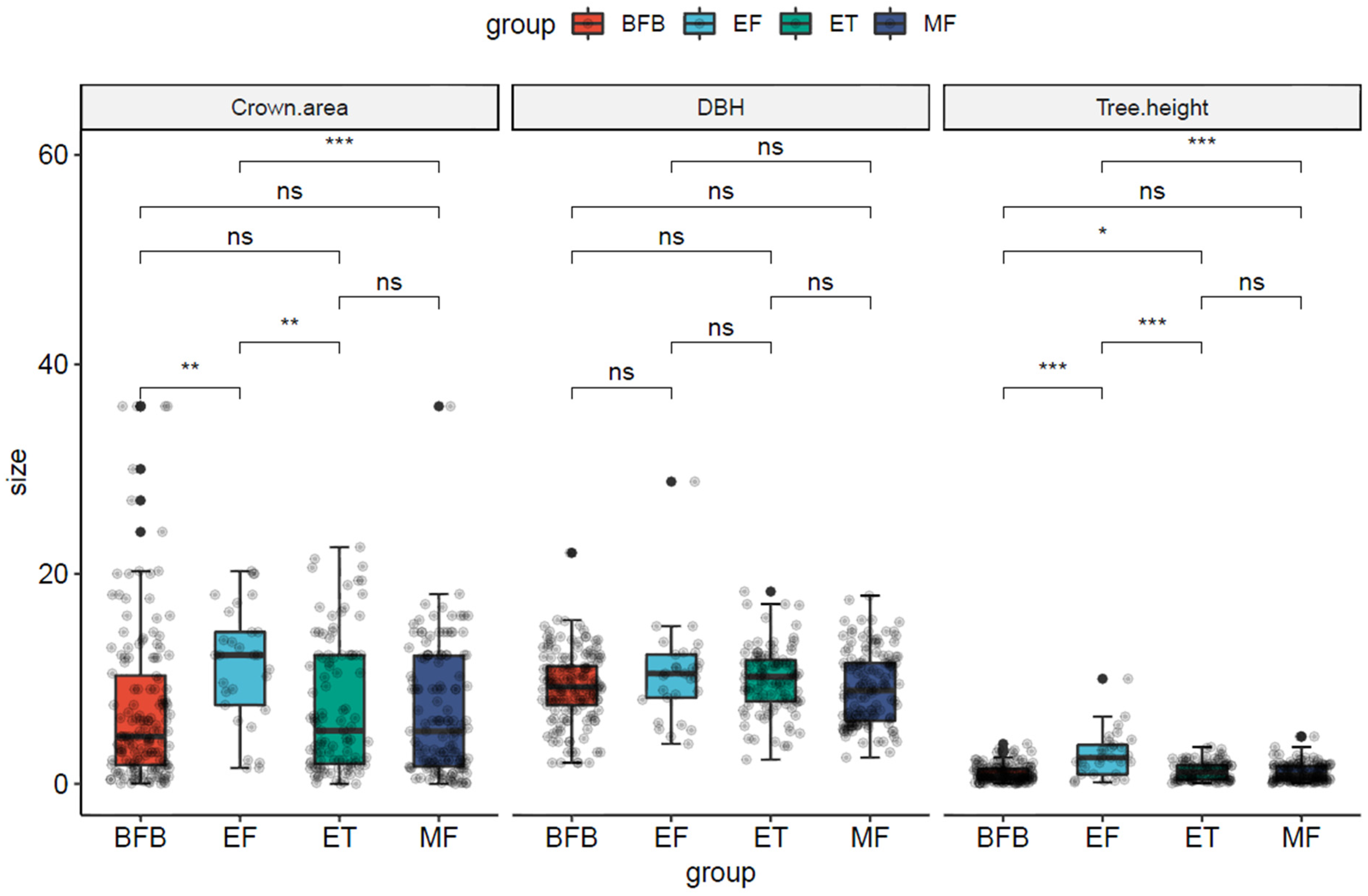
3.2. Structural Characteristics of A. spinulosa Populations under Different Transects
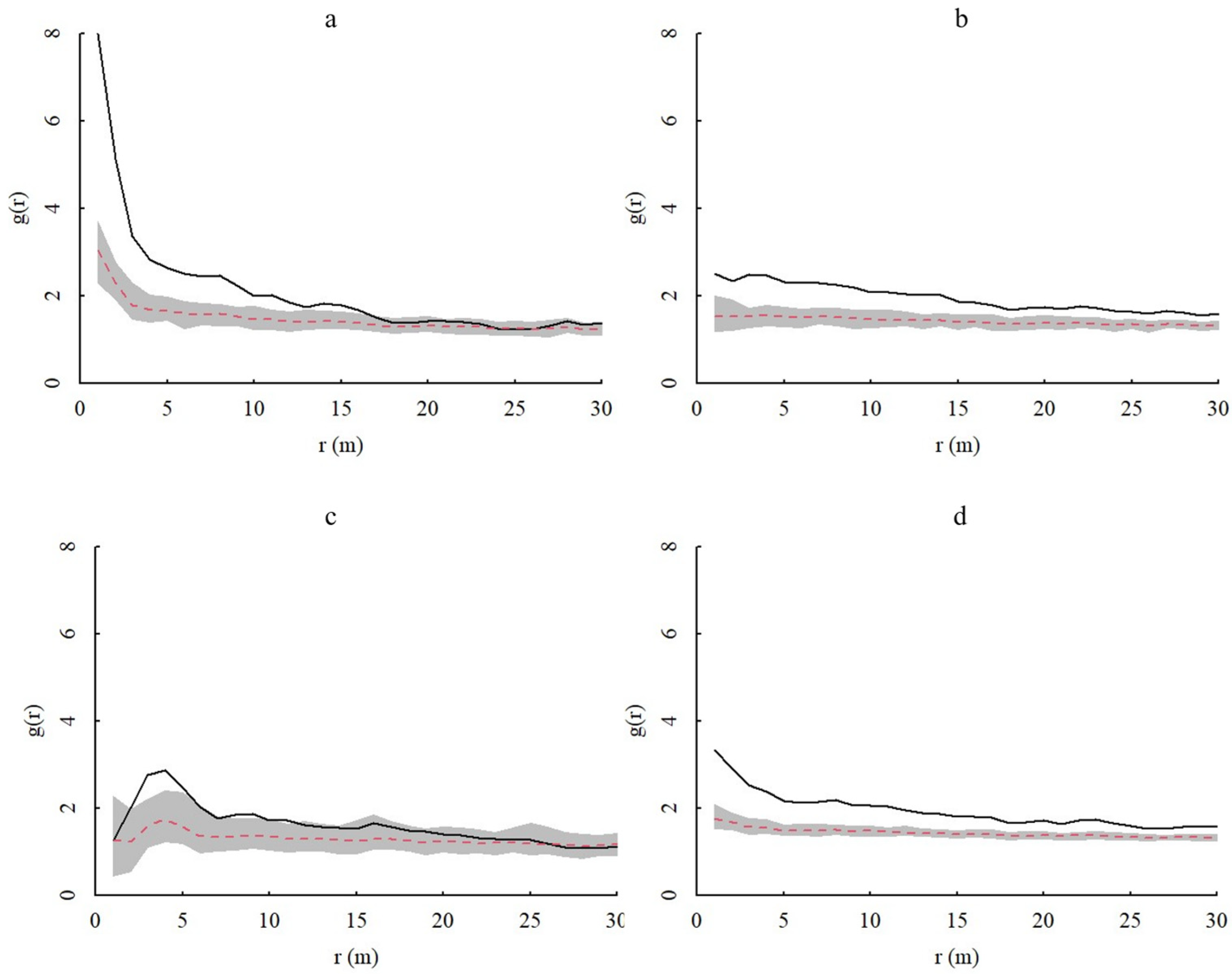
3.3. Spatial Patterns of Different Age Classes of A. spinulosa Populations in Chishui and Their Intraspecific Associations
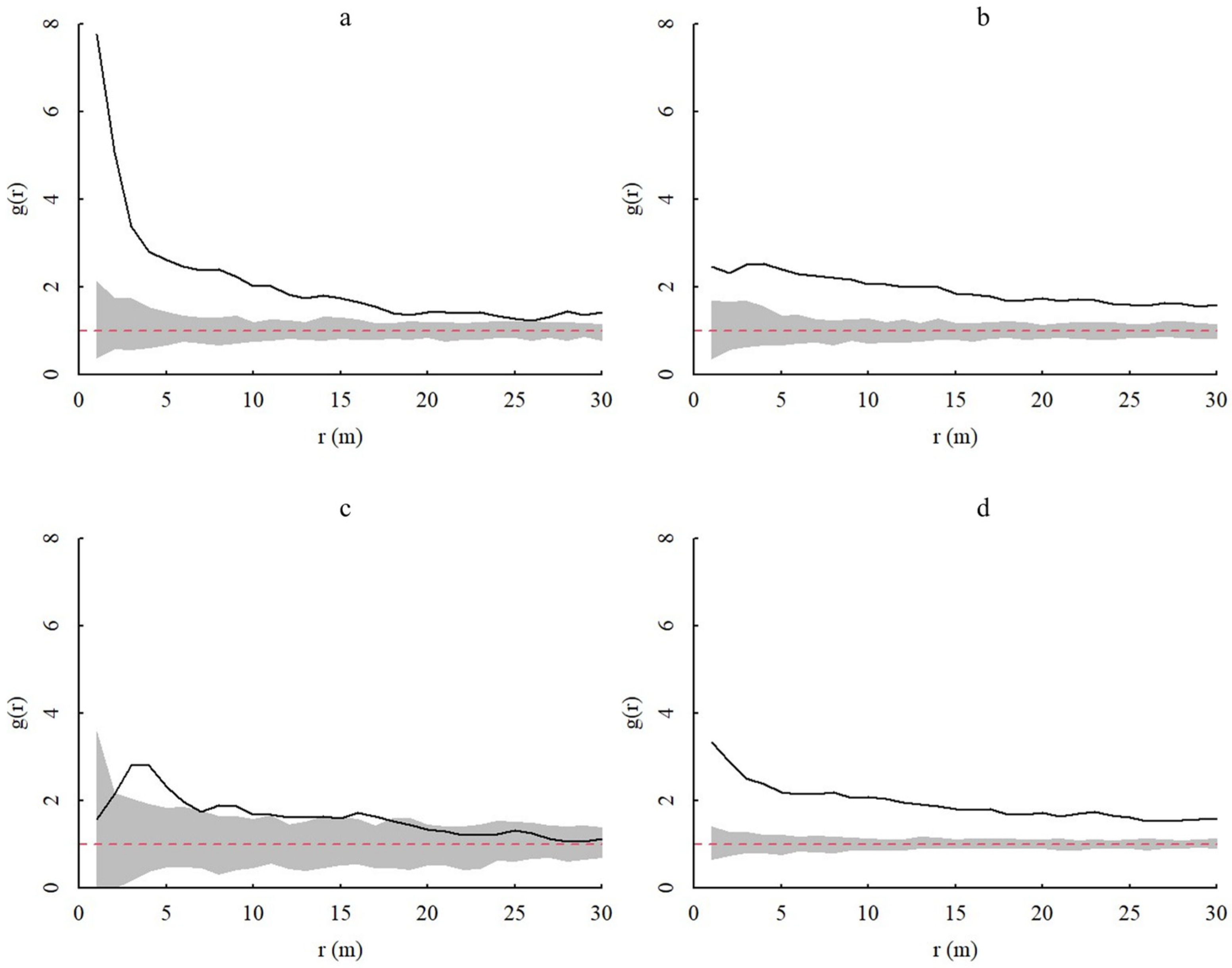
3.3.1. Spatial Distributions Based on the CSR and HP Null Models
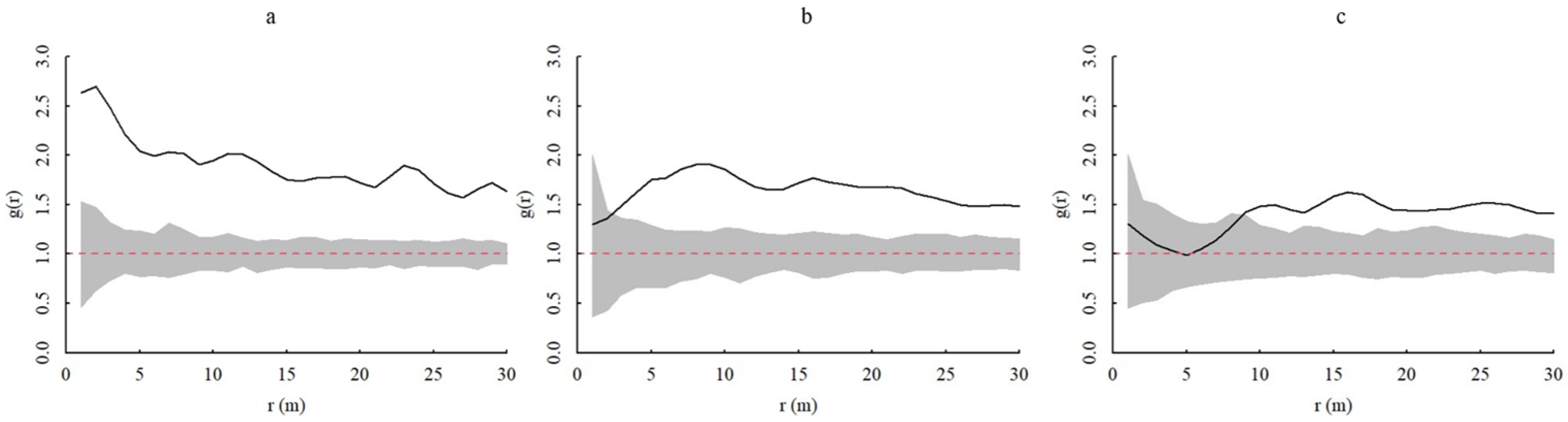
3.3.2. Spatial Correlation between Different Age Classes of A. spinulosa Populations
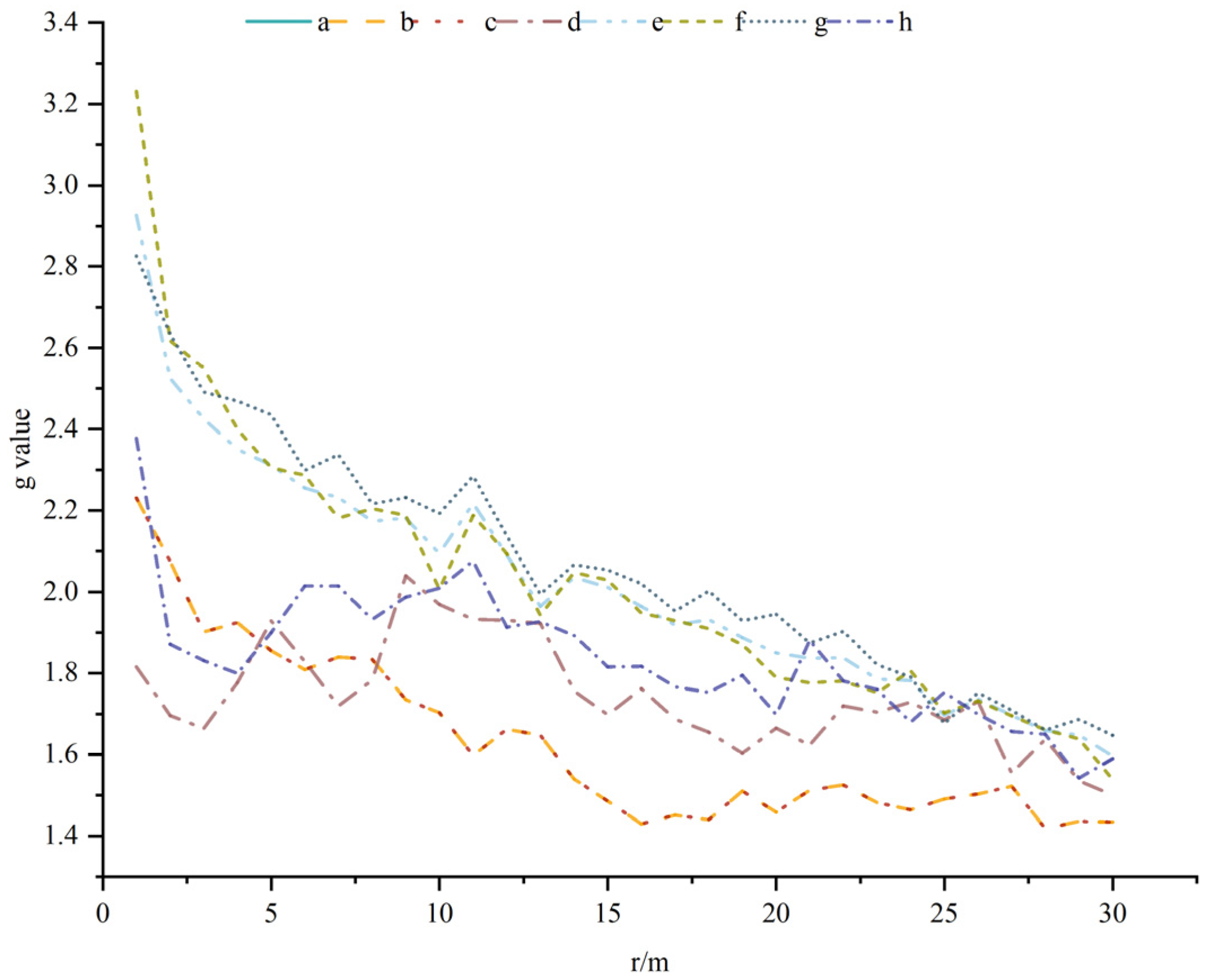
3.4. Spatial Association of Different Forest Layer Plants with A. spinulosa Populations
3.5. Association of Dominant Plants with A. spinulosa Populations
4. Discussion
4.1. Influence of Community Type and Habitat Heterogeneity on A. spinulosa Population Structures
4.2. Spatial Relationships of A. spinulosa
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peng, C.; Ai, W.; Qi, L.; Tu, J.; Meng, Y.; Yang, M.; Li, M. Cunninghamia lanceolata sprout population structure, dynamics and biomass allocation changes during Phyllostachys edulis expansion in middleō-southern China. Trees 2022, 36, 1207–1218. [Google Scholar] [CrossRef]
- Acquah, S.B.; Marshall, P.L.; Eskelson, B.N.; Barbeito, I. Temporal changes in tree spatial patterns in uneven-aged interior Douglas-fir dominated stands managed under different thinning treatments. For. Ecol. Manag. 2023, 528, 120640. [Google Scholar] [CrossRef]
