
Impact of Nesting Mode, Diet, and Taxonomy in Structuring the Associated Microbial Communities of Amazonian Ants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extractions
2.3. DNA Amplification
2.4. Bacterial Quantification
2.5. Bacterial and Microbial Eukaryote Diversity
2.6. Statistical Analysis
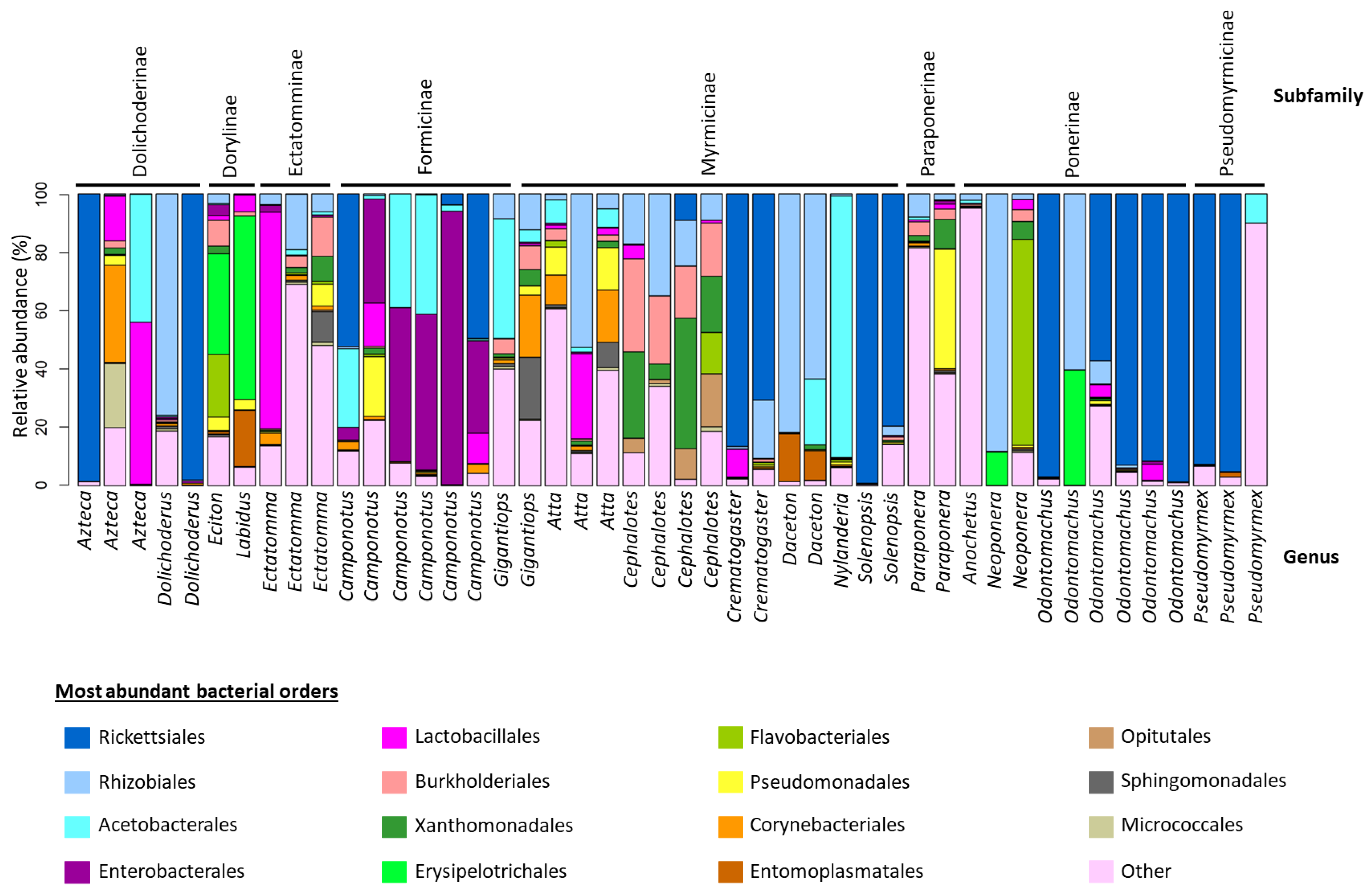
3. Results
3.1. 16S rRNA Assessing Sequencing Quality
3.2. 16S rRNA Alpha Diversity
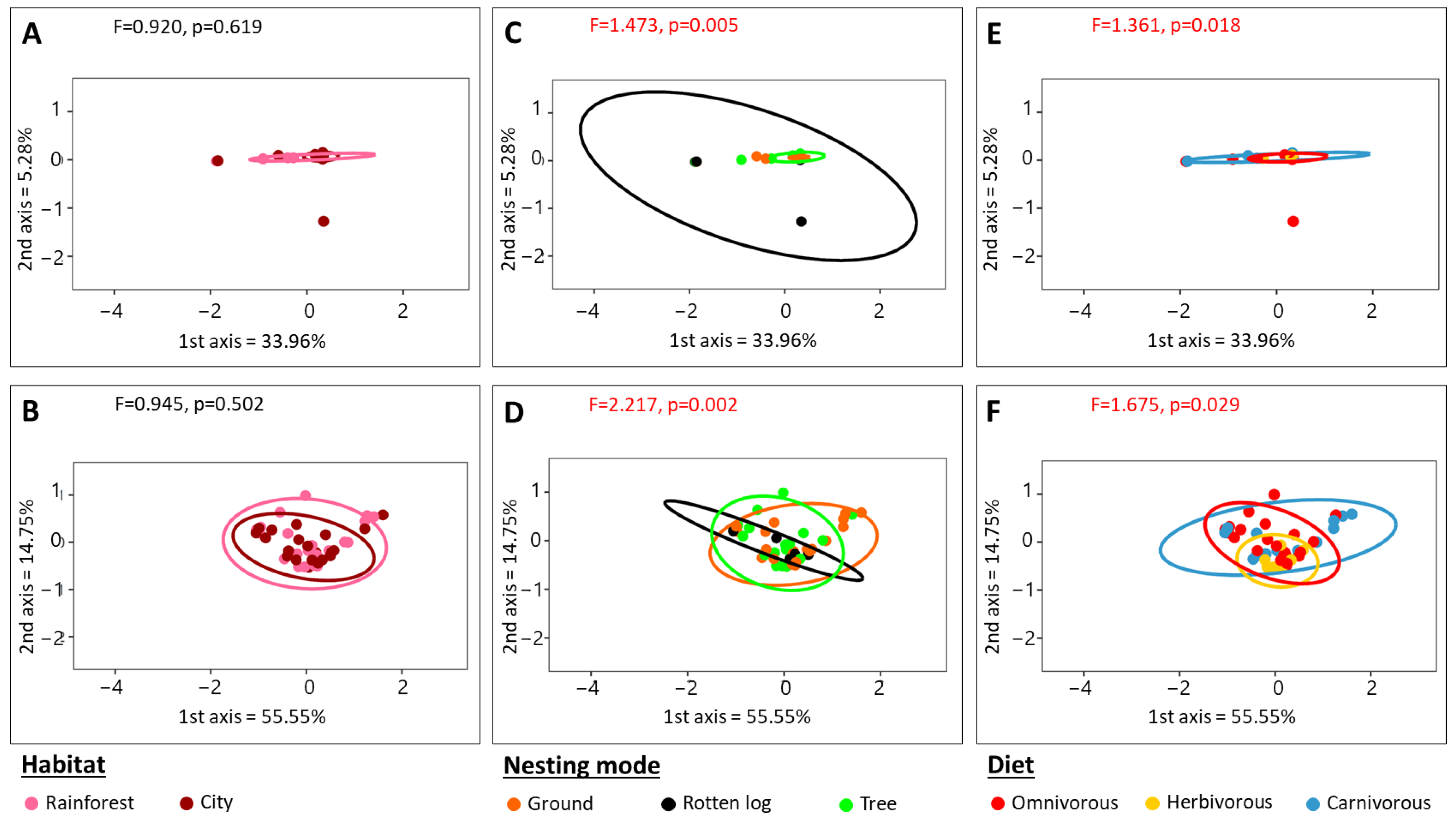
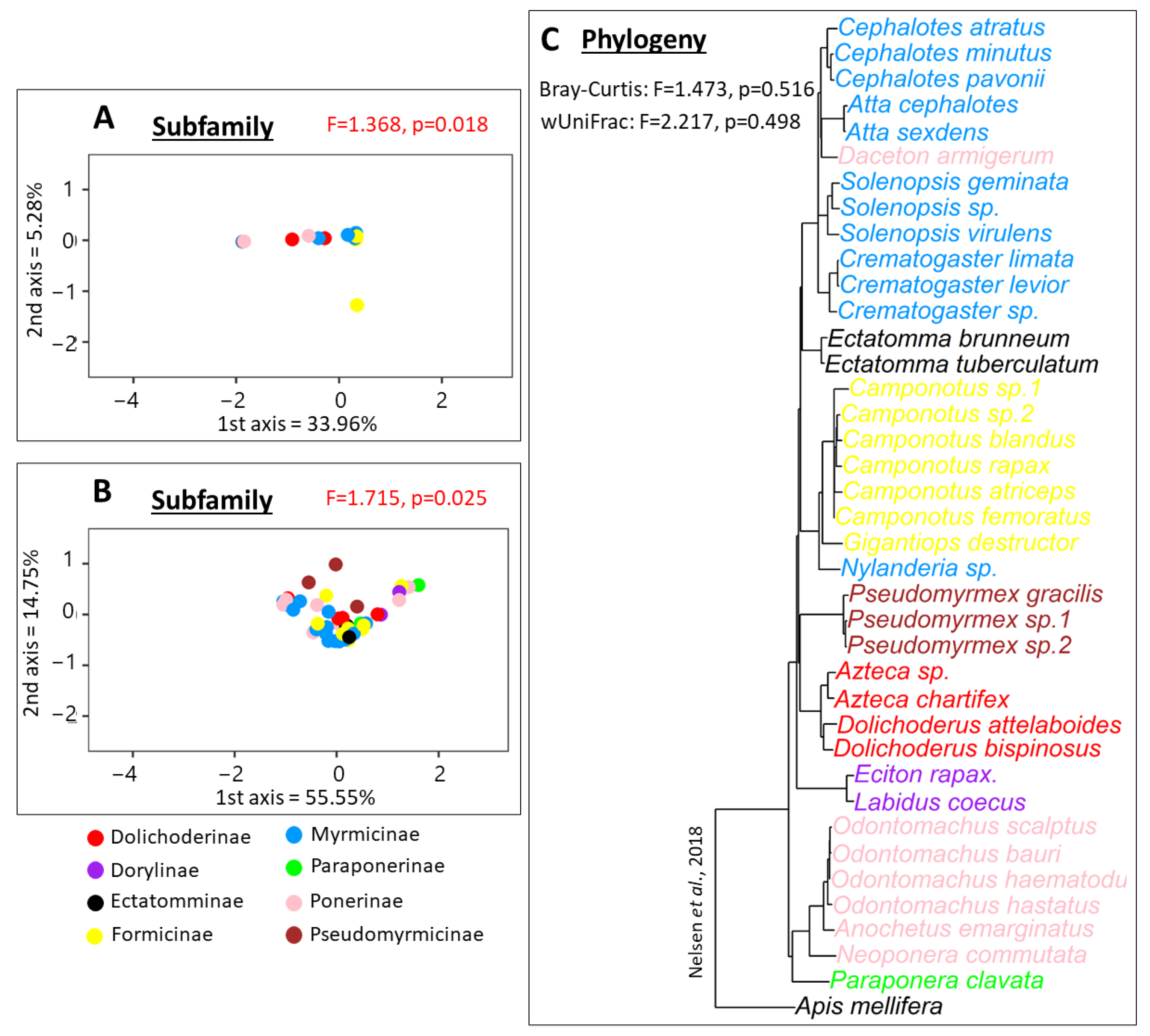
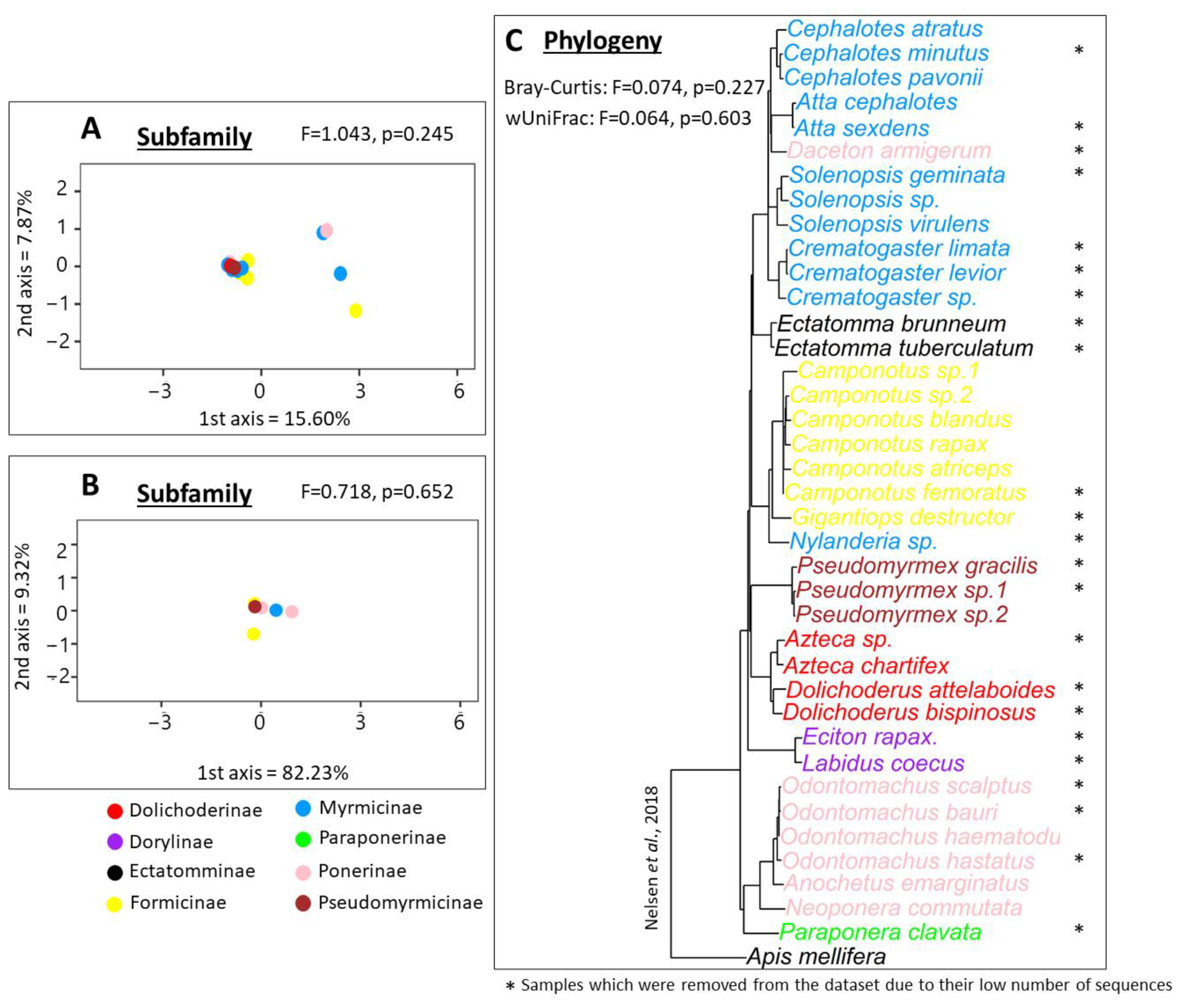
3.3. 16S rRNA Beta Diversity Analysis
3.4. 16S rRNA ASV Similarity Percentage Analysis
3.5. 18S rRNA Assessing Sequencing Quality
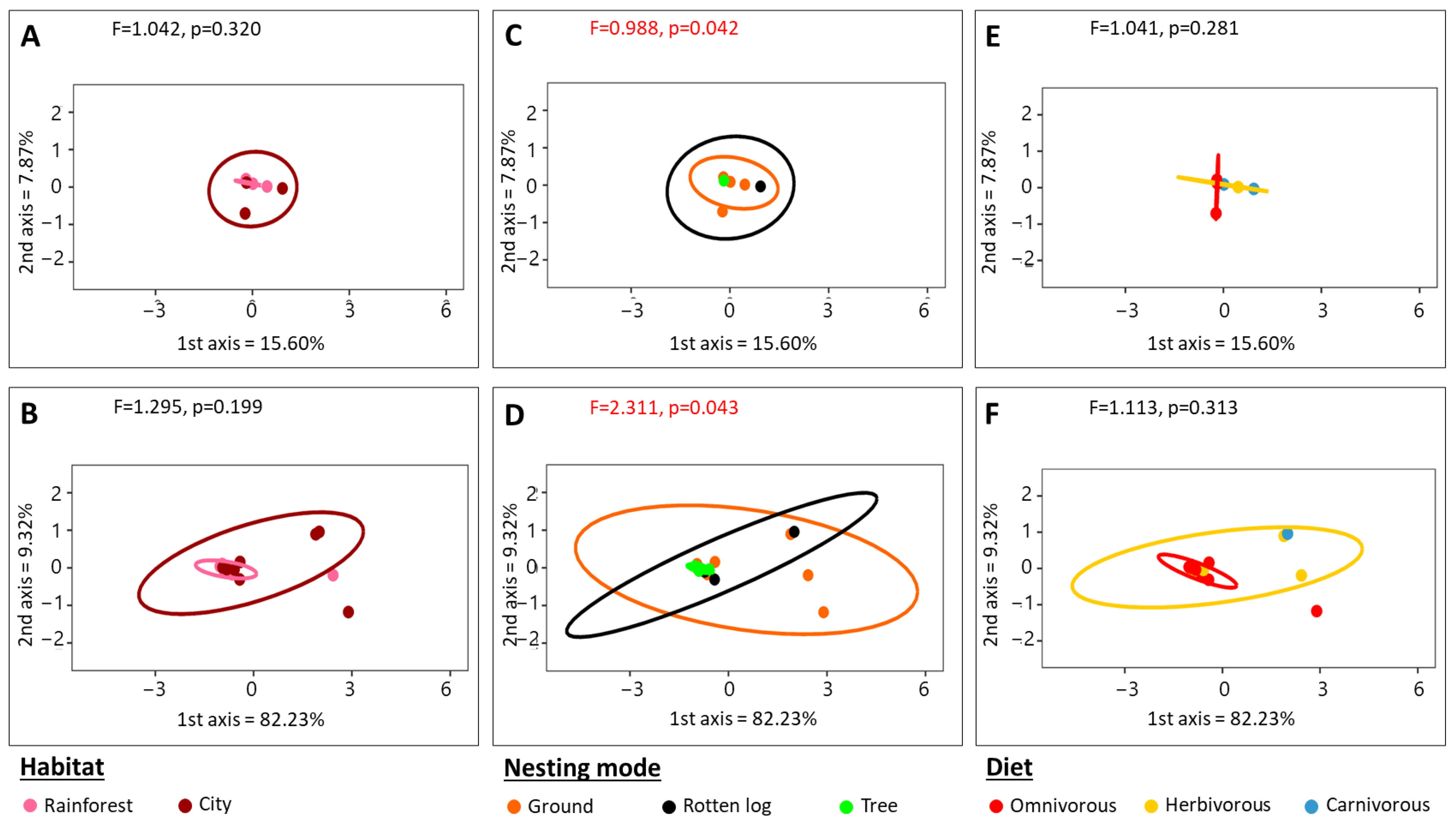
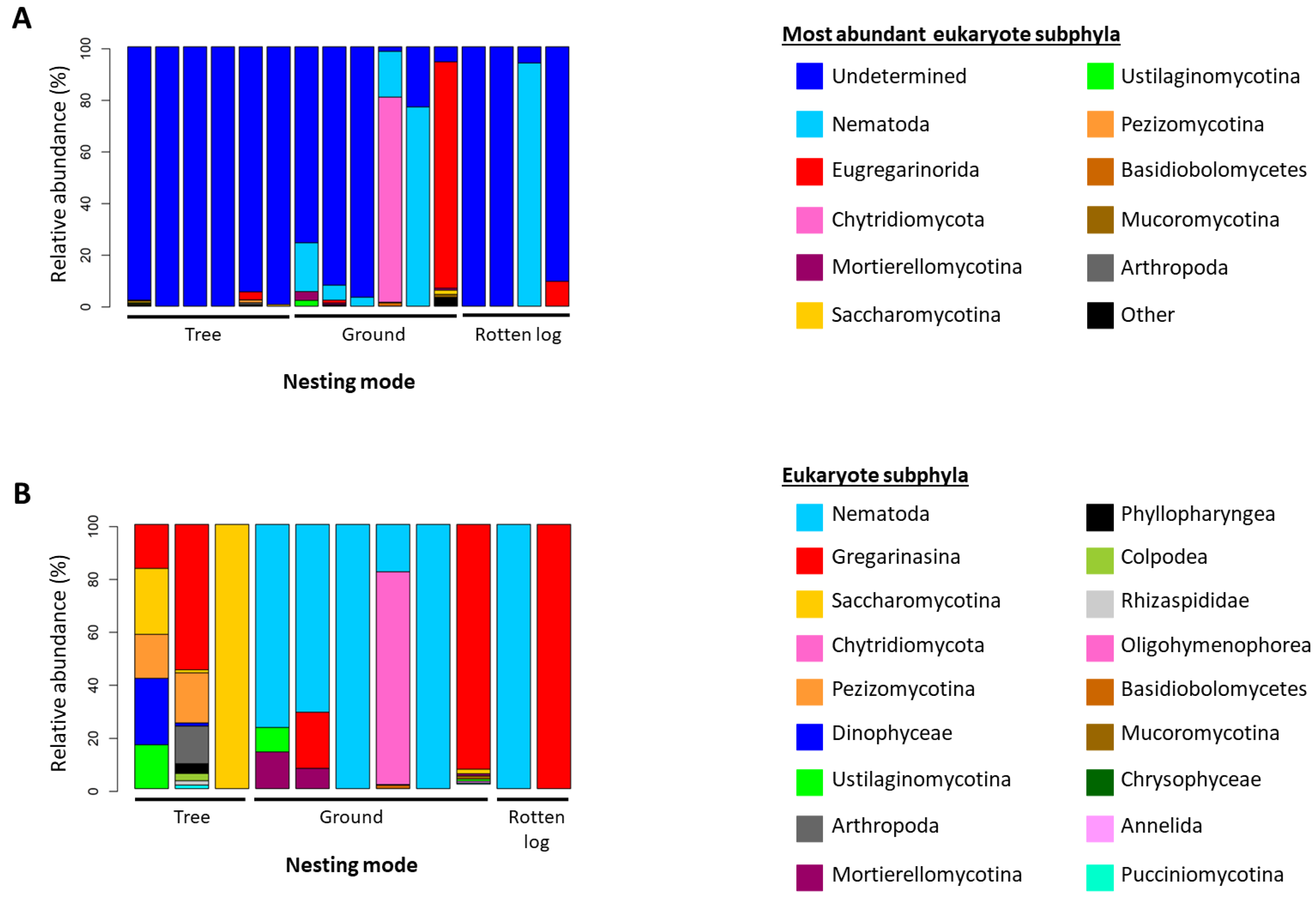
3.6. 18S rRNA Alpha and Beta Diversity Analysis
3.7. 18S rRNA ASV Similarity Percentage Analysis
4. Discussion
4.1. Dietary Niche Structures Bacterial Communities but Not Microbial Eukaryote Communities Associated with Amazonian Ants
4.2. Microbial Community Structure Associated with Amazonian Ants Is Influenced by Abiotic Factors and Nesting Modes
4.3. Bacterial Communities in Long-Term Association with Specific Ant Hosts Are Conserved
4.4. Habitat Does Not Participate in Structuring Microbial Communities Associated with Amazonian Ants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoffmeister, T.S.; Vet, L.E.M.; Biere, A.; Holsinger, K.; Filser, J. Ecological and Evolutionary Consequences of Biological Invasion and Habitat Fragmentation. Ecosystems 2005, 8, 657–667. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Community Diversity: Relative Roles of Local and Regional Processes. Science 1987, 235, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Maire, V.; Gross, N.; Börger, L.; Proulx, R.; Wirth, C.; Pontes, L.d.S.; Soussana, J.F.; Louault, F. Habitat Filtering and Niche Differentiation Jointly Explain Species Relative Abundance within Grassland Communities along Fertility and Disturbance Gradients. New Phytol. 2012, 196, 497–509. [Google Scholar] [CrossRef] [PubMed]
- Arthur, W. The Niche in Competition and Evolution; Wiley-Blackwell: New York, NY, USA, 1987. [Google Scholar]
- Hickman, J.C. Energy Allocation and Niche Differentiation in Four Co-Existing Annual Species of Polygonum in Western North America. J. Ecol. 1977, 65, 317–326. [Google Scholar] [CrossRef]
- Konopka, A. What Is Microbial Community Ecology. ISME J. 2009, 3, 1223–1230. [Google Scholar] [CrossRef] [Green Version]
- Allison, S.D.; Martiny, J.B.H. Resistance, Resilience, and Redundancy in Microbial Communities. Proc. Natl. Acad. Sci. USA 2008, 105, 11512–11519. [Google Scholar] [CrossRef] [Green Version]
- Hollister, E.B.; Engledow, A.S.; Hammett, A.J.M.; Provin, T.L.; Wilkinson, H.H.; Gentry, T.J. Shifts in Microbial Community Structure along an Ecological Gradient of Hypersaline Soils and Sediments. ISME J. 2010, 4, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and Function of the Global Ocean Microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [Green Version]
- Stres, B.; Danevčič, T.; Pal, L.; Fuka, M.M.; Resman, L.; Leskovec, S.; Hacin, J.; Stopar, D.; Mahne, I.; Mandic-Mulec, I. Influence of Temperature and Soil Water Content on Bacterial, Archaeal and Denitrifying Microbial Communities in Drained Fen Grassland Soil Microcosms. FEMS Microbiol. Ecol. 2008, 66, 110–122. [Google Scholar] [CrossRef] [Green Version]
- Nicol, G.W.; Leininger, S.; Schleper, C.; Prosser, J.I. The Influence of Soil PH on the Diversity, Abundance and Transcriptional Activity of Ammonia Oxidizing Archaea and Bacteria. Environ. Microbiol. 2008, 10, 2966–2978. [Google Scholar] [CrossRef]
- Kuffner, M.; Hai, B.; Rattei, T.; Melodelima, C.; Schloter, M.; Zechmeister-Boltenstern, S.; Jandl, R.; Schindlbacher, A.; Sessitsch, A. Effects of Season and Experimental Warming on the Bacterial Community in a Temperate Mountain Forest Soil Assessed by 16S RRNA Gene Pyrosequencing. FEMS Microbiol. Ecol. 2012, 82, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil PH Drives the Spatial Distribution of Bacterial Communities along Elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict Host-Symbiont Cospeciation and Reductive Genome Evolution in Insect Gut Bacteria. PLoS Biol. 2006, 4, 1841–1851. [Google Scholar] [CrossRef]
