

Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia



Abstract
:1. Introduction
2. Materials and Methods
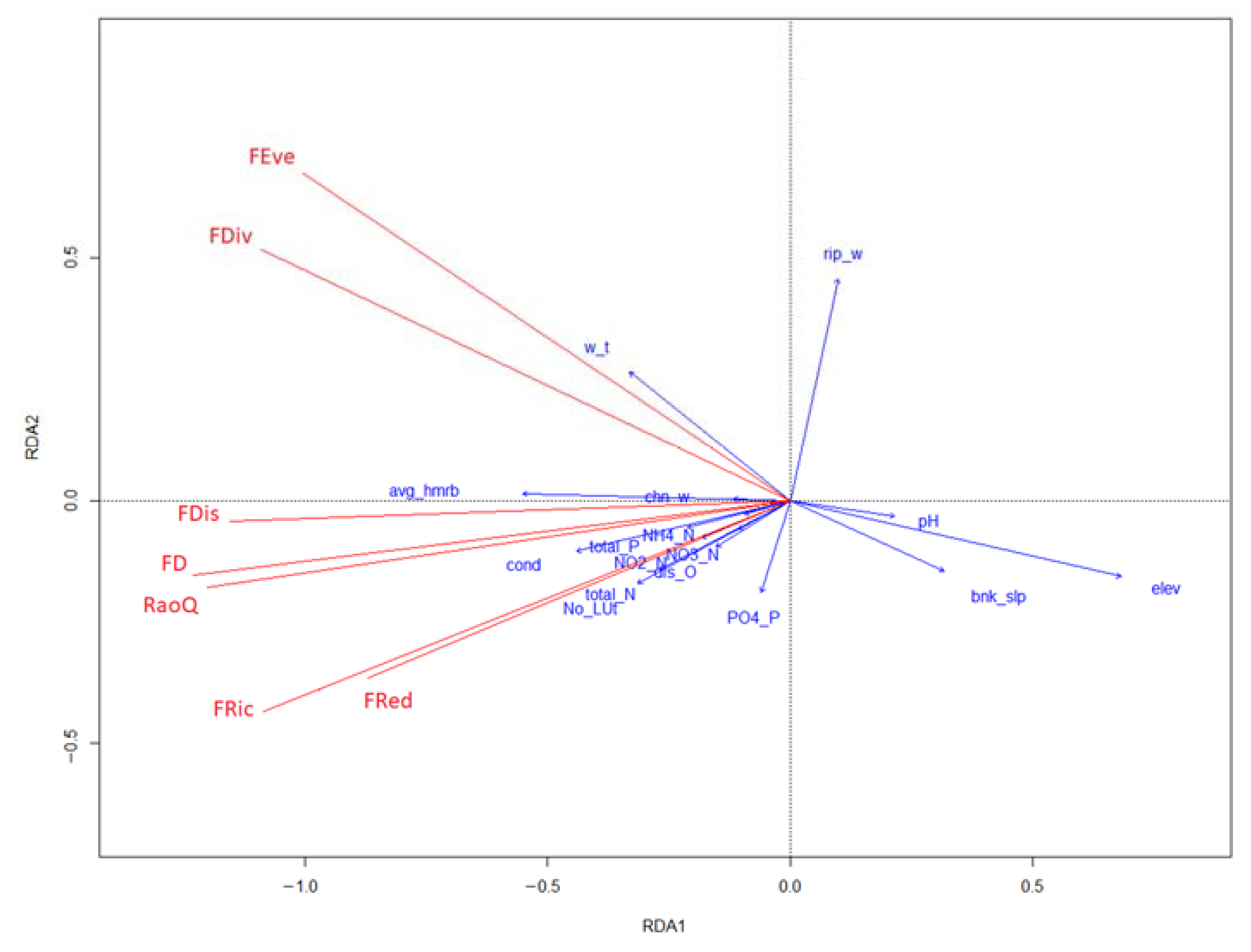
3. Results
4. Discussion
4.1. Correlations between Macrophyte Functional Diversity Indices
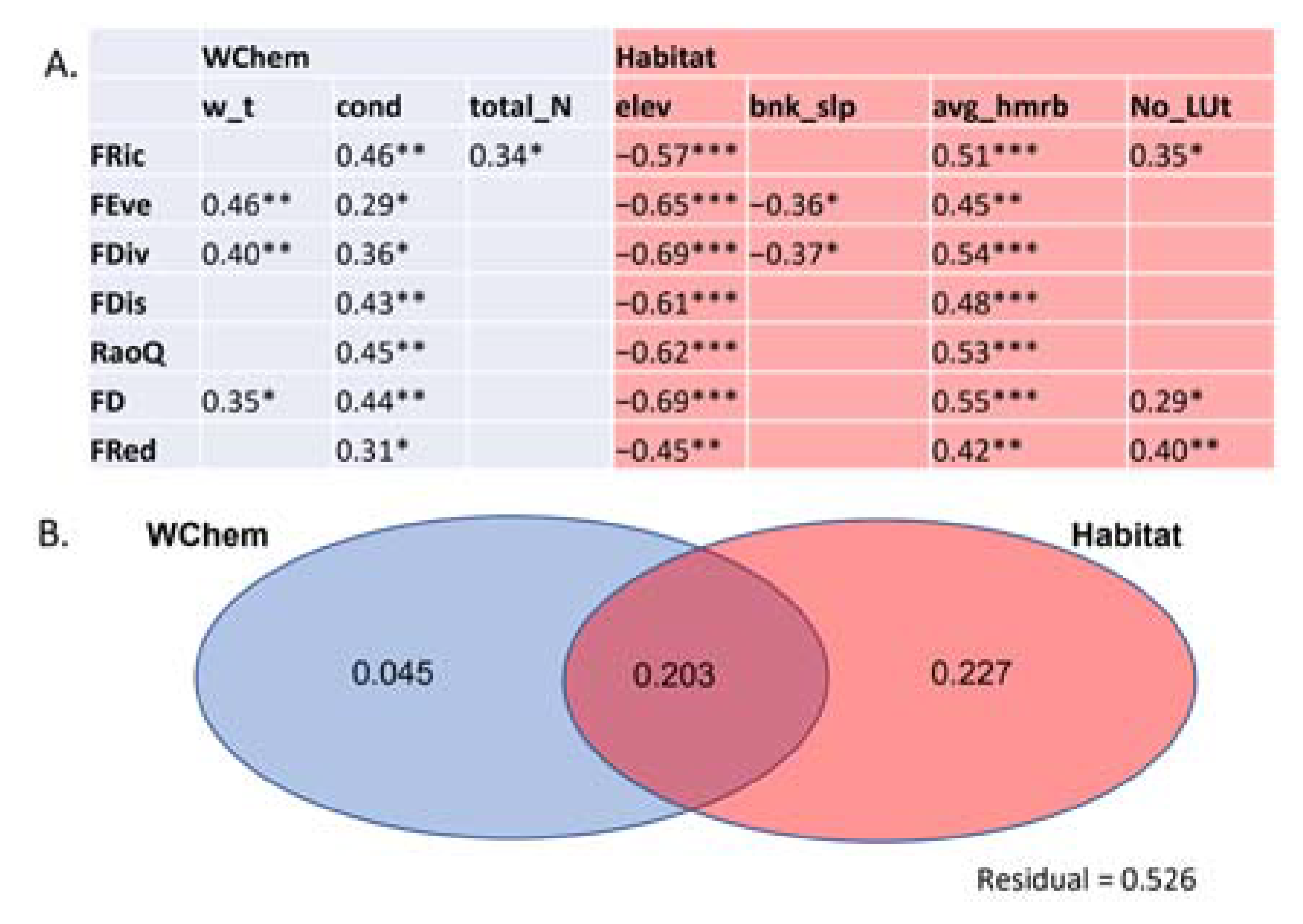
4.2. Effects of Water Chemistry and River Habitat Features on Macrophyte Functional Diversity Indices
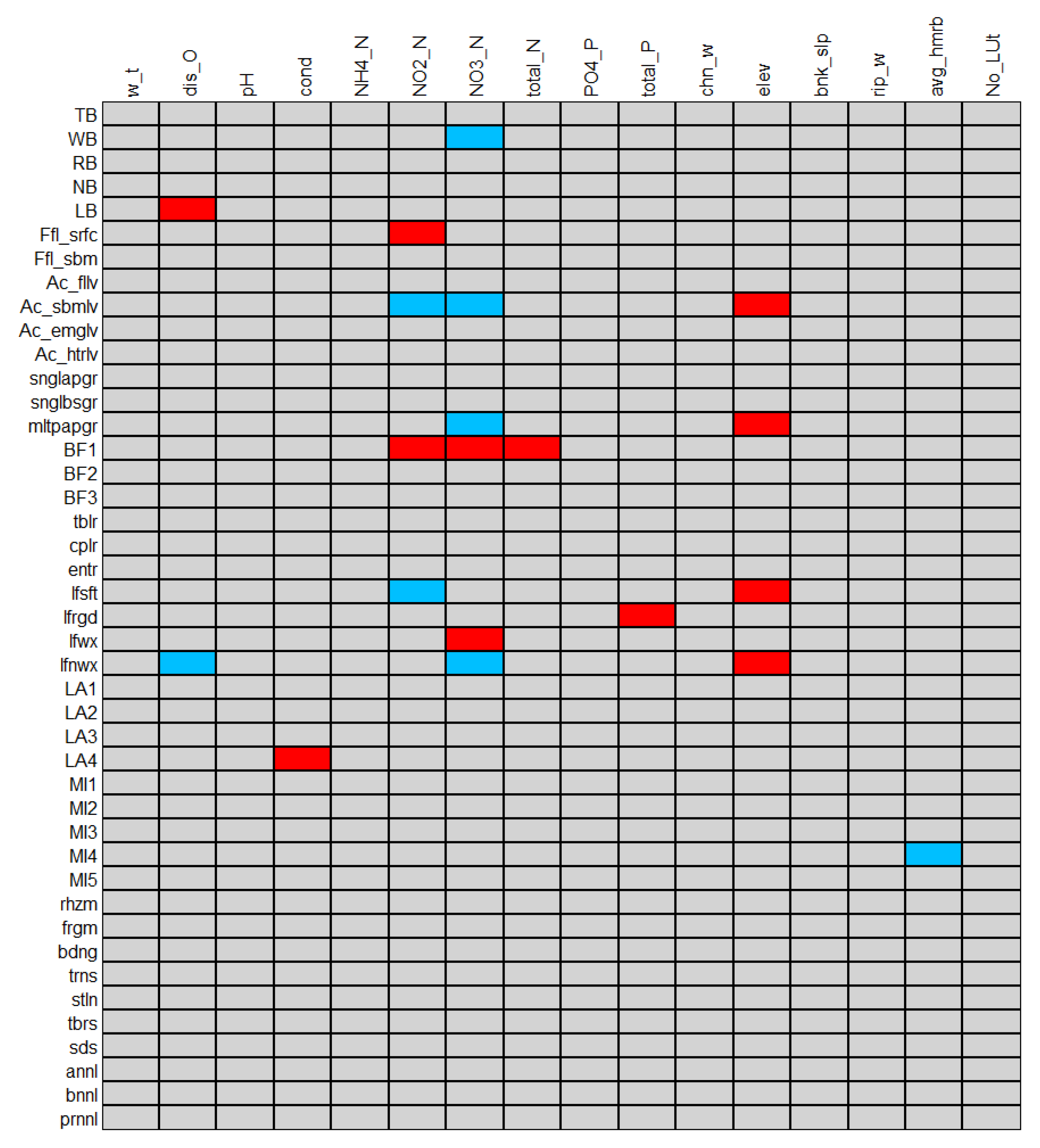
4.3. Effect of Water Chemistry and River Habitat Features on Macrophyte Functional Trait Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGill, B.J.; Enquist, B.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.-L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- Villéger, S.; Miranda, J.R.; Hernández, D.F.; Mouillot, D. Contrasting changes in taxonomic vs. functional diversity of tropical fish communities after habitat degradation. Ecol. Appl. 2010, 20, 1512–1522. [Google Scholar] [CrossRef] [PubMed]
- Baattrup-Pedersen, A.; Göthe, E.; Larsen, S.E.; O’Hare, M.; Birk, S.; Riis, T.; Friberg, N. Plant trait characteristics vary with size and eutrophication in European lowland streams. J. Appl. Ecol. 2015, 52, 1617–1628. [Google Scholar] [CrossRef]
- Lukács, B.A.; E-Vojtkó, A.; Erős, T.; Molnár, V.A.; Szabó, S.; Götzenberger, L. Carbon forms, nutrients and water velocity filter hydrophyte and riverbank species differently: A trait-based study. J. Veg. Sci. 2019, 30, 471–484. [Google Scholar] [CrossRef]
- Paz, L.; Altieri, P.; Ferreira, A.; Ocon, C.; Capítulo, A.R.; Cortelezzi, A. Macrophyte functional traits in channelized streams. Aquat. Bot. 2021, 175, 103434. [Google Scholar] [CrossRef]
- MacArthur, R.; Levins, R. The Limiting Similarity, Convergence, and Divergence of Coexisting Species. Am. Nat. 1967, 101, 377–385. [Google Scholar] [CrossRef]
