1. Introduction
Thalassochelydia corresponds to a very abundant and diverse lineage of turtles for the Upper Jurassic (Oxfordian to Tithonian) record of Europe [
1,
2,
3,
4,
5]. This group was mainly represented by coastal marine forms, with a single known exception: the freshwater taxon
Hylaeochelys Lydekker, 1889 [
6] (see [
7], and references therein). In fact, it has been interpreted that the coastal thalassochelydians disappeared when their habitats, in the European shallow shelf seas, were drastically affected by a major regression at the Jurassic–Cretaceous transition [
4,
7,
8]. Probably thanks to its adaptation to freshwater environments,
Hylaeochelys is the only known thalassochelydian which survived that event, and this genus is recognized in the Cretaceous record (see [
7], and references therein).
The type species of
Hylaeochelys is
Hylaeochelys belli (Mantell, 1844) [
9], a taxon known from the Berriasian to the Valanginian (Early Cretaceous) record of United Kingdom and Germany [
7,
10,
11]. The revision of the genus
Hylaeochelys performed a decade ago allowed the recognition the type species as the only valid representative of the genus (see [
10], and references therein). However, the subsequent study of an articulated partial shell from west-central Portugal allowed the identification of the genus
Hylaeochelys in the Upper Jurassic (Tithonian) record, and the definition of the new species
Hylaeochelys kappa Pérez-García and Ortega, 2014 [
7].
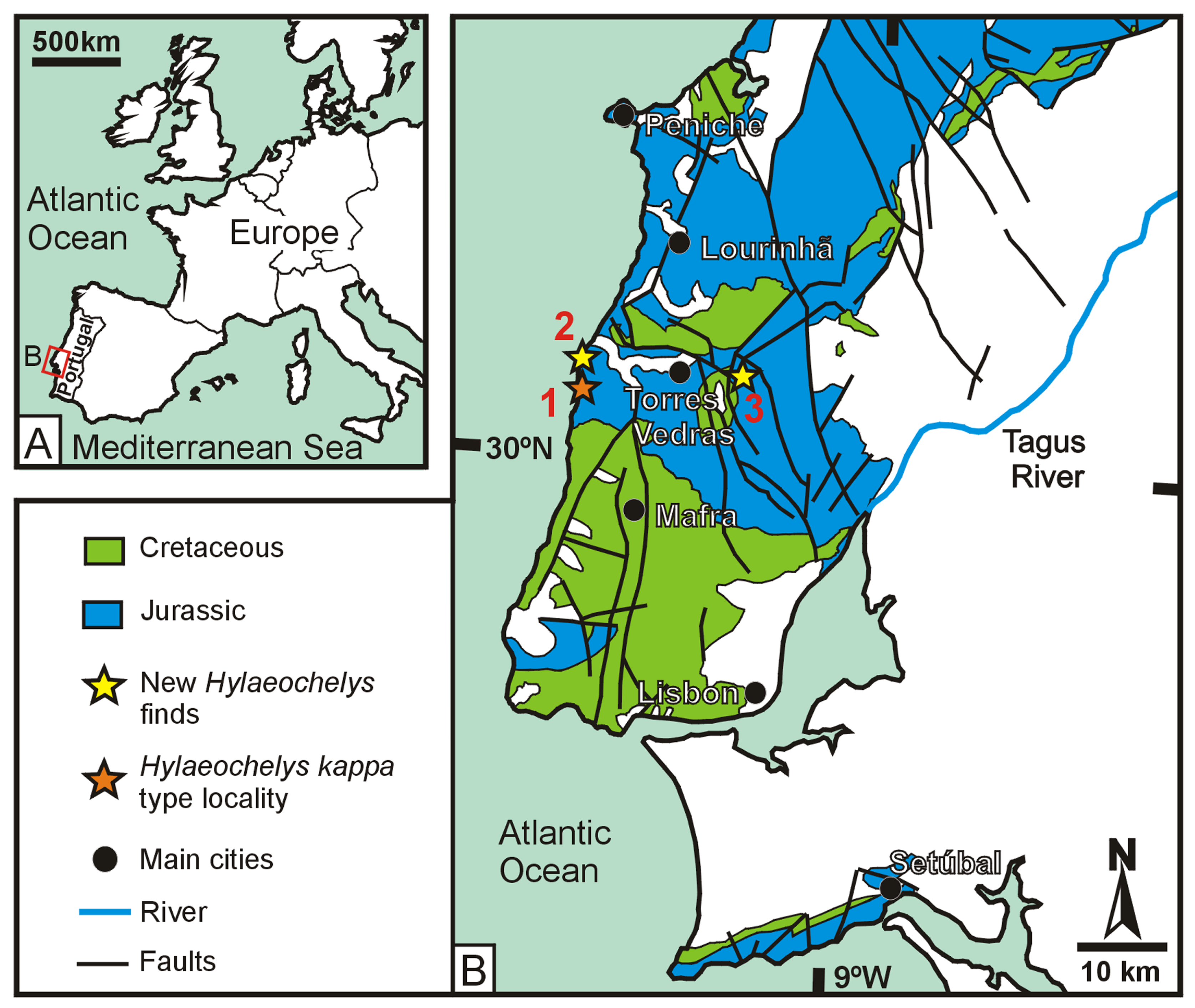
The record of Portuguese Jurassic turtles is relatively abundant and diverse for the European context (see [
4], and references therein), especially considering the finds from the Upper Jurassic (Oxfordian to Tithonian) levels occurred in several areas of the Lusitanian Basin (west-central region of Portugal). Three new turtle taxa were described at these locations. Two of them, belonging to two different clades, are currently identified as valid:
Selenemys lusitanica Pérez-García and Ortega, 2011 [
12], described by several specimens from the Upper Kimmeridgian of Torres Vedras and Lourinhã, and corresponding to the oldest pleurosternid (Paracryptodira) known in Europe [
12], that lineage being also identified in North America; and the thalassochelydian