- Wan, Z.B.; Li, X.Q.; Liu, C.G.; Sun, H.G.; Wang, X.F.; Fang, Y.J. Dynamic Analysis of Relationship among Tree Height, Diameter at the Breast Height and Crown of Artificial Sassafras tzumu Forest at Different Development Stages. Genom. Appl. Biol. 2017, 36, 331–339. [Google Scholar]
- Fibich, P.; Lepš, J.; Novotný, V.; Klimeš, P.; Těšitel, J.; Molem, K.; Damas, K.; Weiblen, G.D. Spatial patterns of tree species distribution in New Guinea primary and secondary lowland rain forest. J. Veg. Sci. 2016, 27, 328–339. [Google Scholar] [CrossRef]
- Dray, S.; Pélissier, R.; Couteron, P.; Fortin, M.J.; Legendre, P.; Peres-Neto, P.R.; Bellier, E.; Bivand, R.; Blanchet, F.G.; De Cáceres, M.; et al. Community ecology in the age of multivariate multiscale spatial analysis. Ecol. Monogr. 2012, 82, 257–275. [Google Scholar] [CrossRef]
- Stuber, E.F.; Carlson, B.S.; Jesmer, B.R. Spatial personalities: A meta-analysis of consistent individual differences in spatial behavior. Behav. Ecol. 2022, 33, 477–486. [Google Scholar] [CrossRef]
- Liu, Y.; Zeng, Y.; Yang, Y.; Wang, N.; Liang, Y. Competition, spatial pattern, and regeneration of Haloxylon ammoden dron and Haloxylon persicum communities in the Gurbantunggut Desert, Northwest China. J. Arid. Land 2022, 14, 1138–1158. [Google Scholar] [CrossRef]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Lefcheck, J.S.; Byrnes, J.E.K.; Isbell, F.; Gamfeldt, L.; Griffin, J.N.; Eisenhauer, N.; Hensel, M.J.S.; Hector, A.; Cardinale, B.J.; Duffy, J.E. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Nat. Commun. 2015, 6, 6936. [Google Scholar] [CrossRef]