- Kraft, N.J.B.; Valencia, R.; Ackerly, D.D. Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest. Science 2008, 322, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.W.H.; Lanoiselée, C.; Mouillot, D.; Irz, P.; Argillier, C. Functional characters combined with null models reveal inconsistency in mechanisms of species turnover in lacustrine fish communities. Oecologia 2007, 153, 441–452. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Should Environmental Filtering be Abandoned? Trends Ecol. Evol. 2017, 32, 429–437. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly Rules, Null Models, and Trait Dispersion: New Questions from Old Patterns. Oikos 1995, 74, 159–164. [Google Scholar] [CrossRef]
- Swenson, N.G.; Enquist, B.J. Ecological and evolutionary determinants of a key plant functional trait: Wood density and its community-wide variation across latitude and elevation. Am. J. Bot. 2007, 94, 451–459. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Macrophytes: Ecology of aquatic plants. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Chichester, UK, 2009. [Google Scholar] [CrossRef]
- Manolaki, P.; Guo, K.; Vieira, C.; Papastergiadou, E.; Riis, T. Hydromorphology as a controlling factor of macrophytes assemblage structure and functional traits in the semi-arid European Mediterranean streams. Sci. Total. Environ. 2020, 703, 134658. [Google Scholar] [CrossRef]
- Papastergiadou, E.; Stefanidis, K.; Dörflinger, G.; Giannouris, E.; Kostara, K.; Manolaki, P. Exploring biodiversity in riparian corridors of a Mediterranean island: Plant communities and environmental parameters in Cyprus rivers. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2014, 150, 91–103. [Google Scholar] [CrossRef]
- Göthe, E.; Baattrup-Pedersen, A.; Wiberg-Larsen, P.; Graeber, D.; Kristensen, E.A.; Friberg, N. Environmental and spatial controls of taxonomic versus trait composition of stream biota. Freshw. Biol. 2016, 62, 397–413. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Garssen, A.; Göthe, E.; Hoffmann, C.C.; Oddershede, A.; Riis, T.; van Bodegom, P.; Larsen, S.E.; Soons, M. Structural and functional responses of plant communities to climate change-mediated alterations in the hydrology of riparian areas in temperate Europe. Ecol. Evol. 2018, 8, 4120–4135. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, M.D.; Jansson, R.; Nilsson, C. The effects of hydropeaking on riverine plants: A review. Biol. Rev. 2018, 93, 658–673. [Google Scholar] [CrossRef] [PubMed]
- Directive 2000/60/EC of October 23, 2000. Of the European Parliament and of the council establishing a framework for community action in the field of water policy. Off. J. Eur. Communities 2000, L327, 1–72. [Google Scholar]
- Baattrup-Pedersen, A.; Göthe, E.; Riis, T.; O’Hare, M.T. Functional trait composition of aquatic plants can serve to disentangle multiple interacting stressors in lowland streams. Sci. Total. Environ. 2016, 543, 230–238. [Google Scholar] [CrossRef]
- Stefanidis, K.; Oikonomou, A.; Papastergiadou, E. Responses of different facets of aquatic plant diversity along environmental gradients in Mediterranean streams: Results from rivers of Greece. J. Environ. Manag. 2021, 296, 113307. [Google Scholar] [CrossRef] [PubMed]
- Mouton, T.L.; Matheson, F.E.; Stephenson, F.; Champion, P.D.; Wadhwa, S.; Hamer, M.P.; Catlin, A.; Riis, T. Environmental filtering of native and non-native stream macrophyte assemblages by habitat disturbances in an agricultural landscape. Sci. Total. Environ. 2018, 659, 1370–1381. [Google Scholar] [CrossRef]
- Vukov, D.; Ilić, M.; Ćuk, M.; Radulović, S.; Igić, R.; Janauer, G.A. Combined effects of physical environmental conditions and anthropogenic alterations are associated with macrophyte habitat fragmentation in rivers—Study of the Danube in Serbia. Sci. Total. Environ. 2018, 634, 780–790. [Google Scholar] [CrossRef]