- Jiggins, F.M.; Hurst, G.D.D. Rapid Insect Evolution by Symbiont Transfer. Science 2011, 332, 185–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnecke, F.; Luginbühl, P.; Ivanova, N.; Ghassemian, M.; Richardson, T.H.; Stege, J.T.; Cayouette, M.; McHardy, A.C.; Djordjevic, G.; Aboushadi, N.; et al. Metagenomic and Functional Analysis of Hindgut Microbiota of a Wood-Feeding Higher Termite. Nature 2007, 450, 560–565. [Google Scholar] [CrossRef] [Green Version]
- Duron, O.; Morel, O.; Noël, V.; Buysse, M.; Binetruy, F.; Lancelot, R.; Loire, E.; Ménard, C.; Bouchez, O.; Vavre, F.; et al. Tick-Bacteria Mutualism Depends on B Vitamin Synthesis Pathways. Curr. Biol. 2018, 28, 1896–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akman Gündüz, E.; Douglas, A.E. Symbiotic Bacteria Enable Insect to Use a Nutritionally Inadequate Diet. Proc. R. Soc. B Biol. Sci. 2012, 276, 987–991. [Google Scholar] [CrossRef] [Green Version]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional Upgrading for Omnivorous Carpenter Ants by the Endosymbiont Blochmannia. BMC Biol. 2007, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Review Lessons from Studying Insect Symbioses. CHOM 2011, 10, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Block, W.; Erzinclioglu, Y.Z.; Worland, M.R. Cold Resistance in All Life Stages of Two Blowfly Species (Diptera, Calliphoridae). Med. Vet. Entomol. 1990, 4, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Hückesfeld, S.; Niederegger, S.; Schlegel, P.; Heinzel, H.-G.; Spiess, R. Feel the Heat: The Effect of Temperature on Development, Behavior and Central Pattern Generation in 3rd Instar Calliphora Vicina Larvae. J. Insect Physiol. 2011, 57, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Peccia, J.; Werth, H.M.; Shelly, M.; Mark, H. Effects of Relative Humidity on the Ultraviolet Induced Inactivation of Airborne Bacteria. Aerosol Sci. Technol. 2001, 35, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Engl, T.; Eberl, N.; Gorse, C.; Krüger, T.; Schmidt, T.H.P.; Plarre, R.; Adler, C.; Kaltenpoth, M. Ancient Symbiosis Confers Desiccation Resistance to Stored Grain Pest Beetles. Mol. Ecol. 2018, 27, 2095–2108. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; Henson, J.; Van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.-O.; Redman, R.S. Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2008, 2, 404–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. The Diversity of Reproductive Parasites among Arthropods: Wolbachia Do Not Walk Alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardo, T.E.; Mondor, E.B. Symbiont Modifies Host Life-History Traits That Affect Gene Flow. Proc. R. Soc. B Biol. Sci. 2006, 273, 1079–1084. [Google Scholar] [CrossRef] [Green Version]
- Goodacre, S.L.; Martin, O.Y.; Bonte, D.; Hutchings, L.; Woolley, C.; Ibrahim, K.; Thomas, C.F.G.; Hewitt, G.M. Microbial Modification of Host Long-Distance Dispersal Capacity. BMC Biol. 2009, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.C. Co-Evolution of Entomogenous Fungi and Their Insect Hosts. In Co-Evolution of Fungi with Plants and Animals; Academic Press: London, UK, 1988. [Google Scholar]
- Karling, J.S. Chytridiosis of Scale Insects. Am. J. Bot. 1948, 35, 246–254. [Google Scholar] [CrossRef]
- Evans, H.C. Mycopathogens of Insects of Epigeal and Aerial Habitats. In Insect-Fungus Interactions; Academic Press: London, UK, 1989; pp. 205–238. [Google Scholar]
- Martin, W.W. Two Additional Species of Catenaria (Chytridiomycetes, Blastocladiales) Parasitic in Midge Eggs. Mycologia 1978, 70, 461–467. [Google Scholar] [CrossRef]
- Matsuura, K.; Tanaka, C.; Nishida, T. Symbiosis of a Termite and a Sclerotium-Forming Fungus: Sclerotia Mimic Termite Eggs. Ecol. Res. 2000, 15, 405–414. [Google Scholar] [CrossRef]
- Evans, H.C.; Samson, R.A. Sporodiniella umbellata, an Entomogenous Fungus of the Mucorales from Cocoa Farms in Ecuador. Can. J. Bot. 1977, 55, 2981–2984. [Google Scholar] [CrossRef]