- Yan, N.; Zhang, H.; Zhang, Z.; Zhang, H.; Zhou, L.; Chen, T.; Feng, S.; Ding, C.; Yuan, M. The extraction, antioxidant and against β-amyloid induced toxicity of polyphenols from Alsophila spinulosa leaves. Arab. J. Chem. 2022, 15, 103707. [Google Scholar] [CrossRef]
- He, Q.Q.; Weng, T.; Liu, B.Y.; Luo, X.H.; Mu, J.; Cheng, J.; An, M.T.; Yan, L.B. Research Advance in Conservation Strategies of Cyatheaceae Plant Communities based on Interspecific Relationships. J. Trop. Subtrop. Bot. 2022, 30, 753–762. [Google Scholar]
- Chen, Z.Y.; Zhang, F.L.; Lan, Q.Y.; Xu, Z.F.; Tao, G.D. Study on the propagatiom and conservation of germplasm in Alsophila spinulosa. Acta Bot. Yunnanica 1991, 13, 181–188. [Google Scholar]
- Lang, Y.T.; Ma, C.W.; Yu, J.; Buo, R.; Yan, Y.H.; Yang, D.M. Technology for Rapid Propagation in Vitro of Two Alsophila Species. Chin. J. Trop. Crops 2021, 42, 1579–1588. [Google Scholar]
- Xie, C.P.; Wang, C.L.; Wang, H.H.; Li, C.J.; Cheng, L. Population Structure and Dynami Characteristics of Alsophila spinulosa in Diaoluo Mountain Tropic Rain Forest of Hainan. J. Xinyang Norm. Univ. (Nat. Sci. Ed.) 2022, 35, 566–575. [Google Scholar]
- Xie, C.P.; Zhao, B.S.; Liu, D.W.; Fang, Y. Study on the Population Structure of Alsophila podophylla Hook in Bawangling Nature Reserve. J. Sichuan Agric. Univ. 2018, 36, 765–771. [Google Scholar]
- Yang, Q.J. Simulating the Potential Suitable Habitats of the Relict Plant Alsophila spinulosa in Different Periods Based on MaxEnt Model; Zhejisng University: Hangzhou, China, 2021. [Google Scholar]
