
Pool Age and Local Habitat Are Associated with Effective Number of Breeders in Spotted Salamander Populations Colonizing Created Vernal Pools




Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Field Sampling
2.3. Genetic Analysis
2.4. Population Structure
2.5. Effective Number of Breeders and Genetic Diversity
2.6. Environmental Factors
3. Results
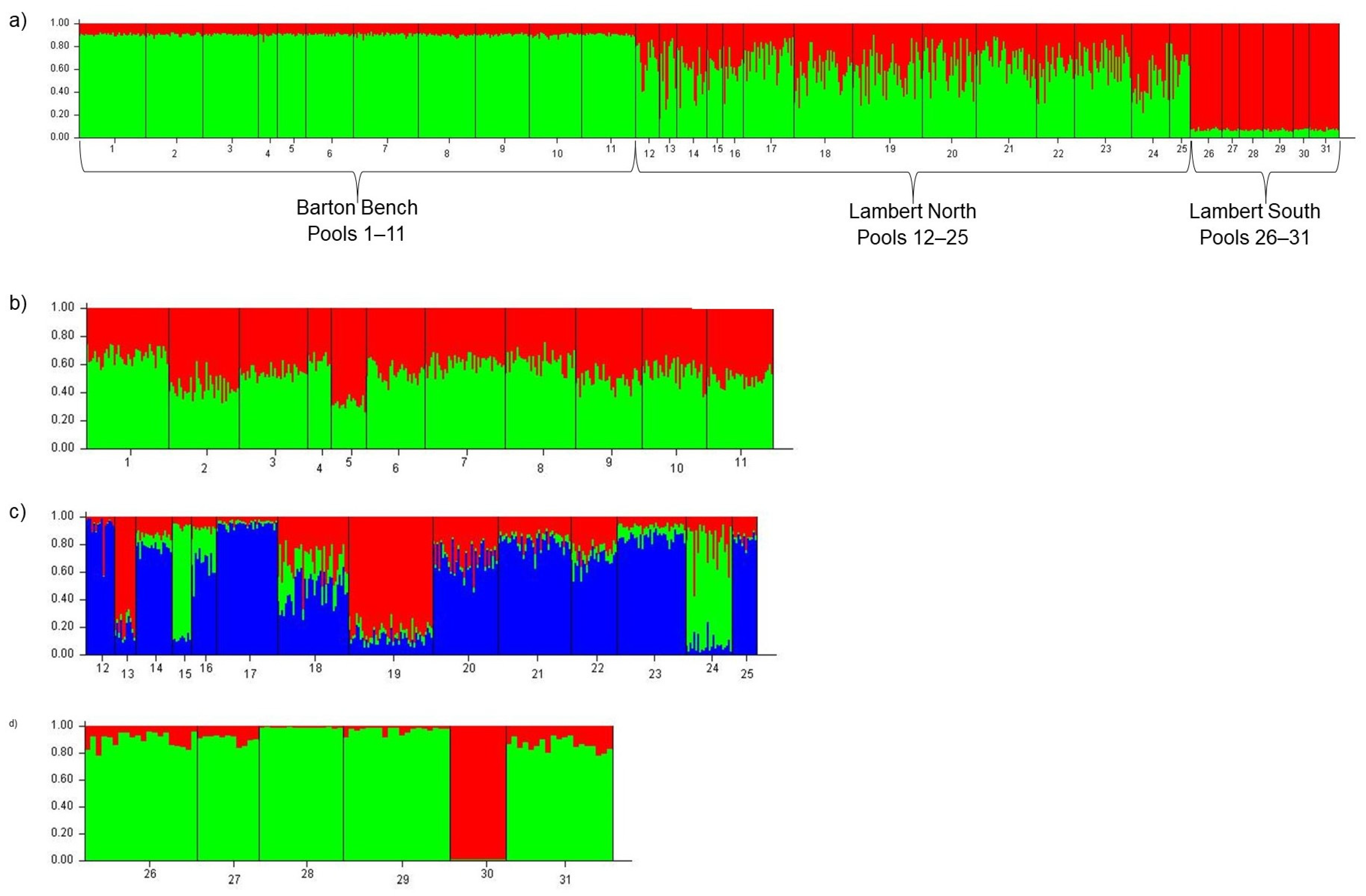
3.1. Population Structure
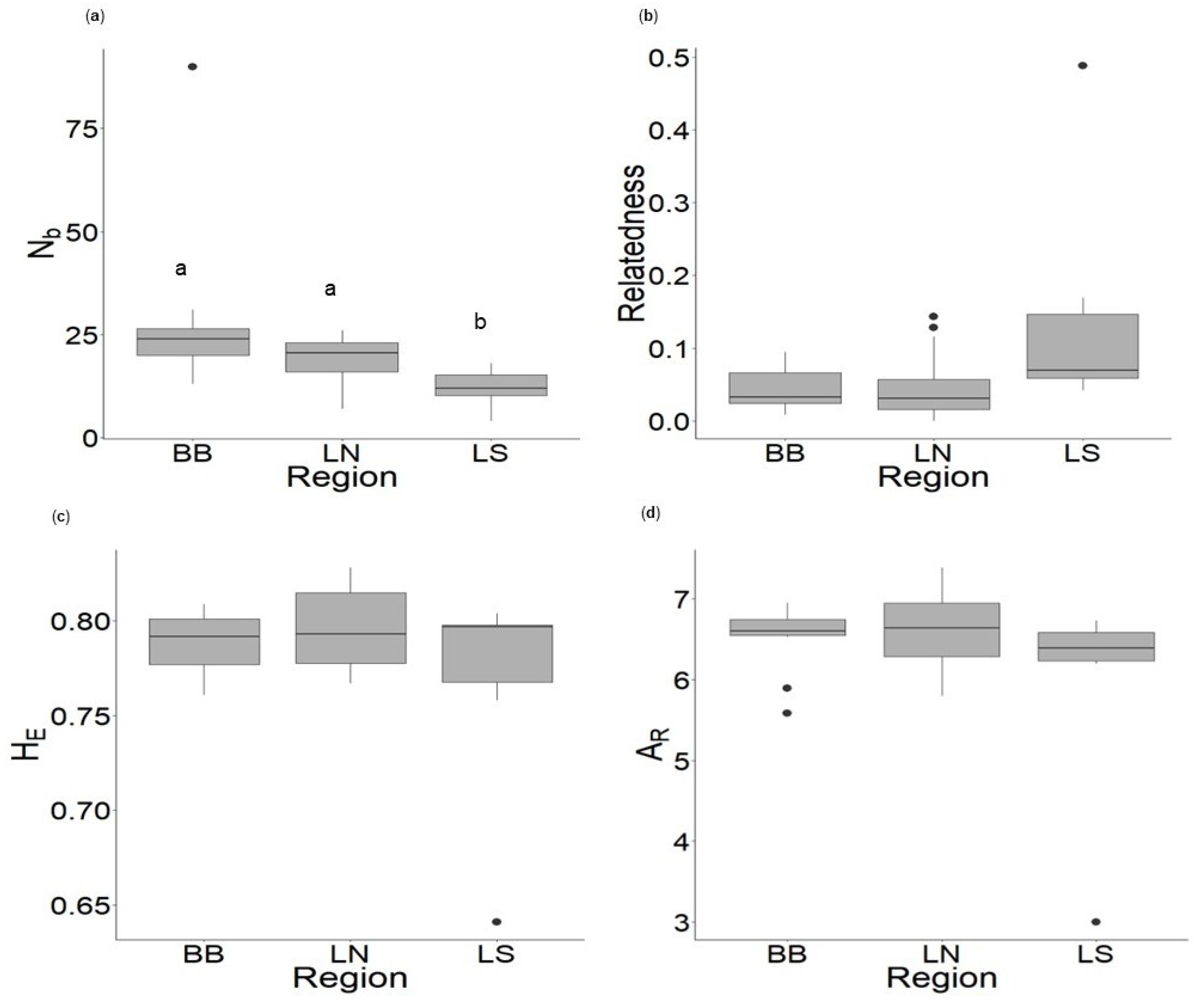
3.2. Effective Number of Breeders and Genetic Diversity
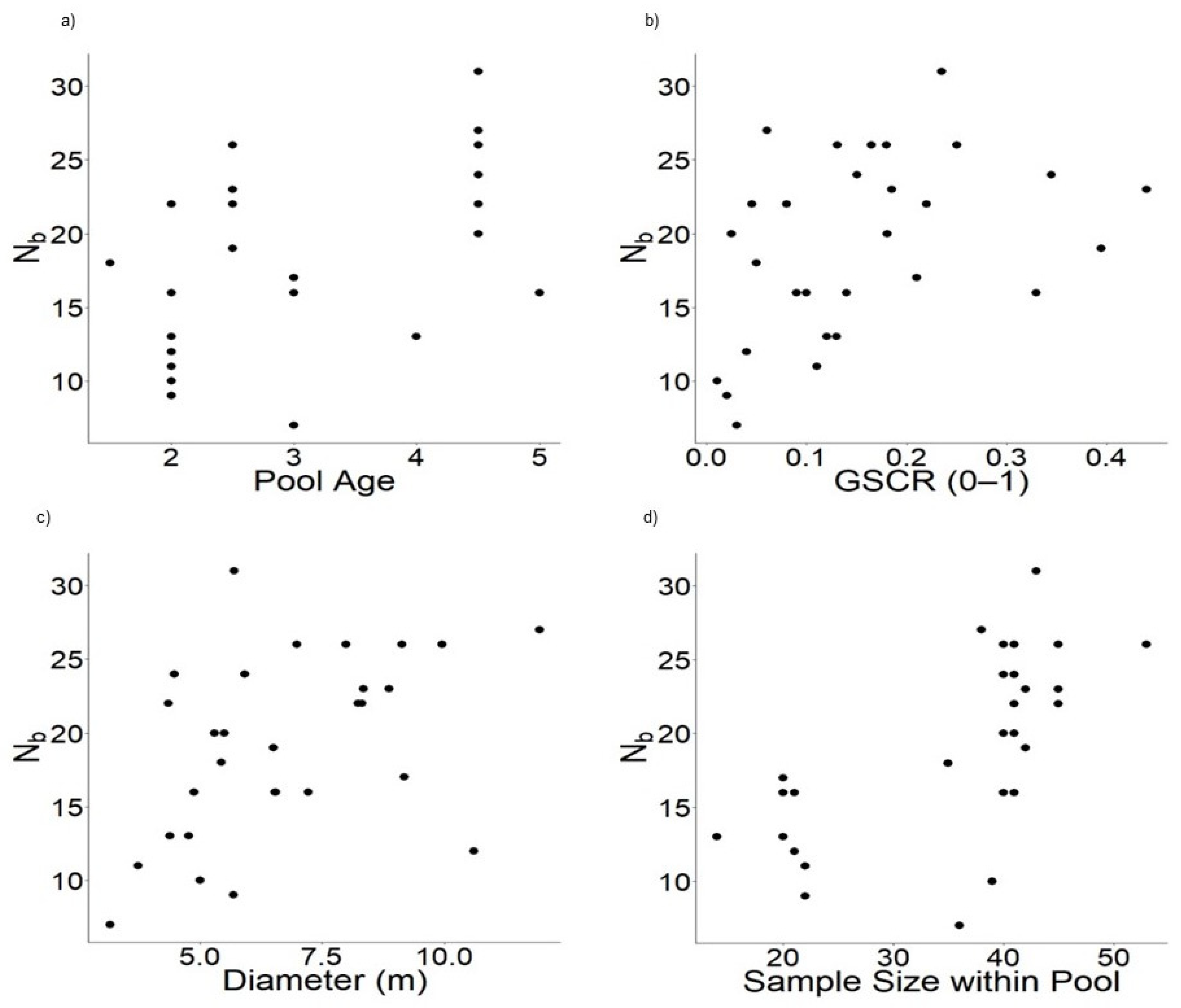
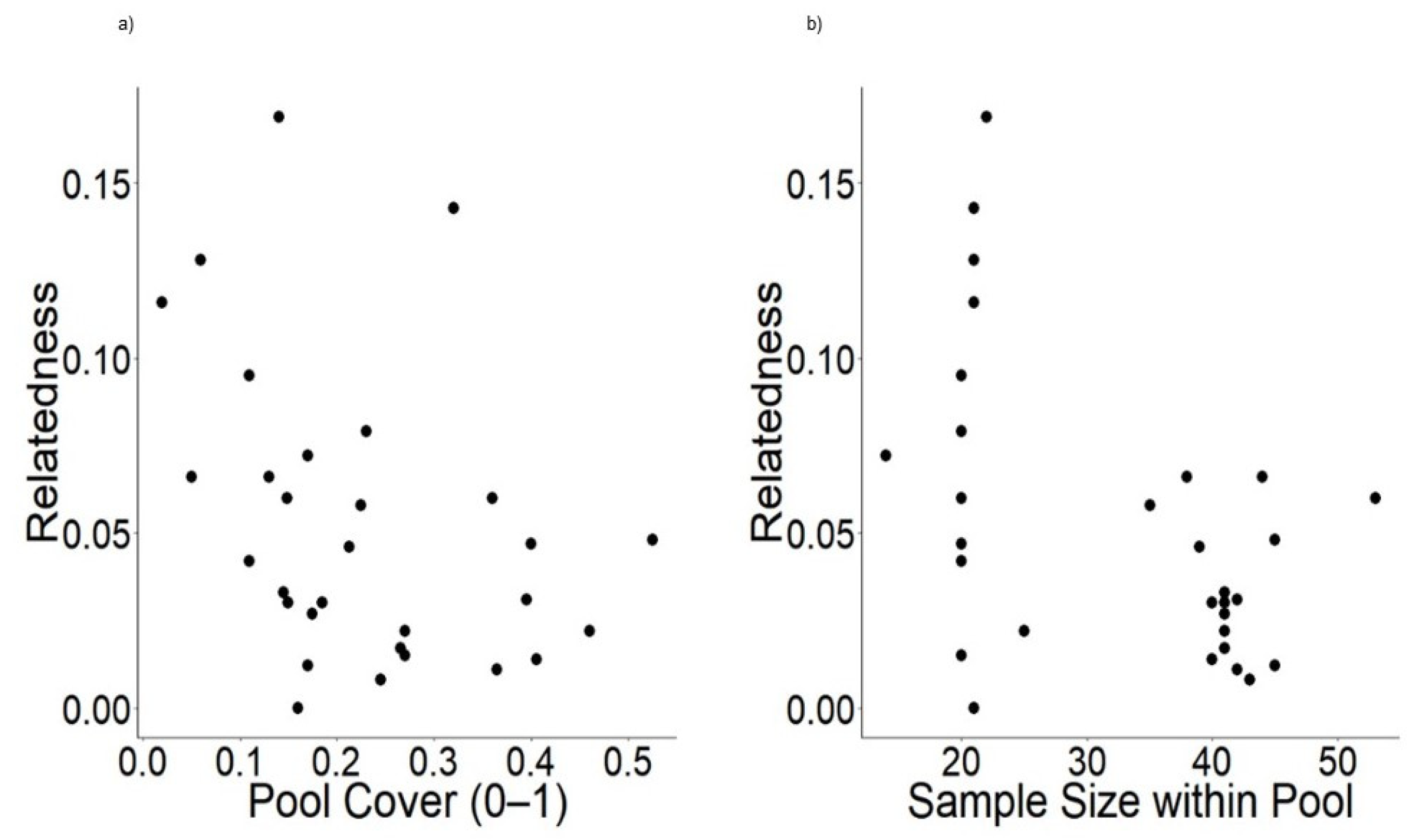
3.3. Environmental Factors
4. Discussion
4.1. Population Structure
4.2. Effective Number of Breeders and Genetic Diversity
4.3. Environmental Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calhoun, A.J.K.; Mushet, D.M.; Bell, K.P.; Boix, D.; Fitzsimons, J.A.; Isselin-Nondedeu, F. Temporary wetlands: Challenges and solutions to conserving a ‘disappearing’ ecosystem. Biol. Conserv. 2017, 211, 3–11. [Google Scholar] [CrossRef]