- Horn, B.W.; Lichtwardt, R.W. Studies on the Nutritional Relationship of Larval Aedes aegypti (Diptera: Culicidae) with Smittium culisetae (Trichomycetes). Mycologia 1981, 73, 724–740. [Google Scholar] [CrossRef]
- Roberts, D.W.; Humber, R.A. Entomogenous Fungi. In Biology of Conidial Fungi; Elsevier: Amsterdam, The Netherlands, 1981; pp. 201–236. [Google Scholar]
- Humber, R.A. Entomophthoromycota: A New Phylum and Reclassification for Entomophthoroid Fungi. Mycotaxon 2012, 120, 477–492. [Google Scholar] [CrossRef]
- Armer, C.A.; Rao, S.; Berry, R.E.; Armer, C.A.; Rao, S.; Berry, R.E. Insect Cellular and Chemical Limitations to Pathogen Development: The Colorado Potato Beetle, the Nematode Heterorhabditis marelatus, and Its Symbiotic Bacteria. J. Invertebr. Pathol. 2004, 87, 114–122. [Google Scholar] [CrossRef]
- Taylor, D.B.; Szalanski, A.L.; Adams, B.J.; Peterson, R.D. Susceptibility of House Fly (Diptera: Muscidae) Larvae to Entomopathogenic Nematodes (Rhabditida: Heterorhabditidae, Steinernematidae). Environ. Entomol. 1998, 27, 1514–1519. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Y.; Cowles, R.S.; Cowles, E.A.; Gaugler, R.; Cox-Foster, D.L.; Li, X.Y.; Cowles, R.S.; Cowles, E.A.; Gaugler, R.; Cox-Foster, D.L. Relationship between the Successful Infection by Entomopathogenic Nematodes and the Host Immune Response. Int. J. Parasitol. 2007, 37, 365–374. [Google Scholar] [CrossRef]
- van Sambeek, J.; Wiesner, A.; Van Sambeek, J.; Wiesner, A. Successful Parasitation of Locusts by Entomopathogenic Nematodes Is Correlated with Inhibition of Insect Phagocytes. J. Invertebr. Pathol. 1999, 73, 154–161. [Google Scholar] [CrossRef]
- Zhao, X.; Ferdig, M.T.; Li, J.; Christensen, B.M. Biochemical Pathway of Melanotic Encapsulation of Brugia malayi in the Mosquito Armigeres subalbatus. Dev. Comp. Immunol. 1995, 19, 205–215. [Google Scholar] [CrossRef]
- Aliota, M.T.; Chen, C.-C.; Dagoro, H.; Fuchs, J.F.; Christensen, B.M. Filarial Worms Reduce Plasmodium Infectivity in Mosquitoes. PLoS Negl. Trop. Dis. 2011, 5, e963. [Google Scholar] [CrossRef]
- Vegni Talluri, M.; Cancrini, G. An Ultrastructural Study on the Early Cellular Response to Dirofilaria immitis (Nematoda) in the Malpighian Tubules of Aedes aegypti (Refractory Strains). Parasite 1994, 1, 343–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cupp, M.S.; Chen, Y.; Cupp, E.W. Cellular Hemolymph Response of Simulium Vittatum (Diptera: Simuliidae) to Intrathoracic Injection of Onchocerca lienalis (Filarioidea: Onchocercidae) Microfilariae. J. Med. Entomol. 1997, 34, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Bartholomay, L.C.; Waterhouse, R.M.; Mayhew, G.F.; Campbell, C.L.; Michel, K.; Zou, Z.; Ramirez, J.L.; Das, S.; Alvarez, K.; Arensburger, P.; et al. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science 2010, 330, 88–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, R.; Briggs, J.D. Occurrence and Control of Aphanomyces (Saprolegniales: Fungi) Infections in Laboratory Colonies of Larval Anopheles. J. Am. Mosq. Control Assoc. 1985, 1, 100–102. [Google Scholar]
- Stephen, K.; Ipek Kurtböke, D. Screening of Oomycete Fungi for Their Potential Role in Reducing the Biting Midge (Diptera: Ceratopogonidae) Larval Populations in Hervey Bay, Queensland, Australia. Int. J. Environ. Res. Public Health 2011, 8, 15601574. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, P.H.W.; Vega, F.E. Ecology and Evolution of Insect—Fungus Mutualisms. Annu. Rev. Entomol. 2020, 65, 431–455. [Google Scholar] [CrossRef] [Green Version]
- Schultz, T.R.; Brady, S.G. Major Evolutionary Transitions in Ant Agriculture. Proc. Natl. Acad. Sci. USA 2008, 105, 5435–5440. [Google Scholar] [CrossRef] [Green Version]
- Hulcr, J.; Stelinski, L.L. The Ambrosia Symbiosis: From Evolutionary Ecology to Practical Management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [Green Version]
- Aanen, D.K.; De Fine Licht, H.H.; Debets, A.J.M.; Kerstes, N.A.G.; Hoekstra, R.F.; Boomsma, J.J. High Symbiont Relatedness Stabilizes Mutualistic Cooperation in Fungusgrowing Termites. Science 2009, 326, 1103–1106. [Google Scholar] [CrossRef] [Green Version]