- Dray, S.; Choler, P.; Dolédec, S.; Peres-Neto, P.R.; Thuiller, W.; Pavoine, S.; ter Braak, C.J.F. Combining the fourth-corner and the RLQ methods for assessing trait responses to environmental variation. Ecology 2014, 95, 14–21. [Google Scholar] [CrossRef]
- Vukov, D.; Ilić, M.; Ćuk, M.; Igić, R. The Effect of Hydro-Morphology and Habitat Alterations on the Functional Diversity and Composition of Macrophyte Communities in the Large River. Front. Environ. Sci. 2022, 10, 863508. [Google Scholar] [CrossRef]
- Reich, P.B.; Wright, I.J.; Cavender-Bares, J.; Craine, J.M.; Oleksyn, J.; Westoby, M.; Walters, M.B. The Evolution of Plant Functional Variation: Traits, Spectra, and Strategies. Int. J. Plant Sci. 2003, 164, S143–S164. [Google Scholar] [CrossRef]
- European Standard EN 14184:2014; Water Quality—Guidance Standard for the Surveying of Macrophytes in Running Waters. Comité Européen de Normalisation: Bruxelles, Belgium, 2014.
- Walz, U.; Stein, C. Indicators of hemeroby for the monitoring of landscapes in Germany. J. Nat. Conserv. 2014, 22, 279–289. [Google Scholar] [CrossRef]
- Ellenberg, H. Vegetation Ecology of Central Europe, 4th ed.; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Borhidi, A. Social Behaviour Types of the Hungarian flora, its Naturalness and Relative Ecological Indicator Values, 1st ed.; Janus Pannonius Tudományegyetem Kiadványa: Pécs, Hungary, 1993. [Google Scholar]
- Willby, N.J.; Abernethy, V.J.; Demars, B.O.L. Attribute-based classification of European hydrophytes and its relationship to habitat utilization. Freshw. Biol. 2000, 43, 43–74. [Google Scholar] [CrossRef] [PubMed]
- Botta-Dukát, Z. Rao’s Quadratic Entropy as a Measure of Functional Diversity Based on Multiple Traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R, 2nd ed.; Springer: New York, NY, USA, 2018; pp. 369–412. [Google Scholar]