Hylaeochelys kappa, identified by its holotype (i.e., a single specimen from the middle to upper Tithonian of Mafra), and recognized as the only representative of the genus known in pre-Cretaceous levels [
7]. ‘
Plesiochelys choffati’ Sauvage, 1897–1898 [
13] was defined by a shell, also from the Tithonian of Mafra [
13]. However, it is currently considered as a
nomen dubium, as its holotype and only specimen attributed to it has been reattributed to the thalassochelydian plesiochelyid
Craspedochelys sp. (see [
5], and references therein). The fossil record of Plesiochelyidae is exclusively restricted to Europe, where several representatives are identified. In fact,
Craspedochelys Rütimeyer, 1873 [
14] is not the only plesiochelyid currently identified in the Portuguese fossil record. In this sense, the plesiochelyid
Tropidemys sp. has been identified in the Portuguese record by a single fragment of the central area of a carapace, from the lower Tithonian of Torres Vedras [
15]. In addition, the plesiochelyid
Plesiochelys sp. is currently identified in the Portuguese record by three almost complete shells found in three different Tithonian outcrops of Torres Vedras [
5]. Although the diversity of turtles represented in the Upper Jurassic levels of Portugal includes remains attributable to other forms, their fragmentary state only allows their current identification as Testudinata indet. However, they allow the identification of several additional clades (see [
4], and references therein).
As indicated, the Jurassic record of
Hylaeochelys was, until now, restricted to the holotype of the Portuguese
Hylaeochelys kappa. Two new specimens attributable to
Hylaeochelys, from the same geological formation as that of the holotype (the middle to upper Tithonian Freixial Formation, representing coastal delta plains to fluvial environments), but from different localities in west-central Portugal (
Figure 1), are presented and analyzed here. The new specimens not only show some anatomical regions not preserved in the holotype and so far only known specimen of
Hylaeochelys kappa, but they also display some anatomical differences with respect to it. Its systematic attribution to
Hylaeochelys kappa is discussed, as well as the potential presence of intraspecific variability for the taxon. In this way, the information about this poorly known species is increased.