- Defossez, E.; Selosse, M.A.; Dubois, M.P.; Mondolot, L.; Faccio, A.; Djieto-Lordon, C.; McKey, D.; Blatrix, R. Ant-Plants and Fungi: A New Threeway Symbiosis. New Phytol. 2009, 182, 942–949. [Google Scholar] [CrossRef]
- Ruiz-González, M.X.; Malé, P.J.G.; Leroy, C.; Dejean, A.; Gryta, H.; Jargeat, P.; Quilichini, A.; Orivel, J. Specific, Non-Nutritional Association between an Ascomycete Fungus and Allomerus Plant-Ants. Biol. Lett. 2011, 7, 475–479. [Google Scholar] [CrossRef]
- Nepel, M.; Voglmayr, H.; Schönenberger, J.; Mayer, V.E. High Diversity and Low Specificity of Chaetothyrialean Fungi in Carton Galleries in a Neotropical Ant-Plant Association. PLoS ONE 2014, 9, e112756. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Guo, W.; Ding, J. Mycangial Fungus Benefits the Development of a Leaf-Rolling Weevil, Euops chinesis. J. Insect Physiol. 2012, 58, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Kobune, S.; Kajimura, H.; Masuya, H.; Kubono, T. Symbiotic Fungal Flora in Leaf Galls Induced by Illiciomyia yukawai (Diptera: Cecidomyiidae) and in its Mycangia. Microb. Ecol. 2012, 63, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Stefanini, I. Yeast-Insect Associations: It Takes Guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Florez, L.V.; Biedermann, P.H.W.; Engl, T.; Kaltenpoth, M. Defensive Symbioses of Animals with Prokaryotic and Eukaryotic Microorganisms. R. Soc. Chem. 2015, 32, 904–936. [Google Scholar] [CrossRef] [Green Version]
- Goodrich-Blair, H.; Clarke, D.J. Mutualism and Pathogenesis in Xenorhabdus and Photorhabdus: Two Roads to the Same Destination. Mol. Microbiol. 2007, 64, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Miranda, V.A.; Navarro, P.D.; Davidowitz, G.; Bronstein, J.; Stock, S.P. Effect of Insect Host Age and Diet on the Fitness of the Entomopathogenic Nematode-Bacteria Mutualism. Symbiosis 2013, 61, 145–153. [Google Scholar] [CrossRef]
- Griffiths, H.M.; Ashton, L.A.; Walker, A.E.; Hasan, F.; Evans, T.A.; Eggleton, P.; Parr, C.L. Ants Are the Major Agents of Resource Removal from Tropical Rainforests. J. Anim. Ecol. 2018, 87, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Davidson, D.W.; Cook, S.C.; Snelling, R.R.; Chua, T.H. Explaining the Abundance of Ants in Lowland Tropical Rainforest Canopies. Science 2003, 300, 969–972. [Google Scholar] [CrossRef] [Green Version]
- Cook, S.C.; Davidson, D.W. Nutritional and Functional Biology of Exudate-Feeding Ants. Entomol. Exp. Appl. 2006, 118, 1–10. [Google Scholar] [CrossRef]
- Anderson, K.E.; Russell, J.A.; Moreau, C.S.; Kautz, S.; Sullam, K.E.; Hu, Y.; Basinger, U.; Mott, B.M.; Buck, N.; Wheeler, D.E. Highly Similar Microbial Communities Are Shared among Related and Trophically Similar Ant Species. Mol. Ecol. 2012, 21, 2282–2296. [Google Scholar] [CrossRef]
- Funaro, C.F.; Kronauer, D.J.C.; Moreau, C.S.; Goldman-Huertas, B.; Pierce, N.E.; Russell, J.A. Army Ants Harbor a Host-Specific Clade of Entomoplasmatales Bacteria. Appl. Environ. Microbiol. 2011, 77, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Łukasik, P.; Newton, J.A.; Sanders, J.G.; Hu, Y.; Moreau, C.S.; Kronauer, D.J.C.; O’Donnell, S.; Koga, R.; Russell, J.A. The Structured Diversity of Specialized Gut Symbionts of the New World Army Ants. Mol. Ecol. 2017, 26, 3808–3825. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.G.; Powell, S.; Kronauer, D.J.C.; Vasconcelos, H.L.; Frederickson, M.E.; Pierce, N.E. Stability and Phylogenetic Correlation in Gut Microbiota: Lessons from Ants and Apes. Mol. Ecol. 2014, 23, 1268–1283. [Google Scholar] [CrossRef]
- Hu, Y.; Sanders, J.G.; Łukasik, P.; D’Amelio, C.L.; Millar, J.S.; Vann, D.R.; Lan, Y.; Newton, J.A.; Schotanus, M.; Kronauer, D.J.C.; et al. Herbivorous Turtle Ants Obtain Essential Nutrients from a Conserved Nitrogen-Recycling Gut Microbiome. Nat. Commun. 2018, 9, 964. [Google Scholar] [CrossRef] [Green Version]