- De Bello, F.; Lavergne, S.; Meynard, C.N.; Lepš, J.; Thuiller, W. The partitioning of diversity: Showing Theseus a way out of the labyrinth. J. Veg. Sci. 2010, 21, 992–1000. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology. 2022, R package version 1.0-12.1. Available online: https://CRAN.R-project.org/package=FD (accessed on 5 October 2022).
- Ricotta, C.; De Bello, F.; Moretti, M.; Caccianiga, M.S.; Cerabolini, B.E.L.; Pavoine, S. Measuring the functional redundancy of biological communities: A quantitative guide. Methods Ecol. Evol. 2016, 7, 1386–1395. [Google Scholar] [CrossRef]
- Heino, J.; Girón, J.G.; Hämäläinen, H.; Hellsten, S.; Ilmonen, J.; Karjalainen, J.; Mäkinen, T.; Nyholm, K.; Ropponen, J.; Takolander, A.; et al. Assessing the conservation priority of freshwater lake sites based on taxonomic, functional and environmental uniqueness. Divers. Distrib. 2022, 28, 1966–1978. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology 1986, 64, 454–462. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022, R Package Version 2.6-5. Available online: https://github.com/vegandevs/vegan (accessed on 10 October 2022).
- Dolédec, S.; Chessel, D.; ter Braak, C.J.F.; Champely, S. Matching species traits to environmental variables: A new three-table ordination method. Environ. Ecol. Stat. 1996, 3, 143–166. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating Behavior to Habitat: Solutions to the Fourth-corner Problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the species traits–environment relationships: The fourth-corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 613–622. [Google Scholar]
- ter Braak, C.J.F.; Cormont, A.; Dray, S. Improved testing of species traits–environment relationships in the fourth-corner problem. Ecology 2012, 93, 1525–1526. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pebesma, E.J.; Dufour, A.; Thioulouse, J. The ade4 Package—I: One-Table Methods. R News 2004, 4, 5–10. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 14 October 2022).