- Mushet, D.M.; Calhoun, A.J.K.; Alexander, L.C.; Cohen, M.J.; DeKeyser, E.S.; Fowler, L.; Lane, C.R.; Lang, M.W.; Rains, M.C.; Walls, S.C. Geographically isolated wetlands: Rethinking a misnomer. Wetlands 2015, 35, 423–431. [Google Scholar] [CrossRef] [Green Version]
- DiBello, F.J.; Calhoun, A.J.; Morgan, D.E.; Shearin, A.F. Efficiency and detection accuracy using print and digital stereo aerial photography for remotely mapping vernal pools in New England landscapes. Wetlands 2016, 36, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Calhoun, A.J.K.; Arrigoni, J.; Brooks, R.P.; Hunter, M.L.; Richter, S.C. Creating successful vernal pools: A literature review and advice for practitioners. Wetlands 2014, 34, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Spielman, D.; Brook, B.W.; Frankham, R. Most species are not driven to extinction before genetic factors impact them. Proc. Natl. Acad. Sci. USA 2004, 101, 15261–15264. [Google Scholar] [CrossRef] [Green Version]
- Frankham, R. Genetics and extinction. Biol. Conserv. 2005, 126, 131–140. [Google Scholar] [CrossRef]
- Beebee, T.J.C. Conservation genetics of amphibians. Heredity 2005, 95, 423–427. [Google Scholar] [CrossRef]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Ruzzante, D.E.; McCracken, G.R.; Parmelee, S.; Hill, K.; Corrigan, A.; MacMillan, J.L.; Walde, S.J. Effective number of breeders, effective population size and their relationship with census size in an iteroparous species, Salvelinus fontinalis. Proc. R. Soc. B 2016, 283, 20152601. [Google Scholar] [CrossRef] [Green Version]
- Wang, J. A new method for estimating effective population sizes from a single sample of multilocus genotypes. Mol. Ecol. 2009, 18, 2148–2164. [Google Scholar] [CrossRef]
- Waples, R.S.; Luikart, G.; Faulker, J.R.; Tallmon, D.A. Simple life-history traits explain key effective population size ratios across diverse taxa. Proc. R. Soc. B 2013, 280, 20131339. [Google Scholar] [CrossRef] [Green Version]
- McKee, A.M.; Maerz, J.C.; Smith, L.L.; Glenn, T.C. Habitat predictors of genetic diversity for two sympatric wetland-breeding amphibian species. Ecol. Evol. 2017, 7, 6271–6283. [Google Scholar] [CrossRef]