- Ji, Y.; Cao, M.Y.; Bai, C.F.; Jing, L.Q.; Li, C.; Li, P. Population Structure and Dynamics of Alsophila spinulosa in Mount Emei. Acta Bot. Boreali Occident. Sin. 2019, 39, 543–551. [Google Scholar]
- Zhao, R.B. Study on Population Structure and Distribution Characteristics of Wild Cyatheaceae in Hainan Island; Hainan University: Haikou, China, 2018. [Google Scholar]
- Hu, Q.; Qian, R.; Zhang, Y.; Ma, X.; Ye, Y.; Zhang, X.; Lin, L.; Liu, H.; Zheng, J. Complete chloroplast genome molecular structure, comparative and phylogenetic analyses of Sphaeropteris lepifera of Cyatheaceae family: A tree fern from China. Sci. Rep. 2023, 13, 1356. [Google Scholar] [CrossRef]
- Du, Y. Study on Population Dynamics and Host Selection of Phytophagous Insects of Alsophila spinulosa; Guizhou Normal University: Guiyang, China, 2022. [Google Scholar]
- Zuo, Y.; Qu, H.; Xia, C.; Zhang, H.; Zhang, J.; Deng, H. Moso Bamboo Invasion Reshapes Community Structure of Denitrifying Bacteria in Rhizosphere of Alsophila spinulosa. Microorganisms 2022, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Jiao, P.; Qi, Z.; Jiang, Z.; Guan, S. Characterization of the complete chloroplast genome of Alsophila spinulosa, an endangered species endemic to China. Mitochondrial DNA Part B 2020, 5, 2262–2263. [Google Scholar] [CrossRef]
- Zhong, X.H. A Tentative Study on Community Characteristics and Population Dynamics of Alsophila spinulata of Chishui Alsophila spinulata National Nature Reserve; Southwest University: Chongqing, China, 2017. [Google Scholar]