2. Materials and Methods
The anatomical and systematic study of a relatively complete and articulated shell of a thalassochelydian turtle from the Late Jurassic (middle to late Tithonian of the Freixial Formation, in the Lusitanian Basin) of west-central Portugal (Torres Vedras Municipality, Lisbon District), as well as that of a partial and disjointed shell also attributable to Thalassochelydia, coming from the same geological formation and Portuguese municipality, are performed here. Both specimens are deposited in the vertebrate paleontology collection of the Sociedade de História Natural (Torres Vedras, Portugal). The first one is recognized under the collection number SHN 025 and the other as SHN 026. The precise locality in which the discovery of SHN 025 occurred is Porto Chão Beach (Cambelas, São Pedro da Cadeira Civil Parish). SHN 026 comes from an outcrop located at Zibreira (Carvoeira Civil Parish).
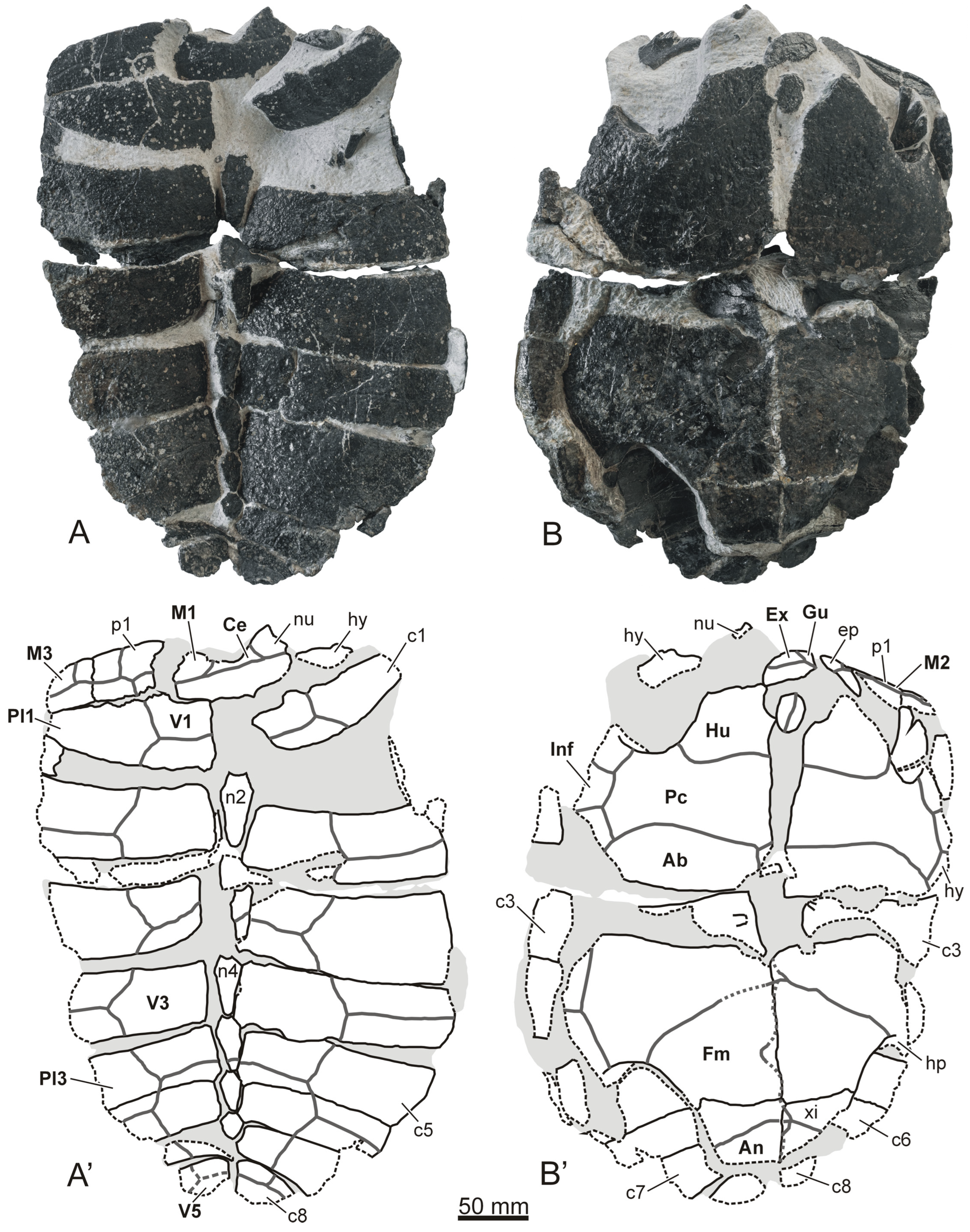
The first-hand analysis of both specimens has been performed. These carapaces have been described in detail. SHN 025 is here represented by photographs of the specimens in five views, i.e., dorsal, ventral, anterior, right lateral, and left lateral views (
Figure 2 and
Figure 3A–C). A selection of elements of SHN 026 are represented in
Figure 3, corresponding to the right hyoplastron, the left third costal, and three neurals (see
Figure 3D–N). Furthermore, details of the outer surface of the carapace of this specimen can also be observed in the same figure, in order to show its ornamental pattern (see
Figure 3G). In addition, schematic drawings of the shell SHN 025 and of the aforementioned plates of SHN 026 are included in these figures. The complete margins of the plates and scutes are represented in these drawings by continuous black lines, and the broken margins of the plates by dotted black lines. The margins of the scutes are represented by thicker gray lines.