- Costanzi, J.-M.; Mège, P.; Boissinot, A.; Isselin-Nondedeu, F.; Guérin, S.; Lourdais, O.; Trochet, A.; Le Petitcorps, Q.; Legrand, A.; Varenne, F.; et al. Agricultural landscapes and the Loire River influence the genetic structure of the Marbled Newt in Western France. Sci. Rep. 2018, 8, 14177. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, B.J.; Phillips, C.A.; Schooley, R.L.; Lowe, W.H.; Douglas, M.R. Linking extinction-colonization dynamics to genetic structure in a salamander metapopulation. Proc. R. Soc. B 2012, 279, 1575–1582. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, D.; Calhoun, A.J.K. Monitoring created seasonal pools for functional success: A six-year case study of amphibian responses, Sears Island, Maine, USA. Wetlands 2006, 26, 992–1003. [Google Scholar] [CrossRef]
- Denton, R.D.; Richter, S.C. Amphibian communities in natural and constructed ridge top wetlands with implications for wetland construction. J. Wildl. Manag. 2013, 77, 886–896. [Google Scholar] [CrossRef]
- Egan, R.S.; Paton, P.W.C. Within-pond parameters affecting oviposition by wood frogs and spotted salamanders. Wetlands 2004, 24, 1–13. [Google Scholar] [CrossRef]
