

Phylogeography and Genetic Diversity of Duck Mussel Anodonta anatina (Bivalvia: Unionidae) in Eurasia

 , , , , , , , ,
, , , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sampling
2.2. DNA Extraction, PCR, and Sequencing
2.3. Phylogeographic Analysis
2.4. Population Genetic Analysis
2.5. Molecular Dating Analysis
2.6. Statistical Analyses
3. Results
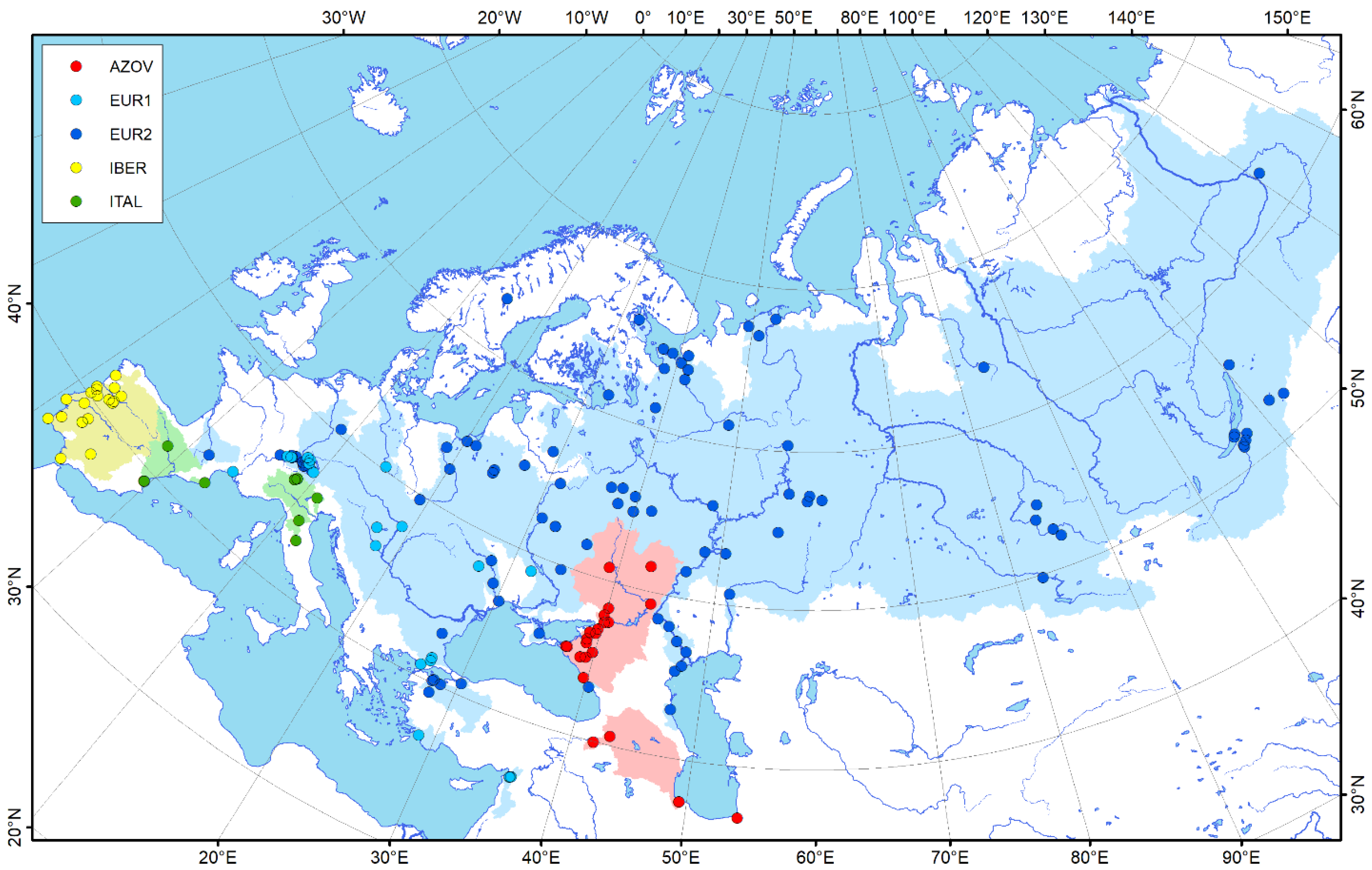
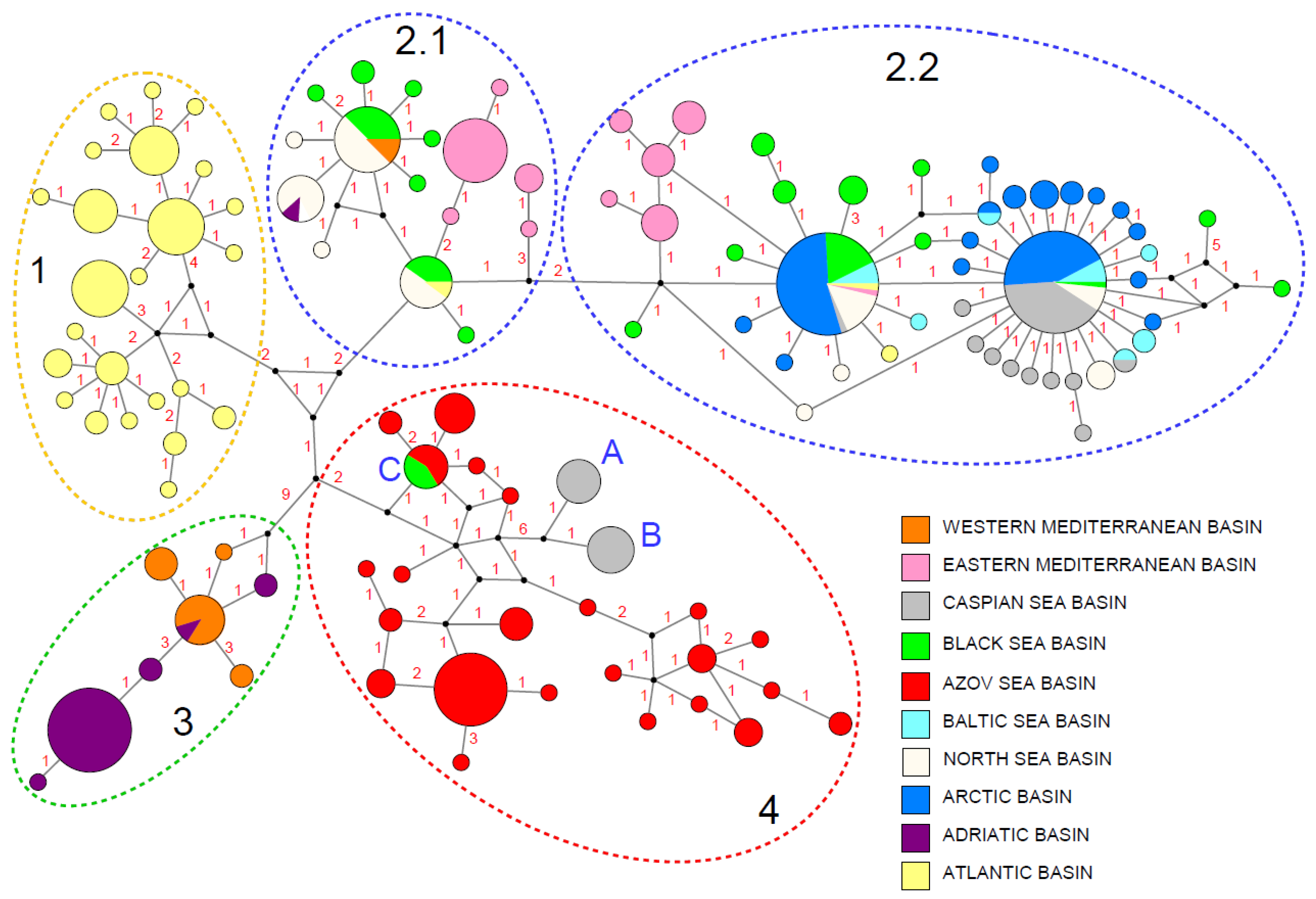
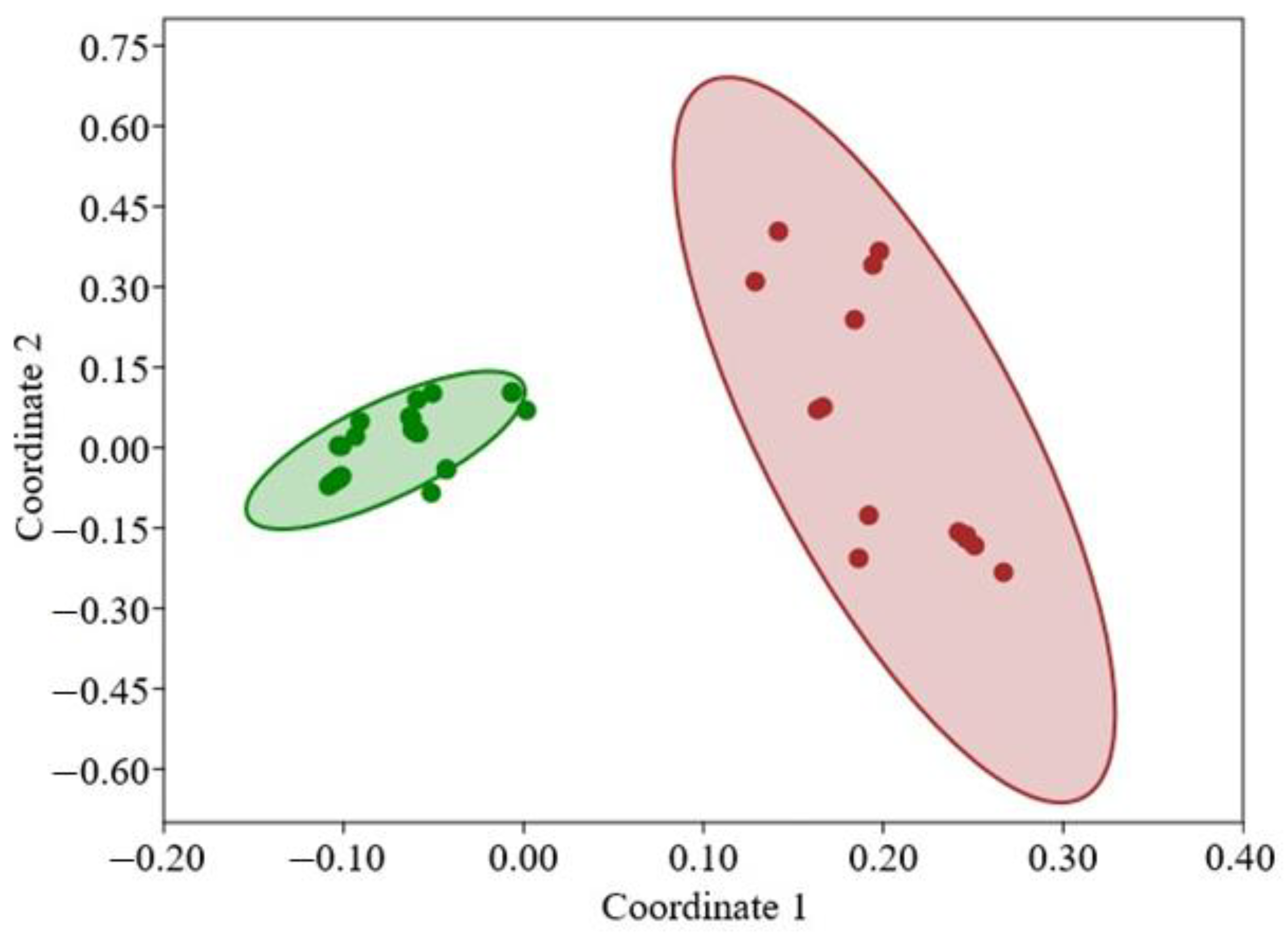
3.1. Phylogeography, Distributional Patterns, and Population Structure
3.2. Genetic Diversity, Population Genetic Indices, and Tests
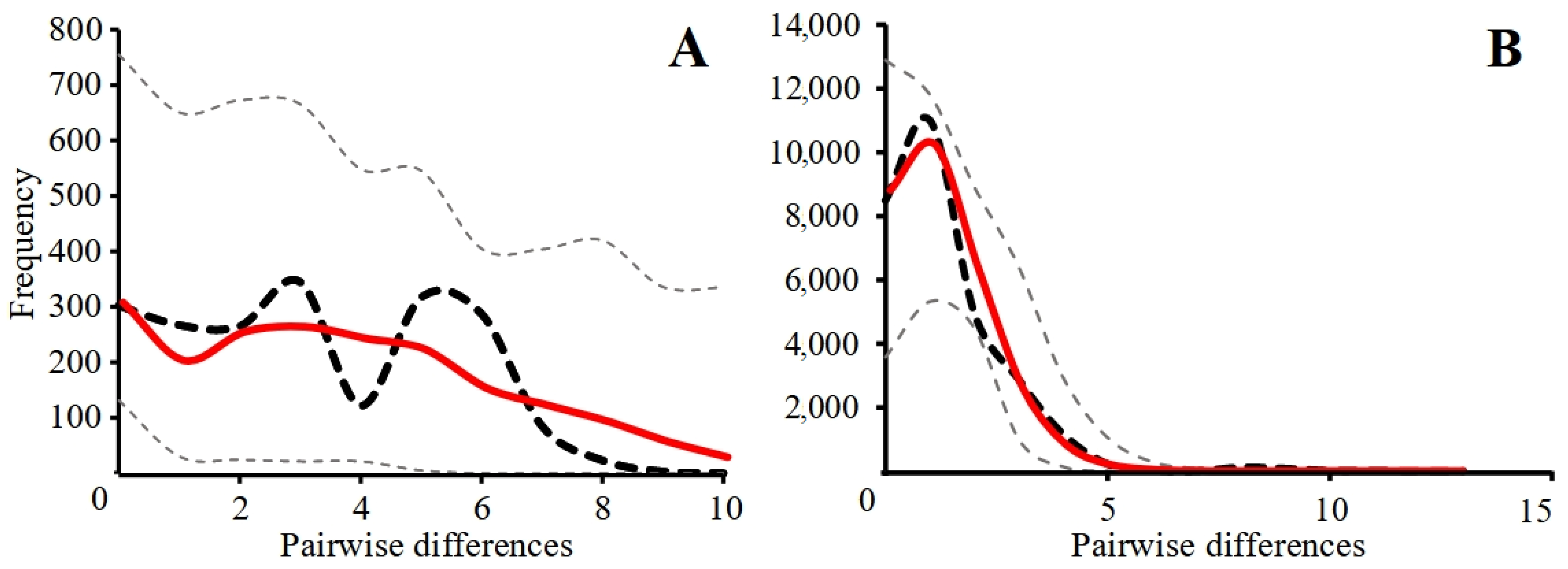
3.2.1. Mismatch Distribution Analysis
3.2.2. Coalescent Simulation
3.2.3. Divergence Time Estimation
4. Discussion
4.1. Distribution Patterns
4.1.1. Interglacial Environment as a Driver of Anodonta anatina Distribution during the Late Pleistocene in Northern Eurasia
4.1.2. Recent Distribution Episodes of the Duck Mussel Induced by Connection between Freshwater basins via Water Channels
4.2. Genetic Diversity and Differentiation
4.3. Conservation Priorities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagel, K.O.; Badino, G.; Alessandria, B. Population genetics of European Anodontinae (Bivalvia: Unionidae). J. Molluscan Stud. 1996, 62, 343–357. [Google Scholar] [CrossRef]