- Jiao, J.; Hu, G.; Yan, J. Heterogeneity pursuit for spatial point pattern with application to tree locations: A Bayesian semipar ametric recourse. Environmetrics 2021, 32, e2694. [Google Scholar] [CrossRef]
- Wang, X.; Wang, D.; Li, H.; Tai, Y.; Jiang, C.; Liu, F.; Li, S.; Miao, B. Cumulative effects of K-function in point patterns. Chin. J. Appl. Ecol. 2022, 33, 1275–1282. [Google Scholar]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Miao, N.; Xu, H.; Moermond, T.C.; Li, Y.; Liu, S. Density-dependent and distance-dependent effects in a 60-ha tropical moun tain rain forest in the Jianfengling mountains, Hainan Island, China: Spatial pattern analysis. For. Ecol. Manag. 2018, 429, 226–232. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Zhang, L.; Feng, L.; Yan, L.; Li, F.; Zhao, X.; Yu, L.; Liu, N. Relationship between diversity and stability of a karst plant community. Ecol. Evol. 2022, 12, e9254. [Google Scholar] [CrossRef]
- Wang, X.T.; Hou, Y.L.; Liang, C.Z.; Wang, W.; Liu, F. Point pattern analysis based on different null models for detecting spatial patterns. Biodivers. Sci. 2012, 20, 151–158. [Google Scholar]
- Jiao, J.; Wu, C.; Jiang, B.; Wang, Z.; Yuan, W.; Zhu, J.; Li, T.; Yang, S.; Yao, L. Negative Density Restricts the Coexistence and Spatial Distribution of Dominant Species in Subtropical Evergreen Broad-Leaved Forests in China. Forests 2022, 13, 1227. [Google Scholar] [CrossRef]