- Wernegreen, J.J.; Kauppinen, S.N.; Brady, S.G.; Ward, P.S. One Nutritional Symbiosis Begat Another: Phylogenetic Evidence That the Ant Tribe Camponotini Acquired Blochmannia by Tending Sap-Feeding Insects. BMC Evol. Biol. 2009, 9, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho, M.O.; Duplais, C.; Orivel, J.; Dejean, A.; Gibson, J.C.; Suarez, A.V.; Moreau, C.S. Development but Not Diet Alters Microbial Communities in the Neotropical Arboreal Trap Jaw Ant Daceton armigerum: An Exploratory Study. Sci. Rep. 2020, 10, 7350. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.S.; Rubin, B.E.R. Diversity and Persistence of the Gut Microbiome of the Giant Neotropical Bullet Ant. Integr. Comp. Biol. 2017, 57, 682–689. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, T.B.; Ferro, M.; Bacci, M.; De Souza, D.J.; Fontana, R.; Delabie, J.H.C.; Silva, A. Bacterial Communities in the Midgut of Ponerine Ants (Hymenoptera: Formicidae: Ponerinae). Sociobiology 2016, 63, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Rubin, B.E.R.; Kautz, S.; Wray, B.D.; Moreau, C.S. Dietary Specialization in Mutualistic Acacia-Ants Affects Relative Abundance but Not Identity of Host-Associated Bacteria. Mol. Ecol. 2019, 28, 900–916. [Google Scholar] [CrossRef]
- Martins, C.; Souza, R.F.; Bueno, O.C. Presence and Distribution of the Endosymbiont Wolbachia among Solenopsis spp. (Hymenoptera: Formicidae) from Brazil and its Evolutionary History. J. Invertebr. Pathol. 2012, 109, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramalho, M.O.; Bueno, O.C.; Moreau, C.S. Microbial Composition of Spiny Ants (Hymenoptera: Formicidae: Polyrhachis) across Their Geographic Range. BMC Evol. Biol. 2017, 17, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Snyder, L.; Meador, C.A.D.; Naldo, R.; Mott, B.; Anderson, K.E. Parasaccharibacter apium, Gen. Nov., Sp. Nov., Improves Honey Bee (Hymenoptera: Apidae) Resistance to Nosema. J. Econ. Entomol. 2016, 109, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Mitraka, E.; Stathopoulos, S.; Siden-Kiamos, I.; Christophides, G.K.; Louis, C. Asaia Accelerates Larval Development of Anopheles gambiae. Pathog. Glob. Health 2013, 107, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapountzis, P.; Zhukova, M.; Hansen, L.H.; Sørensen, S.J.; Schiøtt, M.; Boomsma, J.J. Acromyrmex Leaf-Cutting Ants Have Simple Gut Microbiota with Nitrogen-Fixing Potential. Appl. Environ. Microbiol. 2015, 81, 5527–5537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dussutour, A.; Simpson, S.J. Ant Workers Die Young and Colonies Collapse When Fed a High-Protein Diet. Proc. R. Soc. B Biol. Sci. 2012, 279, 2402–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuvonen, M.M.; Tamarit, D.; Näslund, K.; Liebig, J.; Feldhaar, H.; Moran, N.A.; Guy, L.; Andersson, S.G.E. The Genome of Rhizobiales Bacteria in Predatory Ants Reveals Urease Gene Functions but No Genes for Nitrogen Fixation. Sci. Rep. 2016, 6, 39197. [Google Scholar] [CrossRef] [Green Version]
- Wenseleers, T.; Billen, J. No Evidence for Wolbachia-Induced Parthenogenesis in the Social Hymenoptera. J. Evol. Biol. 2000, 13, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Wolbachia as a Bacteriocyte-Associated Nutritional Mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.V.; Dillon, R.J.; Dillon, V.M.; Reynolds, S.E.; Samuels, R.I. Ocurrence of the Antibiotic Producing Bacterium Burkholderia sp. in Colonies of the Leaf-Cutting Ant Atta sexdens rubropilosa. FEMS Microbiol. Lett. 2004, 239, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.P.; Andersen, S.B.; Hywel-Jones, N.L.; Himaman, W.; Billen, J.; Boomsma, J.J. Behavioral Mechanisms and Morphological Symptoms of Zombie Ants Dying from Fungal Infection. BMC Ecol. 2011, 11, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poinar, G. Nematode Parasites and Associates of Ants: Past and Present. Psyche 2012, 2012, 192017. [Google Scholar] [CrossRef] [Green Version]
- Baccaro, F.B.; Feitosa, R.M.; Fernández, F.; Fernandes, I.O.; Izzo, T.J.; Souza, J.D.; Solar, R. Guia Para Os Gêneros de Formigas Do Brasil, 1st ed.; Editora INPA: Manaus, Brazil, 2015. [Google Scholar] [CrossRef]
- Rubin, B.E.R.; Sanders, J.G.; Hampton-Marcell, J.; Owens, S.M.; Gilbert, J.A.; Moreau, C.S. DNA Extraction Protocols Cause Differences in 16S RRNA Amplicon Sequencing Efficiency but Not in Community Profile Composition or Structure. Microbiologyopen 2014, 3, 910–921. [Google Scholar] [CrossRef] [Green Version]