- Petranka, J.W.; Kennedy, C.A.; Murray, S.S. Response of amphibians to restoration of a southern Appalachian wetland: A long-term analysis of community dynamics. Wetlands 2003, 23, 1030–1042. [Google Scholar] [CrossRef]
- Skidds, D.E.; Golet, F.C.; Paton, P.W.; Mitchell, J.C. Habitat correlates of reproductive effort in wood frogs and spotted salamanders in an urbanizing watershed. J. Herpetol. 2007, 41, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Felix, Z.I.; Wang, Y.; Schweitzer, C.J. Effects of experimental canopy manipulation on amphibian egg deposition. J. Wildl. Manag. 2010, 74, 496–503. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Furman, B.L.S.; Evans, J.P. Salamanders continue to breed in ephemeral ponds following the removal of surrounding terrestrial habitat. Herpetol. Conserv. Biol. 2013, 8, 715–723. [Google Scholar]
- Petranka, J.W.; Murray, S.S.; Kennedy, C.A. Responses of amphibians to restoration of a southern Appalachian wetland: Perturbations confound post-restoration assessment. Wetlands 2003, 23, 278–290. [Google Scholar] [CrossRef]
- Canty, J.L. Weatherbase. Available online: http://weatherbase.com (accessed on 22 June 2018).