- Shi, W.; Wang, Y.Q.; Xiang, W.S.; Li, X.K.; Cao, K.F. Environmental filtering and dispersal limitation jointly shaped the taxonomic and phylogenetic beta diversity of natural forests in southern China. Ecol. Evol. 2021, 11, 8783–8794. [Google Scholar] [CrossRef]
- Wu, H.; Lei, J.; Li, X.; Wang, H.; Duan, A.; Zhang, J. Aggregation distributions across stand age in provenances of Cunninghamia lanceolata (Lamb.) Hook. For. Ecol. Manag. 2021, 494, 119317. [Google Scholar] [CrossRef]
- Yusup, A.; Halik, Ü.; Abliz, A.; Aishan, T.; Keyimu, M.; Wei, J. Population Structure and Spatial Distribution Pattern of Populus euphratica Riparian Forest under Environmental Heterogeneity along the Tarim River, Northwest China. Front. Plant Sci. 2022, 13, 844819. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N.; Okuda, T. Analyzing the Spatial Structure of a Sri Lankan Tree Species with Multiple Scales of Clustering. Ecology 2007, 88, 3088–3102. [Google Scholar] [CrossRef]
- Bland, M.; Baldocchi, D.D. Vertical structure heterogeneity in broadleaf forests: Effects on light interception and canopy photosynthesis. Agric. For. Meteorol. 2021, 307, 108525. [Google Scholar] [CrossRef]
- Song, Y.; Yan, G.; Zhang, G. Light Competition Contributes to the Death of Masson Pines of Coniferous-Broadleaf Mixed Forests in Subtropical China. Forests 2022, 13, 85. [Google Scholar] [CrossRef]
- Dodonov, P.; Harper, K.A. Spatial patterns of structural diversity across the boreal forest-tundra ecotone in Churchill, Canada. Acta Oecologica 2022, 117, 103862. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transect | Slope Position | Altitude/m | Slope | Stand Density/(Plant/m2) | Alsophila spinulosa Density/(Plant/m2) | Community Height/m | Number of Species | Dominant Species |