- Moreau, C.S. A Practical Guide to DNA Extraction, PCR, and Gene-Based DNA Sequencing in Insects. Halteres 2014, 5, 32–42. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-High-Throughput Microbial Community Analysis on the Illumina HiSeq and MiSeq Platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Creer, S.; Fonseca, V.G.; Porazinska, D.L.; Giblin-Davis, R.M.; Sung, W.; Power, D.M.; Packer, M.; Carvalho, G.R.; Blaxter, M.L.; Lambshead, P.J.D.; et al. Ultrasequencing of the Meiofaunal Biosphere: Practice, Pitfalls and Promises. Mol. Ecol. 2010, 19, 4–20. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Bokulich, N.; Kaehler, B.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.; Caporaso, G. Optimizing Taxonomic Classification of Marker Gene Amplicon Sequences. PeerJ Prepr. 2018, 6, e3208v2. [Google Scholar] [CrossRef]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaccard, P. Nouvelles Recherches Sur La Distribution Florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Sorenson, T. A Method of Establishing Groups of Equal Amplitude in Plant Sociology Based on Similarity of Species Content. K. Danske Vidensk. Selsk. 1948, 5, 1–34. [Google Scholar]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and Qualitative β Diversity Measures Lead to Different Insights into Factors That Structure Microbial Communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kassambara, A. Ggpubr: “ggplot2” Based Publication Ready Plots. R Packag. Version 0.3.0. 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 6 January 2023).
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package PMCMR. R Package. 2014. Available online: https://CRAN.R-project.org/package=PMCMR (accessed on 6 January 2023).
- Gómez-Rubio, V. Ggplot2—Elegant Graphics for Data Analysis (2nd Edition). J. Stat. Softw. 2017, 77, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified Interface to Visualize Statistical Results of Popular r Packages. R J. 2016, 8, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. Phytools: An R Package for Phylogenetic Comparative Biology (and Other Things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Ree, R.H.; Moreau, C.S. Ant–Plant Interactions Evolved through Increasing Interdependence. Proc. Natl. Acad. Sci. USA 2018, 115, 12253–12258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muegge, B.D.; Kuczynski, J.; Knights, D.; Clemente, J.C.; González, A.; Fontana, L.; Henrissat, B.; Knight, R.; Gordon, J.I. Diet Drives Convergence in Gut Microbiome Functions across Mammalian Phylogeny and within Humans. Science 2011, 332, 970–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.A.; Moreau, C.S.; Goldman-Huertas, B.; Fujiwara, M.; Lohman, D.J.; Pierce, N.E. Bacterial Gut Symbionts Are Tightly Linked with the Evolution of Herbivory in Ants. Proc. Natl. Acad. Sci. USA 2009, 106, 21236–21241. [Google Scholar] [CrossRef] [PubMed]
- Freter, R.; Brickner, H.; Fekete, J.; Vickerman, M.M.; Carey, K.E. Survival and Implantation of Escherichia coli in the Intestinal Tract. Infect. Immun. 1983, 39, 686–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freter, R.; Brickner, H.; Botney, M.; Cleven, D.; Aranki, A. Mechanisms That Control Bacterial Populations in Continuous-Flow Culture Models of Mouse Large Intestinal Flora. Infect. Immun. 1983, 39, 676–685. [Google Scholar] [CrossRef] [Green Version]
- Papadimitriou, K.; Pot, B.; Tsakalidou, E. How Microbes Adapt to a Diversity of Food Niches. Curr. Opin. Food Sci. 2015, 2, 29–35. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in Animal Nutrition and Health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Steed, H.; Macfarlane, S. Bacterial Metabolism and Health-Related Effects of Galacto-Oligosaccharides and Other Prebiotics. J. Appl. Microbiol. 2008, 104, 305–344. [Google Scholar] [CrossRef]