- Graf, D.L. Palearctic freshwater mussel (Mollusca: Bivalvia: Unionoida) diversity and the Comparatory Method as a species concept. Proc. Acad. Nat. Sci. Phila. 2007, 156, 71–88. [Google Scholar] [CrossRef]
- Reis, J.; Machordom, A.; Araujo, R. Morphological and molecular diversity of Unionidae (Mollusca, Bivalvia) from Portugal. Graellsia 2013, 69, 17–36. [Google Scholar] [CrossRef]
- Froufe, E.; Sobral, C.; Teixeira, A.; Sousa, R.; Varandas, S.; Aldridge, D.; Lopes-Lima, M. Genetic diversity of the pan-European freshwater mussel Anodonta anatina (Bivalvia: Unionoida) based on CO1: New phylogenetic insights and implications for conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 561–574. [Google Scholar] [CrossRef]
- Froufe, E.; Lopes-Lima, M.; Riccardi, N.; Zaccara, S.; Vanetti, I.; Lajtner, J.; Teixeira, A.; Varandas, S.; Prié, V.; Zieritz, A.; et al. Lifting the curtain on the freshwater mussel diversity from the Italian Peninsula and Croatian Adriatic coast. Biodivers. Conserv. 2017, 26, 3255–3274. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; Damme, D.V.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2017, 92, 572–607. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Froufe, E.; Ghamizi, M.; Mock, K.E.; Kebapçı, Ü.; Klishko, O.; Kovitvadhi, S.; Kovitvadhi, U.; Paulo, O.S.; Pfeiffer, J.M., III; et al. Phylogeny of the most species-rich freshwater bivalve family (Bivalvia: Unionida: Unionidae): Defining modern subfamilies and tribes. Mol. Phylogenet. Evol. 2017, 106, 174–191. [Google Scholar] [CrossRef]