- Millikin, A.R.; Davis, D.R.; Brown, D.J.; Woodley, S.K.; Coster, S.; Welsh, A.; Kerby, J.L.; Anderson, J.T. Prevalence of ranavirus in spotted salamander larvae from created vernal pools in West Virginia, USA. J. Wildl. Dis. 2023. [Google Scholar] [CrossRef]
- Millikin, A.R.; Woodley, S.K.; Davis, D.R.; Anderson, J.T. Habitat characteristics in created vernal pools impact spotted salamander water-borne corticosterone levels. Wetlands 2019, 39, 803–814. [Google Scholar] [CrossRef]
- Barbour, M.G.; Burk, J.H.; Pitts, W.D.; Gilliam, F.S.; Schwartz, M.W. Terrestrial Plant Ecology; Benjamin and Cummings: Menlo Park, CA, USA, 1999. [Google Scholar]
- Julian, S.E.; King, T.L.; Savage, W.K. Isolation and characterization of novel tetranucleotide microsatellite DNA markers for the spotted salamander, Ambystoma maculatum. Mol. Ecol. Resour. 2003, 3, 7–9. [Google Scholar] [CrossRef]
- Blacket, M.J.; Robin, C.; Good, R.T.; Lee, S.F.; Miller, A.D. Universal primers for fluorescent labeling of PCR fragments—An efficient and cost-effective approach to genotyping by fluorescence. Mol. Ecol. Resour. 2012, 12, 456–463. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Resour. 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Wang, J. Sibship reconstruction from genetic data with typing errors. Genetics 2004, 166, 1963–1979. [Google Scholar] [CrossRef]
- Goldberg, C.S.; Waits, L.P. Quantification and reduction of bias from sampling larvae to infer population and landscape genetic structure. Mol. Ecol. Resour. 2010, 10, 304–313. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. FSTAT (Version 1.2): A computer program to calculate F-statistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Wang, I.J.; Johnson, J.R.; Johnson, B.B.; Shaffer, H.B. Effective population size is strongly correlated with breeding pond size in the endangered California tiger salamander, Ambystoma californiense. Conserv. Genet. 2011, 12, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research–an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Queller, D.C.; Goodnight, K.F. Estimating relatedness using genetic markers. Evolution 1989, 43, 258–275. [Google Scholar] [CrossRef]
- Dinno, A. Dunn.Test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 17 July 2017).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2017. Available online: https://www.R-project.org/ (accessed on 15 July 2017).
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R Package Version 2.1-1. 2017. Available online: https://cran.r-project.org/package=AICcmodavg (accessed on 15 July 2017).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer Science & Business Media, LLC.: New York, NY, USA, 2002. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis. A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre (ICRAF): Nairobi, Kenya, 2005; ISBN 92-9059-179-X. [Google Scholar]
- Komsta, L. Outliers: Tests for Outliers. R Package Version 0.14. 2011. Available online: https://CRAN.R-project.org/package=outliers (accessed on 20 July 2017).