The morphological characters recognized in both SHN 025 and SHN 026 are analyzed in detail, so that the systematic attribution of the specimens is performed. They are recognized as attributable to the same representative of Thalassochelydia, i.e., Hylaeochelys kappa, hitherto exclusively known from its holotype. Therefore, new anatomical data on this species are provided, as well as information on its intraspecific variability.
3. Systematic Paleontology
Testudinata Klein, 1760 [
16]
Thalassochelydia Anquetin et al., 2017 [
3]
Hylaeochelys Lydekker, 1889 [
6]
Hylaeochelys kappa Pérez-García and Ortega, 2014 [
7]
New referred material: A relatively complete and articulated shell, SHN 025 (
Figure 2 and
Figure 3A–C); as well as a partial and disjointed shell, SHN 026 (
Figure 3D–N).
Localities and horizon: Both specimens come from the Torres Vedras Municipality (Lisbon District, Lusitanian Basin, west-central Portugal). SHN 025 was found in the Porto Chão Beach (Cambelas, São Pedro da Cadeira Civil Parish). SHN 026 comes from an outcrop in Zibreira (Carvoeira Civil Parish). Freixial Formation, middle to late Tithonian [
17,
18,
19].
Description: The length of the preserved region of the SHN 025 carapace is about 40 cm (
Figure 2 and
Figure 3A–C). The preserved plates of the SHN 026 specimen show that both individuals had a similar length (
Figure 3D–N). The thickness of the plates of both specimens is low. The preservation of SHN 025 does not allow one to characterize its outer surface. However, that of SHN 026 is very well preserved, showing a slightly fluted carapace and plastron surfaces (i.e., showing serial, low parallel ridges) (
Figure 3G). Regardless of taphonomic flattening, the shell SHN 025 is interpreted as low (
Figure 2 and
Figure 3A–C).