- Klishko, O.K.; Lopes-Lima, M.; Bogan, A.E.; Matafonov, D.V.; Froufe, E. Morphological and molecular analyses of Anodontinae species (Bivalvia, Unionidae) of Lake Baikal and Transbaikalia. PLoS ONE 2018, 13, e0194944. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Aksenova, O.V.; Aksenov, A.S.; Bespalaya, Y.V.; Borovskoy, A.V.; Danilov, P.P.; Dvoryankin, G.A.; et al. Integrative taxonomy, biogeography and conservation of freshwater mussels (Unionidae) in Russia. Sci. Rep. 2020, 10, 3072. [Google Scholar] [CrossRef]
- Tomilova, A.A.; Lyubas, A.A.; Kondakov, A.V.; Vikhrev, I.V.; Gofarov, M.Y.; Kolosova, Y.S.; Vinarski, M.V.; Palatov, D.M.; Bolotov, I.N. Evidence for plio pleistocene duck mussel refugia in the Azov Sea river basins. Diversity 2020, 12, 118. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Gürlek, M.E.; Kebapçı, Ü.; Şereflişan, H.; Yanık, T.; Mirzajani, A.; Neubert, E.; Prie, V.; Teixeira, A.; Gomes-dos-Santos, A.; et al. Diversity, biogeography, evolutionary relationships, and conservation of Eastern Mediterranean freshwater mussels (Bivalvia: Unionidae). Mol. Phylogenet. Evol. 2021, 163, 107261. [Google Scholar] [CrossRef]