- Pebesma, E.J.; Dufour, A.; Chessel, D. The ade4 Package—II: Two-Table and K-Table Methods. R News 2007, 7, 47–52. Available online: https://cran.r-project.org/doc/Rnews/ (accessed on 14 October 2022).
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Lacoul, P.; Freedman, B. Relationships between aquatic plants and environmental factors along a steep Himalayan altitudinal gradient. Aquat. Bot. 2006, 84, 3–16. [Google Scholar] [CrossRef]
- Chappuis, E.; Gacia, E.; Ballesteros, E. Environmental factors explaining the distribution and diversity of vascular aquatic macrophytes in a highly heterogeneous Mediterranean region. Aquat. Bot. 2014, 113, 72–82. [Google Scholar] [CrossRef]
- Fernández-Aláez, C.; Fernández-Aláez, M.; García-Criado, F.; García-Girón, J. Environmental drivers of aquatic macrophyte assemblages in ponds along an altitudinal gradient. Hydrobiologia 2016, 812, 79–98. [Google Scholar] [CrossRef]
- Kochjarová, J.; Novikmec, M.; Oťaheľová, H.; Hamerlík, L.; Svitok, M.; Hrivnák, M.; Senko, D.; Bubíková, K.; Matúšová, Z.; Paľove-Balang, P.; et al. Vegetation-Environmental Variable Relationships in Ponds of Various Origins along an Altitudinal Gradient. Pol. J. Environ. Stud. 2017, 26, 1575–1583. [Google Scholar] [CrossRef]
- Stefanidis, K.; Sarika, M.; Papastegiadou, E. Exploring environmental predictors of aquatic macrophytes in water-dependent Natura 2000 sites of high conservation value: Results from a long-term study of macrophytes in Greek lakes. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 29, 1133–1148. [Google Scholar] [CrossRef]
- Alahuhta, J.; Kosten, S.; Akasaka, M.; Auderset, D.; Azzella, M.M.; Bolpagni, R.; Bove, C.P.; Chambers, P.A.; Chappuis, E.; Clayton, J.; et al. Global variation in the beta diversity of lake macrophytes is driven by environmental heterogeneity rather than latitude. J. Biogeogr. 2017, 44, 1758–1769. [Google Scholar] [CrossRef]
- Oikonomou, A.; Stefanidis, K. α- and β-Diversity Patterns of Macrophytes and Freshwater Fishes are Driven by Different Factors and Processes in Lakes of the Unexplored Southern Balkan Biodiversity Hotspot. Water 2020, 12, 1984. [Google Scholar] [CrossRef]
- Kowarik, I. Zum menschlichen Einfluss auf Flora und Vegetation: Theoretische Konzepte und ein Quantifizierungsansatz am Beispiel von Berlin (West). Landsch. Umweltforsch. 1988, 56, 1–280. [Google Scholar]
- Sukopp, H. Dynamik und Konstanz in der Flora der Bundesrepublik Deutschland. Schr. Veg. 1976, 10, 9–26. [Google Scholar]
- Janauer, G.; Exler, N.; Anačkov, G.; Barta, V.; Berczik, Á.; Boža, P.; Dinka, M.; Georgiev, V.; Germ, M.; Holcar, M.; et al. Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity. Water 2021, 13, 918. [Google Scholar] [CrossRef]
- Gutiérrez-Cánovas, C.; Sánchez-Fernández, D.; Velasco, J.; Millán, A.; Bonada, N. Similarity in the difference: Changes in community functional features along natural and anthropogenic stress gradients. Ecology 2015, 96, 2458–2466. [Google Scholar] [CrossRef] [PubMed]
- Sleith, R.S.; Wehr, J.D.; Karol, K.G. Untangling climate and water chemistry to predict changes in freshwater macrophyte distributions. Ecol. Evol. 2018, 8, 2802–2811. [Google Scholar] [CrossRef]
- Lukács, B.A.; Tóthmérész, B.; Borics, G.; Várbíró, G.; Juhász, P.; Kiss, B.; Müller, Z.; G-Tóth, L.; Erős, T. Macrophyte diversity of lakes in the Pannon Ecoregion (Hungary). Limnologica 2015, 53, 74–83. [Google Scholar] [CrossRef]