SHN 025 shows a nuchal plate that is wider than it is long (
Figure 2A and
Figure 3B). A carapacial notch is present on its anterior margin. It is relatively shallow and, especially, wide, being developed along the entire anterior margin of the plate but restricted to it. The anterior regions of the lateral nuchal margins are slightly divergent posteriorly. This specimen had a continuous neural series, anteriorly contacting the nuchal and posteriorly reaching the seventh pair of costals (
Figure 2A). Thus, the seventh and eighth pairs of costals reach the medial plane. Its neural series was composed of seven elements. The first plate was rectangular and the remainder hexagonal, except the last one, which was pentagonal. All of them are longer than wide. In fact, they are recognized as relatively narrow, as those of SHN 026 are slightly wider (
Figure 3I–N). The costal plates are remarkably wide in relation to their length, so the carapace is interpreted as relatively wide (
Figure 2A). In this sense, the first pair of costals is more than twice as wide as it is long. The first and second peripherals are rectangular, slightly wider than long. Dorsal thickening in the anterior carapace edge is absent in this taxon.
SHN 025 possesses a single and very broad cervical scute, almost five times as wide as it is long (
Figure 2A). Considering the original morphology and arrangement of the plates in the shell (i.e., excluding the modifications generated by the taphonomic processes to which the specimen was subjected), its vertebral scutes are recognized as wide, being noticeably wider than long, especially the most anterior. The first is subrectangular, and the second to fourth vertebrals are hexagonal. The anterolateral margins of the fourth one are notably shorter than the posterolaterals, the margins being more similar in length on the second and third vertebrals. The first vertebral is wider than the nuchal plate. In fact, that vertebral is almost as wide as the fourth. However, the second and third vertebrals are similar to each other and are distinctly the widest.
SHN 026 shows that the connection between the carapace and the plastron was partially osseous (
Figure 3D–E). This specimen shows no evidence of bridge fontanelles. However, it shows a central plastral fontanelle, which is absent in SHN 025 (
Figure 2B). The anterior plastral lobe of SHN 025 is nearly complete, but it is largely disarticulated in situ. It is noticeably wider than long, and slightly less wide than the posterior. The anterior margin of the anterior lobe is subrounded. Its entoplastron is smaller than each epiplastra. It is slightly longer than it is wide. The sutures between the epiplastra and the hyoplastra are approximately straight. The hyoplastra are longer than the hypoplastra. No mesoplastra are present. The lateral margins of the posterior plastral lobe are gently concave laterally.
SHN 025 shows the presence of a pair of gular scutes, with a size like that of the extragulars (
Figure 2B). Posteriorly, the gulars reach the anterior margin of the entoplastron. However, a long distance is present between the extragulars and that plate. The humero-pectoral sulci show a marked anteriorly-directed curve laterally. These sulci show a relatively large distance in relation to the entoplastron. The specimens analyzed possess five inframarginal scutes (
Figure 2B and
Figure 3D). The anal scutes are exclusively developed on the xiphiplastra, the femoro-anal surci being noticeably apart from the suture between the hypoplastra and the xiphiplatra (
Figure 2B). Posteriorly, SHN 025 shows the presence of an at least partially sinuous medial plastral sulcus.
5. Discussion
Until now, the record of
Hylaeochelys in the Iberian Peninsula was restricted to a single specimen, i.e., the holotype of
Hylaeochelys kappa, from the Portuguese Porto Barril Beach (Barril, Encarnação Civil Parish, Mafra Municipality, Lusitanian Basin, west-central Portugal;
Figure 1), found in the middle to upper Tithonian Freixial Formation (see [
7], and references therein). The specimens presented here come from the adjacent municipality of Torres Vedras (
Figure 1), having been found in the same geological formation as the holotype of
Hylaeochelys kappa. The holotype was the only known specimen for this species currently recognized [
4], the other fossils so far attributed to the genus corresponding to
Hylaeochelys belli, identified in the Lower Cretaceous (Berriasian to Valanginian) record of other European countries (United Kingdom and Germany) [
7,
10,
11].