- Coster, S.S.; Babbitt, K.J.; Cooper, A.; Kovach, A.I. Limited influence of local and landscape factors on finescale gene flow in two pond-breeding amphibians. Mol. Ecol. 2015, 24, 742–758. [Google Scholar] [CrossRef]
- Montieth, K.E.; Paton, P.W.C. Emigration behavior of spotted salamanders on golf courses in southern Rhode Island. J. Herpetol. 2006, 40, 195–205. [Google Scholar] [CrossRef]
- Wilbur, H.M. Propagule size, number, and dispersion pattern in Ambystoma and Asclepias. Am. Nat. 1977, 111, 43–68. [Google Scholar] [CrossRef]
- Flageole, S.; Leclair, R. Demography of a salamander (Ambystoma maculatum) population studied by skeletochronology. Can. J. Zool. 1992, 70, 740–749. [Google Scholar] [CrossRef]
- Cornuet, J.-M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef] [PubMed]
- Williamson-Natesan, E.G. Comparison of methods for detecting bottlenecks from microsatellite loci. Conserv. Genet. 2005, 6, 551–562. [Google Scholar] [CrossRef]
- Peery, M.Z.; Kirby, R.; Reid, B.N.; Stoelting, R.; Doucet-Bëer, E.; Robinson, S.; Vásquez-Carrillo, C.; Pauli, J.N.; Palsbøll, P.J. Reliability of genetic bottleneck tests for detecting recent population declines. Mol. Ecol. 2012, 21, 3403–3418. [Google Scholar] [CrossRef]
- Whiteley, A.R.; McGarigal, K.; Schwartz, M.K. Pronounced differences in genetic structure despite overall ecological similarity for two Ambystoma salamanders in the same landscape. Conserv. Genet. 2014, 15, 573–591. [Google Scholar] [CrossRef]
- Richardson, J.L. Divergent landscape effects on population connectivity in two co-occurring amphibian species. Mol. Ecol. 2012, 21, 4437–4451. [Google Scholar] [CrossRef]
- Groff, L.A.; Loftin, C.S.; Calhoun, A.J.K. Predictors of breeding site occupancy by amphibians in montane landscapes. J. Wildl. Manag. 2017, 81, 269–278. [Google Scholar] [CrossRef]
- Rothenberger, M.B.; Vera, M.K.; Germanoski, D.; Ramirez, E. Comparing amphibian habitat quality and functional success among natural, restored, and created vernal pools. Restor. Ecol. 2019, 27, 881–891. [Google Scholar] [CrossRef]
- Cosentino, B.J.; Schooley, R.L.; Phillips, C.A. Spatial connectivity moderates the effect of predatory fish on salamander metapopulation dynamics. Ecosphere 2011, 2, 1–14. [Google Scholar] [CrossRef]
- Greenwald, K.R.; Gibbs, H.L.; Waite, T.A. Efficacy of land-cover models in predicting isolation of marbled salamander populations in a fragmented landscape. Conserv. Biol. 2009, 23, 1232–1241. [Google Scholar] [CrossRef]
- Lomolino, M.V. The target area hypothesis: The influence of island area on immigration rates of non-volant mammals. Oikos 1990, 57, 297–300. [Google Scholar] [CrossRef]
- Formanowicz, D.R.; Bobka, M.S. Predation risk and microhabitat preference: An experimental study of the behavioral responses of prey and predator. Am. Midl. Nat. 1989, 121, 379–386. [Google Scholar] [CrossRef]
- Kern, M.M.; Nassar, A.A.; Guzy, J.C.; Dorcas, M.E. Oviposition site selection by spotted salamanders (Ambystoma maculatum) in an isolated wetland. J. Herpetol. 2013, 47, 445–449. [Google Scholar] [CrossRef]
- Peterman, W.E.; Anderson, T.L.; Drake, D.L.; Ousterhout, B.H.; Semlitsch, R.D. Maximizing pond biodiversity across the landscape: A case study of larval Ambystomatid salamanders. Anim. Conserv. 2014, 17, 275–285. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | ΔAICc | wi | Deviance Explained | All Model Coefficients ≠ 0 |
|---|---|---|---|---|