- Bini, L.M.; Thomaz, S.M.; Murphy, K.J.; Camargo, A.F.M. Aquatic macrophyte distribution in relation to water and sediment conditions in the Itaipu Reservoir, Brazil. Hydrobiologia 1999, 415, 147–154. [Google Scholar] [CrossRef]
- Rolon, A.S.; Lacerda, T.; Maltchik, L.; Guadagnin, D.L. Influence of area, habitat and water chemistry on richness and composition of macrophyte assemblages in southern Brazilian wetlands. J. Veg. Sci. 2008, 19, 221–228. [Google Scholar] [CrossRef]
- Barker, T.; Hatton, K.O.; O’Connor, M.; Connor, L.; Moss, B. Effects of nitrate load on submerged plant biomass and species richness: Results of a mesocosm experiment. Fundam. Appl. Limnol. 2008, 173, 89–100. [Google Scholar] [CrossRef]
- Moss, B.; Jeppesen, E.; Søndergaard, M.; Lauridsen, T.L.; Liu, Z.W. Nitrogen, macrophytes, shallow lakes and nutrient limita-tion: Resolution of a current controversy? Hydrobiologia 2013, 710, 3–21. [Google Scholar] [CrossRef]
- Bornette, G.; Puijalon, S. Response of aquatic plants to abiotic factors: A review. Aquat. Sci. 2011, 73, 1–14. [Google Scholar] [CrossRef]
- Smith, W.S.; Espíndola, E.L.G.; Rocha, O. Environmental gradient in reservoirs of the medium and low Tietê River: Limnological differences through the habitat sequence. Acta Limnol. Bras. 2014, 26, 73–88. [Google Scholar] [CrossRef]
- Scheffer, M.; Szabó, S.; Gragnani, A.; van Nes, E.H.; Rinaldi, S.; Kautsky, N.; Norberg, J.; Roijackers, R.M.M.; Franken, R.J.M. Floating plant dominance as a stable state. Proc. Natl. Acad. Sci. USA 2003, 100, 4040–4045. [Google Scholar] [CrossRef] [PubMed]
- Szabó, S.; Koleszár, G.; Braun, M.; Nagy, Z.; Vicei, T.T.; Peeters, E.T.H.M. Submerged Rootless Macrophytes Sustain a Stable State Against Free-Floating Plants. Ecosystems 2021, 25, 17–29. [Google Scholar] [CrossRef]
- Engelhardt, K.A.M.; Ritchie, M.E. Effects of macrophyte species richness on wetland ecosystem functioning and services. Nature 2001, 411, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Zhong, J.; Yuan, G.; Ni, L.; Xie, P.; Cao, T. Functional traits composition predict macrophytes community productivity along a water depth gradient in a freshwater lake. Ecol. Evol. 2014, 4, 1516–1523. [Google Scholar] [CrossRef]
- Hill, M.J.; White, J.C.; Biggs, J.; Briers, R.A.; Gledhill, D.; Ledger, M.E.; Thornhill, I.; Wood, P.J.; Hassall, C. Local contributions to beta diversity in urban pond networks: Implications for biodiversity conservation and management. Divers. Distrib. 2021, 27, 887–900. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VarPart | Environmental Variable | Unit | Code | Mean | Std.Dev | Min | Median | Max |
|---|---|---|---|---|---|---|---|---|
| Wchem | dissolved oxygen | mg/L | dis_O | 8.55 | 1.55 | 5.21 | 8.93 | 11.29 |
| pH | - | pH | 8.03 | 0.18 | 7.56 | 8.01 | 8.39 | |
| conductivity | µS/cm | cond | 404 | 164 | 126 | 395 | 1234 | |
| ammonium | mg/L | NH4_N | 0.15 | 0.18 | 0.03 | 0.11 | 1.14 | |
| nitrite | mg/L | NO2_N | 0.02 | 0.01 | 0.01 | 0.01 | 0.09 | |
| nitrate | mg/L | NO3_N | 0.70 | 0.34 | 0.23 | 0.66 | 1.53 | |
| total nitrogen | mg/L | total_N | 1.48 | 0.72 | 0.64 | 1.45 | 4.57 | |
| orthophosphate | mg/L | PO4_P | 0.07 | 0.08 | 0.01 | 0.05 | 0.37 | |