The articulated specimen presented here, SHN 025 (
Figure 2 and
Figure 3A–C), preserves some previously unknown anatomical regions of the shell for the Jurassic record of
Hylaeochelys (restricted to the holotype of
Hylaeochelys kappa). Thus, the morphology of the fourth vertebral scute can be recognized for the first time in the species, as well as the anterior region of the fifth one, but also the medial area of the plastron (showing the absence of a fontanelle), and the hypoplastra and partial xiphiplastra (preserving the femoro-anal sulcus). The disjointed partial shell studied here (i.e., SHN 026;
Figure 3D–N) also allows the assessment of the central region of the plastron, which, by contrast, shows the development of a fontanelle. The presence or absence of a central plastral fontanelle for the adult individuals was a character recognized as variable for the genus
Hylaeochelys, and the absence or different degree of development of this structure was recognized for
Hylaeochelys belli (see Figures 5 and 6 in [
10]), being also a variable character in an intraspecific way for the Portuguese species.
Although SHN 025 does not preserve the first neural, the morphology of the adjacent plates shows that its anterior margin was relatively broad and straight, that of the holotype of
Hylaeochelys kappa being narrow and more distinctly rounded (
Figure 2 and
Figure 3A–C). Although this new specimen SHN 025 shares with the diagnosis of the genus the presence of vertebral scutes that are noticeably wider than long, the width of the vertebrals shows some degree of variability when it is compared with the Portuguese holotype. Thus, the maximum width of the second vertebral of SHN 025 on its right second costal is similar to the length of the sulcus between the first and second pleurals on that plate. However, it is wider than this sulcus in the
Hylaeochelys kappa holotype (see Figure 2 in [
7]). The neural plates of the
Hylaeochelys kappa holotype, especially the anterior and middle ones, are slightly wider than those of SHN 025. In this sense, the neurals of the other specimen analyzed here, SHN 026, are more similar to those of the
Hylaeochelys kappa holotype. Similar variability in both characters can be recognized for the type species of the genus, i.e.,
Hylaeochelys belli (see Figures 5 and 6 in [
10]). The presence of a continuous neural series, composed of seven plates, anteriorly contacting the nuchal, and posteriorly reaching the seventh pair of costals, is shared by the
Hylaeochelys kappa holotype and SHN 025. However, the holotype possesses an additional posterior neural, its neural series being discontinuous. As previously indicated (see [
10], and references therein), the neural series in
Hylaeochelys belli also shows variability, with specimens with a discontinuous neural series and others in which it is continuous, and with a total number of plates that varies between seven and nine elements having been recognized (see Figures 5 and 6 in [
10]). A continuous neural series like that of SHN 025, which meets both the condition of having seven neurals and that the series does not reach the eighth pair of costals, had not been documented until now for this genus. A neural of SHN 026 which, considering its morphology in relation to that of several British
Hylaeochelys specimens (see Figures 5 and 6 in [
10]), as well as the presence of a sulcus on it (interpreted as that between the fourth and fifth vertebrals), is identified as probably the last one (
Figure 3M–N). In this way, the neural series of this specimen would contact the anterior suprapygal posteriorly. The humero-pectoral sulci of SHN 025 are more sinuous than those of the
Hylaeochelys kappa holotype, an intermediate condition being recognized for SHN 026. This is also compatible with the intraspecific variability recognized for the British specimens attributed to
Hylaeochelys belli (see Figures 5 and 6 in [
10]).
Two of the characters used to diagnose the Portuguese Jurassic
Hylaeochelys kappa (exclusively based on its holotype, since it was so far the only Jurassic specimen known for the genus), versus the British and German Lower Cretaceous
Hylaeochelys belli (based on several specimens; see [
10]), are here recognized as variables in the Portuguese material. Thus, the presence of an omega-shaped sulcus between the third and fourth vertebrals, laterally located on the sixth pair of costals and medially on the fifth pair and on the fifth neural, differs from the condition present in SHN 025. The morphology and arrangement of that sulcus in this new specimen falls within the range of variability known for
Hylaeochelys belli (see Figures 5 and 6 in [
10]). In addition, SHN 025 allows us to recognize variability in the gular scutes, not overlapping the anterior margin of the entoplastron. By contrast, an overlap of the gulars of the anterior area of the entoplastron is present in the
Hylaeochelys kappa holotype. Thus, the condition recognized in SHN 025, reaching the posterior margin of the suture between the epiplastra, is compatible with that observed in the only
Hylaeochelys belli specimen for which this region was known (see Figures 5F and 6F in [
10]), so that a slight intraspecific variability for this character is also identified based on the Upper Jurassic material.