|---|---|---|---|---|---|---|---|---|
| BFB | Downhill position | 526 | 16.32 | 0.432857 | 0.054286 | 7.91 ± 5.45 | 34 | Phyllostachys heterocycla, Brassaiopsis glomerulata, A. spinulosa, Bridelia tomentosa, Ficus virens |
| MF | Downhill position | 526 | 16.84 | 0.548929 | 0.047857 | 6.99 ± 5.23 | 49 | Phyllostachys heterocycla, Brassaiopsis glomerulata, Bridelia tomentosa, A. spinulosa, Ficus virens |
| ET | Medium position | 551 | 41.99 | 0.564286 | 0.035714 | 7.00 ± 4.88 | 37 | Phyllostachys heterocycla, Phyllostachys heterocycla, Brassaiopsis glomerulata, Maesa japonica, A. spinulosa |
| EF | Medium position | 551 | 44.76 | 0.4325 | 0.010357 | 4.08 ± 3.41 | 37 | Mallotus philippensis, Maesa japonica, Cinnamomum glanduliferum, Diplospora dubia, Phyllostachys heterocycla |
| Species | Significant Values | Highly/m | Associations |
|---|---|---|---|
| Phyllostachys edulis | 33.05 | 12.09 ± 2.20 | (+) |
| Maesa japonica | 8.28 | 3.27 ± 2.04 | (+) |
| Bridelia retusa | 5.59 | 3.90 ± 2.56 | (+) |
| Brassaiopsis glomerulata | 5.20 | 3.00 ± 1.37 | (+) |
| Ficus virens | 3.53 | 4.56 ± 2.19 | (+) |
| Diplospora dubia | 3.01 | 3.04 ± 1.71 | 0~8 m scale (0), under the rest of the scale (+) |
| Symplocos cochinchinensis | 2.94 | 4.72 ± 3.93 | (+) |
| Mallotus philippensis | 2.68 | 3.08 ± 1.95 | 0~21 m scale (−), under the rest of the scale (0) |
| Mallotus barbatus | 2.36 | 3.89 ± 1.78 | (+) |
| Dichroa febrifuga | 2.28 | 2.06 ± 1.09 | (+) |