| total phosphorus | mg/L | total_P | 0.14 | 0.09 | 0.04 | 0.12 | 0.46 | |
| habitat | water temperature | °C | w_t | 15.08 | 2.16 | 7.95 | 15.35 | 18.79 |
| channel width | m | chn_w | 357 | 364 | 9 | 220 | 1622 | |
| elevation | m a.s.l. | elev | 180 | 185 | 32 | 80 | 832 | |
| bank slope degree | 0–3 | bnk_slp | 1.98 | 0.69 | 1.00 | 2.00 | 3.00 | |
| riparian width | m | rip_w | 116 | 253 | 0 | 5 | 1140 | |
| average degree of hemeroby | 1–7 | avg_hmrb | 3.63 | 1.10 | 1.50 | 3.84 | 6.00 | |
| number of land cover classes | - | No_LUt | 2.80 | 1.44 | 1.00 | 3.00 | 7.00 |
| Trait | Attribute | Code |
|---|---|---|
| Ellenberg Temperature | 1–9 | TB |
| Ellenberg Water | 1–12 | WB |
| Ellenberg Reaction | 1–9 | RB |
| Ellenberg Nitrogen | 1–9 | NB |
| Ellenberg Light | 1–9 | LB |
| Growth form: | free-floating, surface | Ffl_srfc |
| free-floating, submerged | Ffl_sbm | |
| floating leaves, anchored | Ac_fllv | |
| submerged leaves, anchored | Ac_sbmlv | |
| emergent leaves, anchored | Ac_emglv | |
| heterophylly, anchored | Ac_htrlv | |
| Vertical shoot architecture: | single apical growth point | snglapgr |
| single basal growth point | snglbsgr | |
| multiple apical growth point | mltpapgr | |
| Leaf type: | tubular | tblr |
| capillary | cplr | |
| entire | entr | |
| Leaf area: | small (<1 cm2) | LA1 |
| medium (1–20 cm2) | LA2 | |
| large (20–100 cm2) | LA3 | |
| extra-large (>100 cm2) | LA4 | |
| Morphology index (score): | <1 | MI1 |
| 1–10 | MI2 | |
| 10–40 | MI3 | |
| 40–100 | MI4 | |
| >100 | MI5 | |
| Mode of reproduction | rhizome | rhzm |
| fragmentation | frgm | |
| budding | bdng | |
| turions | trns | |
| stolon | stln | |
| tubers | tbrs | |
| seeds | sds | |
| Perennation: | annual | annl |
| biennial | bnnl | |
| perennial | prnnl | |
| Body flexibility: | low (<45°) | BF1 |
| intermediate (45–300°) | BF2 | |
| high (>300°) | BF3 | |
| Leaf texture: | soft | lfsft |
| rigid | lfrgd | |
| waxy | lfwx | |
| non-waxy | lfnwx |
| Index | Abbreviation | Mean | Std.Dev | Min | Median | Max |
|---|---|---|---|---|---|---|
| Functional Richness | FRic | 0.38 | 0.28 | 0 | 0.37 | 0.92 |
| Functional Evenness | FEve | 0.73 | 0.3 | 0 | 0.83 | 0.99 |
| Functional Divergence | FDiv | 0.69 | 0.28 | 0 | 0.79 | 0.88 |
| Functional Dispersion | FDis | 5.63 | 1.12 | 0 | 5.88 | 6.85 |
| Rao Quadratic Entropy | RaoQ | 33.8 | 9.68 | 0 | 35.91 | 47.76 |
| Functional Diversity | FD | 1.37 | 0.12 | 1 | 1.38 | 1.56 |
| Functional Redundancy | FRed | 6.36 | 5.14 | 0 | 4.86 | 19.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukov, D.; Ilić, M.; Ćuk, M.; Igić, R. Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia. Diversity 2023, 15, 231. https://doi.org/10.3390/d15020231
Vukov D, Ilić M, Ćuk M, Igić R. Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia. Diversity. 2023; 15(2):231. https://doi.org/10.3390/d15020231
Chicago/Turabian StyleVukov, Dragana, Miloš Ilić, Mirjana Ćuk, and Ružica Igić. 2023. "Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia" Diversity 15, no. 2: 231. https://doi.org/10.3390/d15020231
APA StyleVukov, D., Ilić, M., Ćuk, M., & Igić, R. (2023). Environmental Drivers of Functional Structure and Diversity of Vascular Macrophyte Assemblages in Altered Waterbodies in Serbia. Diversity, 15(2), 231. https://doi.org/10.3390/d15020231