The only specimen of
Hylaeochelys belli for which the entoplastron is known shows that the element is relatively large and significantly longer than wide (see Figures 5F and 6F in [
10]). Thus, the distance between the posterior margin of the entoplastron and the humero-pectoral sulcus was characterized, for that species, as less than half of entoplastron length [
7]. The other
Hylaeochelys belli specimens of which the hyoplastron is known allow us to confirm these differences against the
Hylaeochelys kappa holotype. The two new Jurassic specimens from Portugal analyzed here share with the
Hylaeochelys kappa holotype the presence of a relatively long distance between the posterior entoplastral margin and the humero-pectoral sulcus relative to all Lower Cretaceous specimens so far identified for
Hylaeochelys.
6. Conclusions
The European genus of thalassochelydian turtles Hylaeochelys, whose type species was defined in the first half of the 19th century, was, until recently, restricted to the Lower Cretaceous record. Thus, it was exclusively known through the species Hylaeochelys belli, identified in the United Kingdom and Germany fossil records. It corresponded to the only member of the abundant and diverse lineage of Thalassochelydia recognized in the post-Jurassic record. Less than a decade ago, the genus Hylaeochelys was identified in the Upper Jurassic record, by a partial shell from Portugal. Thus, Hylaeochelys is currently the only genus of Thalassochelydia recorded in both the Upper Jurassic and the Lower Cretaceous records.
The analysis of the intraspecific variability of Hylaeochelys belli, and the updating of the knowledge of this form, allowed the attribution of the Portuguese specimen to a new species, Hylaeochelys kappa. However, no other Jurassic remains attributable to Hylaeochelys had so far been documented. Two unpublished specimens from the Upper Jurassic levels of Portugal are analyzed here. Like the Hylaeochelys kappa holotype, they come from the middle to upper Tithonian Freixial Formation, in the Lusitanian Basin. Although they were also found in the Lisbon District (west-central Portugal), they do not come from the Mafra Municipality (where the type locality of the Portuguese species is located) but from the adjacent Torres Vedras Municipality, one of them having been found in the São Pedro da Cadeira Civil Parish, and the other in the Carvoeira Civil Parish. Both are attributed to Hylaeochelys kappa.
The new Portuguese specimens preserve some hitherto unknown anatomical regions for the Jurassic record of the genus Hylaeochelys. As with the Lower Cretaceous specimens attributable to Hylaeochelys belli, the comparison between the three Portuguese specimens allows the recognition of intraspecific variability in the species Hylaeochelys kappa for several characters. In this sense, two characters that were previously proposed in the diagnosis of Hylaeochelys kappa (related to the morphology and arrangement of the sulcus between the third and fourth vertebral scutes, and to the overlapping of the gular scute on the entoplastron), are recognized as variable, showing some character states compatible with those previously reported for some Hylaeochelys belli specimens. The presence of a short entoplastron in the Jurassic material in relation to those of all known specimens for Hylaeochelys belli, as well as the notably longer distance between this plate and the humero-pectoral sulcus in all Portuguese individuals, supports the validity of the species Hylaeochelys kappa. Thus, knowledge about the intraspecific variability of this taxon is increased. The character variability recognized here for Hylaeochelys kappa does not seem to be a result of dimorphism or ontogeny but rather due to individual variability.




{kind=link}
{kind=link}
{kind=link}