- Ramirez-Farias, C.; Slezak, K.; Fuller, Z.; Duncan, A.; Holtrop, G.; Louis, P. Effect of Inulin on the Human Gut Microbiota: Stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br. J. Nutr. 2009, 101, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Ivarsson, E.; Roos, S.; Liu, H.Y.; Lindberg, J.E. Fermentable Non-Starch Polysaccharides Increases the Abundance of Bacteroides-Prevotella-Porphyromonas in Ileal Microbial Community of Growing Pigs. Animal 2014, 8, 1777–1787. [Google Scholar] [CrossRef]
- Chung, W.S.F.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the Human Gut Microbiota by Dietary Fibres Occurs at the Species Level. BMC Biol. 2016, 14, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigosky, S.R. Tropical Microclimatic Considerations. In Forest Canopies; Lowman, M.D., Rinker, H.B., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2004; pp. 24–48. [Google Scholar]
- Kumagai, T.; Kuraji, K.; Noguchi, H.; Tanaka, Y.; Tanaka, K.; Suzuki, M. Vertical Profiles of Environmental Factors within Tropical Rainforest, Lambir Hills National Park, Sarawak, Malaysia. J. For. Res. 2001, 6, 257–264. [Google Scholar] [CrossRef]
- Basset, Y.; Hammond, P.M.; Barrios, H.; Holloway, J.D.; Miller, S.E. Vertical Stratification of Arthropod Assemblages. In Arthropods of Tropical Forests: Spatio-Temporal Dynamics and Resource Use in the Canopy; Basset, Y., Novotny, V., Miller, S.E., Kitching, R.L., Eds.; Cambridge University Press: New York, NY, USA, 2003; pp. 17–27. [Google Scholar]
- Gruntenko, N.E.; Ilinsky, Y.Y.; Adonyeva, N.V.; Burdina, E.V.; Bykov, R.A.; Menshanov, P.N.; Rauschenbach, I.Y. Various Wolbachia Genotypes Differently Influence Host Drosophila Dopamine Metabolism and Survival under Heat Stress Conditions. BMC Evol. Biol. 2017, 17, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Burke, G.; Fiehn, O.; Moran, N. Effects of Facultative Symbionts and Heat Stress on the Metabolome of Pea Aphids. ISME J. 2010, 4, 242–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, L.V.; Dhakal, P.; Lebenzon, J.E.; Heinrichs, D.E.; Bucking, C.; Sinclair, B.J. Seasonal Shifts in the Insect Gut Microbiome Are Concurrent with Changes in Cold Tolerance and Immunity. Funct. Ecol. 2018, 32, 2357–2368. [Google Scholar] [CrossRef] [Green Version]
- Rio, R.V.M.; Wu, Y.N.; Filardo, G.; Aksoy, S. Dynamics of Multiple Symbiont Density Regulation during Host Development: Tsetse Fly and its Microbial Flora. Proc. R. Soc. B Biol. Sci. 2006, 273, 805–814. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, M.M.; Engl, T.; Kaltenpoth, M. Microbial Symbionts Expanding or Constraining Abiotic Niche Space in Insects. Curr. Opin. Insect Sci. 2020, 39, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Lawley, B.; Ripley, S.; Bridge, P.; Convey, P. Molecular Analysis of Geographic Patterns of Eukaryotic Diversity in Antarctic Soils. Appl. Environ. Microbiol. 2004, 70, 5963–5972. [Google Scholar] [CrossRef] [Green Version]
- Fell, J.W.; Scorzetti, G.; Connell, L.; Craig, S. Biodiversity of Micro-Eukaryotes in Antarctic Dry Valley Soils with <5% Soil Moisture. Soil Biol. Biochem. 2006, 38, 3107–3119. [Google Scholar] [CrossRef]
- Tian, F.; Yu, Y.; Chen, B.; Li, H.; Yao, Y.F.; Guo, X.K. Bacterial, Archaeal and Eukaryotic Diversity in Arctic Sediment as Revealed by 16S RRNA and 18S RRNA Gene Clone Libraries Analysis. Polar Biol. 2009, 32, 93–103. [Google Scholar] [CrossRef]
- Meadow, J.F.; Zabinski, C.A. Spatial Heterogeneity of Eukaryotic Microbial Communities in an Unstudied Geothermal Diatomaceous Biological Soil Crust: Yellowstone National Park, WY, USA. FEMS Microbiol. Ecol. 2012, 82, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Wolschin, F.; Hölldobler, B.; Gross, R.; Zientz, E. Replication of the Endosymbiotic Bacterium Blochmannia floridanus is Correlated with the Developmental and Reproductive Stages of its Ant Host. Appl. Environ. Microbiol. 2004, 70, 4096–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zientz, E.; Beyaert, I.; Gross, R.; Feldhaar, H. Relevance of the Endosymbiosis of Blochmannia floridanus and Carpenter Ants at Different Stages of the Life Cycle of the Host. Appl. Environ. Microbiol. 2006, 72, 6027–6033. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.P.; Wernegreen, J.J. Deep Divergence and Rapid Evolutionary Rates in Gut-Associated Acetobacteraceae of Ants. BMC Microbiol. 