| Age + GSCR + Diameter + Sample Size | 0.00 | 0.70 | 0.73 | |
| Diameter + Canopy Cover + Sample Size | 2.41 | 0.21 | 0.67 | * |
| Age + GSCR + Diameter + Predators + Canopy Cover + Sample Size | 4.57 | 0.07 | 0.75 | |
| GSCR + Predators + Sample Size | 8.69 | 0.01 | 0.59 | |
| Age + GSCR + Diameter | 9.90 | 0.00 | 0.57 | * |
| Sample Size | 12.13 | 0.00 | 0.44 | * |
| Diameter + Canopy Cover | 14.21 | 0.00 | 0.45 | * |
| GSCR + Predators | 18.10 | 0.00 | 0.37 | * |
| Predators | 21.21 | 0.00 | 0.23 | * |
| Diameter | 22.10 | 0.00 | 0.21 | * |
| GSCR | 23.25 | 0.00 | 0.17 | * |
| Age | 23.41 | 0.00 | 0.17 | * |
| ~1 | 26.28 | 0.00 | 0.00 |
| Coefficients | Estimate | Std Error | 95% Confidence Interval | t | p | |
|---|---|---|---|---|---|---|
| Intercept | −6.46 | 3.39 | −13.10 | 0.18 | −1.91 | 0.07 |
| Age | 2.10 | 0.63 | 0.87 | 3.32 | 3.36 | 0.003 |
| GSCR | 11.46 | 6.05 | −0.40 | 23.32 | 1.89 | 0.07 |
| Diameter | 0.012 | 0.003 | 0.006 | 0.018 | 3.85 | 0.001 |
| Sample Size | 0.26 | 0.07 | 0.12 | 0.40 | 3.704 | 0.001 |
| Predictor | Min | Max | Mean ± SE |
|---|---|---|---|
| Age | 1.5 | 5 | 3.12 ± 0.21 |
| Canopy Cover (0–1) | 0.00 | 0.52 | 0.06 ± 0.03 |
| Diameter (m) | 3.16 | 11.94 | 6.77 ± 0.40 |
| GSCR (0–1) | 0.01 | 0.44 | 0.15 ± 0.02 |
| Pool Cover (0–1) | 0 | 0.53 | 0.23 ± 0.02 |
| Predators (0–3) | 0.00 | 3.00 | 1.72 ± 0.13 |
| Sample Size per Pool | 10.00 | 44.00 | 26.17 ± 2.00 |
| Model | ΔAICc | wi | Deviance Explained | All Predictors Significant p < 0.05 |
|---|---|---|---|---|
| Pool Cover + Sample Size | 0.00 | 0.27 | 0.32 | |
| GSCR + Sample Size | 0.89 | 0.17 | 0.30 | |
| Sample Size | 1.11 | 0.16 | 0.23 | * |
| Pool Cover + Sample Size + Region | 1.69 | 0.12 | 0.34 | |
| Pool Cover + Sample Size + Diameter | 1.92 | 0.1 | 0.34 | |
| Pool Cover + Sample Size + Predators | 2.46 | 0.08 | 0.33 | |
| GSCR | 3.20 | 0.05 | 0.17 | * |
| Pool Cover | 4.76 | 0.03 | 0.12 | |
| ~1 | 6.27 | 0.01 | 0.00 | |
| Pool Cover + Region + Diameter + Predators | 6.62 | 0.01 | 0.30 |
| Coefficients | Estimate | Std Error | 95% Confidence Interval | t | p | |
|---|---|---|---|---|---|---|
| Intercept | 0.13 | 0.02 | 0.08 | 0.17 | 5.66 | <0.0001 |
| Pool Cover | −0.10 | 0.05 | −0.21 | 0.003 | −1.91 | 0.07 |
| Sample Size | −0.002 | 0.0006 | −0.003 | −0.0005 | −2.76 | 0.01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Millikin, A.R.; Coster, S.S.; Welsh, A.B.; Anderson, J.T. Pool Age and Local Habitat Are Associated with Effective Number of Breeders in Spotted Salamander Populations Colonizing Created Vernal Pools. Diversity 2023, 15, 124. https://doi.org/10.3390/d15020124
Millikin AR, Coster SS, Welsh AB, Anderson JT. Pool Age and Local Habitat Are Associated with Effective Number of Breeders in Spotted Salamander Populations Colonizing Created Vernal Pools. Diversity. 2023; 15(2):124. https://doi.org/10.3390/d15020124
Chicago/Turabian StyleMillikin, Alice R., Stephanie S. Coster, Amy B. Welsh, and James T. Anderson. 2023. "Pool Age and Local Habitat Are Associated with Effective Number of Breeders in Spotted Salamander Populations Colonizing Created Vernal Pools" Diversity 15, no. 2: 124. https://doi.org/10.3390/d15020124
APA StyleMillikin, A. R., Coster, S. S., Welsh, A. B., & Anderson, J. T. (2023). Pool Age and Local Habitat Are Associated with Effective Number of Breeders in Spotted Salamander Populations Colonizing Created Vernal Pools. Diversity, 15(2), 124. https://doi.org/10.3390/d15020124