| Eurya kueichowensis | 2.24 | 3.04 ± 2.01 | (+) |
| Musa basjoo | 2.02 | 4.29 ± 2.72 | 0~26 m scale (+), under the rest of the scale (0) |
| Toxicodendron sylvestre | 1.95 | 4.88 ± 3.57 | (+) |
| Cinnamomum glanduliferum | 1.86 | 3.28 ± 1.62 | (+) |
| Eurya nitida | 1.62 | 3.36 ± 1.90 | 0~24 m scale (−), under the rest of the scale (0) |
| Debregeasia orientalis | 1.46 | 1.70 ± 0.46 | (+) |
| Lasianthus chinensis | 1.02 | 1.61 ± 0.20 | 4.5~8 m scale (−), 24~29 m scale (+), under the rest of the scale (0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, D.; Yang, M.; Yu, L.; An, M.; He, Q.; Mu, J.; Yan, L. Population Structure, Distribution, and Spatial Characteristics of Alsophila spinulosa in Chishui, China. Diversity 2023, 15, 1200. https://doi.org/10.3390/d15121200
Yuan D, Yang M, Yu L, An M, He Q, Mu J, Yan L. Population Structure, Distribution, and Spatial Characteristics of Alsophila spinulosa in Chishui, China. Diversity. 2023; 15(12):1200. https://doi.org/10.3390/d15121200
Chicago/Turabian StyleYuan, Dongmei, Man Yang, Lifei Yu, Mingtai An, Qinqin He, Jun Mu, and Lingbin Yan. 2023. "Population Structure, Distribution, and Spatial Characteristics of Alsophila spinulosa in Chishui, China" Diversity 15, no. 12: 1200. https://doi.org/10.3390/d15121200
APA StyleYuan, D., Yang, M., Yu, L., An, M., He, Q., Mu, J., & Yan, L. (2023). Population Structure, Distribution, and Spatial Characteristics of Alsophila spinulosa in Chishui, China. Diversity, 15(12), 1200. https://doi.org/10.3390/d15121200