2016, 16, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary Origin of Insect-Wolbachia Nutritional Mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stouthamer, R.; Breeuwer, J.A.J.; Hurst, G.D.D. Wolbachia pipientis: Microbial Manipulator of Arthropod Reproduction. Annu. Rev. Microbiol. 1999, 53, 71–102. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.G.; Lukasik, P.; Frederickson, M.E.; Russell, J.A.; Koga, R.; Knight, R.; Pierce, N.E. Dramatic Differences in Gut Bacterial Densities Correlate with Diet and Habitat in Rainforest Ants. Integr. Comp. Biol. 2017, 57, 705–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.A.; Goldman-Huertas, B.; Moreau, C.S.; Baldo, L.; Stahlhut, J.K.; Werren, J.H.; Pierce, N.E. Specialization and Geographic Isolation among Wolbachia Symbionts from Ants and Lycaenid Butterflies. Evolution 2009, 63, 624–640. [Google Scholar] [CrossRef] [PubMed]
- Greiner, T.; Bäckhed, F. Effects of the Gut Microbiota on Obesity and Glucose Homeostasis. Trends Endocrinol. Metab. 2011, 22, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Hang, I.; Rinttila, T.; Zentek, J.; Kettunen, A.; Alaja, S.; Apajalahti, J.; Harmoinen, J.; de Vos, W.M.; Spillmann, T. Effect of High Contents of Dietary Animal-Derived Protein or Carbohydrates on Canine Faecal Microbiota. BMC Vet. Res. 2012, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnusson, K.R.; Hauck, L.; Jeffrey, B.M.; Elias, V.; Humphrey, A.; Nath, R.; Perrone, A.; Bermudez, L.E. Relationships between Diet-Related Changes in the Gut Microbiome and Cognitive Flexibility. Neuroscience 2015, 300, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Larson, H.K.; Goffredi, S.K.; Parra, E.L.; Vargas, O.; Pinto-Tomas, A.A.; McGlynn, T.P. Distribution and Dietary Regulation of an Associated Facultative Rhizobiales-Related Bacterium in the Omnivorous Giant Tropical Ant, Paraponera clavata. Naturwissenschaften 2014, 101, 397–406. [Google Scholar] [CrossRef]
- Pyle, R.; Bentzien, M.; Opler, P. Insect Conservation. Annu. Rev. Entomol. 1981, 26, 233–258. [Google Scholar] [CrossRef]
- McIntyre, N. Ecology of Urban Arthropods: A Review and a Call to Action. Ann. Entomol. Soc. Am. 2009, 93, 271–276. [Google Scholar] [CrossRef]
- Shaffer, B.; Lighthart, B. Survey of Culturable Airborne Bacteria at Four Diverse Locations in Oregon: Urban, Rural, Forest, and Coastal. Microb. Ecol. 1997, 34, 167–177. [Google Scholar] [CrossRef]
- Belt, K.T.; Hohn, C.; Gbakima, A.; Higgins, J.A. Identification of Culturable Stream Water Bacteria from Urban, Agricultural, and Forested Watersheds Using 16S RRNA Gene Sequencing. J. Water Health 2007, 5, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Reeson, A.F.; Jankovic, T.; Kasper, M.L.; Rogers, S.; Austin, A.D. Application of 16S RDNA-DGGE to Examine the Microbial Ecology Associated with a Social Wasp Vespula germanica. Insect Mol. Biol. 2003, 12, 85–91. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Anaplasma phagocytophilum in Questing Ixodes ricinus Ticks: Comparison of Prevalences and Partial 16S RRNA Gene Variants in Urban, Pasture, and Natural Habitats. Appl. Environ. Microbiol. 2013, 79, 1730–1734. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chanson, A.; Moreau, C.S.; Duplais, C. Impact of Nesting Mode, Diet, and Taxonomy in Structuring the Associated Microbial Communities of Amazonian Ants. Diversity 2023, 15, 126. https://doi.org/10.3390/d15020126
Chanson A, Moreau CS, Duplais C. Impact of Nesting Mode, Diet, and Taxonomy in Structuring the Associated Microbial Communities of Amazonian Ants. Diversity. 2023; 15(2):126. https://doi.org/10.3390/d15020126
Chicago/Turabian StyleChanson, Anaïs, Corrie S. Moreau, and Christophe Duplais. 2023. "Impact of Nesting Mode, Diet, and Taxonomy in Structuring the Associated Microbial Communities of Amazonian Ants" Diversity 15, no. 2: 126. https://doi.org/10.3390/d15020126
APA StyleChanson, A., Moreau, C. S., & Duplais, C. (2023). Impact of Nesting Mode, Diet, and Taxonomy in Structuring the Associated Microbial Communities of Amazonian Ants. Diversity, 15(2), 126. https://doi.org/10.3390/d15020126