- Gomes-dos-Santos, A.; Froufe, E.; Gonçalves, D.V.; Sousa, R.; Prié, V.; Ghamizi, M.; Benaissa, H.; Varandas, S.; Teixeira, A.; Lopes-Lima, M. Freshwater conservation assessments in (semi-)arid regions: Testing river intermittence and buffer strategies using freshwater mussels (Bivalvia, Unionida) in Morocco. Biol. Conserv. 2019, 236, 420–434. [Google Scholar] [CrossRef]
- Tomilova, A.A. Morphological Variability and Phylogeography of the Duck Mussel Anodonta anatina in Russia and Adjacent Territories. Ph.D. Thesis, Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences, Borok, Russia, 2021. [Google Scholar]
- Tomilova, A.A.; Lyubas, A.A.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Gofarov, M.Y.; Ozcan, T.; Altun, A.; Ozcan, G.; Gürlekd, M.E.; et al. An endemic freshwater mussel species from the Orontes River basin in Turkey and Syria represents duck mussel’s intraspecific lineage: Implications for conservation. Limnologica 2020, 84, 125811. [Google Scholar] [CrossRef]
- Inoue, K.; Monroe, E.M.; Elderkin, C.L.; Berg, D.J. Phylogeographic and Population Genetic Analyses Reveal Pleistocene Isolation Followed by High Gene Flow in a Wide Ranging, but Endangered, Freshwater Mussel. Heredity 2014, 112, 282–290. [Google Scholar] [CrossRef]
- Klunzinger, M.W.; Lopes-Lima, M.; Gomes-dos-Santos, A.; Froufe, E.; Lymbery, A.J.; Kirkendale, L. Phylogeographic study of the West Australian freshwater mussel, Westralunio carteri, uncovers evolutionarily significant units that raise new conservation concerns. Hydrobiologia 2021, 848, 2951–2964. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Lobo, J.; Costa, P.M.; Teixeira, M.A.; Ferreira, M.S.; Costa, M.H.; Costa, F.O. Enhanced primers for amplification of DNA barcodes from a broad range of marine metazoans. BMC Ecol. 2013, 13, 34. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Kondakov, A.V.; Vikhrev, I.V.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kolosova, Y.S.; Konopleva, E.S.; Spitsyn, V.M.; Tanmuangpak, K.; et al. Ancient river inference explains exceptional Oriental freshwater mussel radiations. Sci. Rep. 2017, 7, 2135. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Kallersjo, M.; Von Proschwitz, T.; Lundberg, S.; Eldenas, P.; Erseus, C. Evaluation of ITS rDNA as a complement to mitochondrial gene sequences for phylogenetic studies in freshwater mussels: An example using Unionidae from north-western Europe. Zool. Scr. 2005, 34, 415–424. [Google Scholar] [CrossRef]
- Soroka, M. Identification of gender-associated mitochondrial haplotypes in Anodonta anatina (Bivalvia: Unionidae). Folia Malacol. 2008, 16, 21–26. [Google Scholar] [CrossRef]
- Hinzmann, M.; Lopes-Lima, M.; Teixeira, A.; Varandas, S.; Sousa, R.; Lopes, A.; Froufe, E.; Machado, J. Reproductive cycle and strategy of Anodonta anatina (L. 1758): Notes on hermaphroditism. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2013, 319, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Mezhzherin, S.V.; Yanovich, L.M.; Zhalay, E.I.; Vasilieva, L.A.; Pampura, M.M. Genetic and morphological variability and differentiation of freshwater mussels (Bivavia, Unionidae, Anodontinae) in Ukraine. Vestn. Zool. 2014, 48, 99–110. [Google Scholar] [CrossRef]
- Araujo, R.; Buckley, D.; Nagel, K.O.; Machordom, A. Potomida littoralis (Bivalvia, Unionidae) evolutionary history: Slow evolution or recent speciation? Zool. J. Linn. Soc. 2017, 179, 277–290. [Google Scholar] [CrossRef]
- Araujo, R.; Buckley, D.; Nagel, K.O.; García-Jiménez, R.; Machordom, A. Species boundaries, geographic distribution and evolutionary history of the Western Palaearctic freshwater mussels Unio (Bivalvia: Unionidae). Zool. J. Linn. Soc. 2018, 182, 275–299. [Google Scholar] [CrossRef]
- Pfarrer, B.; Carlevaro, A.; Stucki, P.; Schwarzer, A.; Vicentini, H.; Neubert, E. New phylogenetic insights on some species of Unionidae from Switzerland (Bivalvia, Palaeoheterodonta, Unionidae). Hydrobiologia 2022, 849, 2967–2981. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Feher, Z.; Major, A.; Krizsik, V. Spatial pattern of intraspecific mitochondrial diversity in the Northern Carpathian endemic spring snail, Bythinella pannonica (Frauenfeld, 1865) (Gastropoda: Hydrobiidae). Org. Divers. Evol. 2013, 13, 569–581. [Google Scholar] [CrossRef]
- Bolotov, I.N.; Aksenova, O.V.; Bespalaya, Y.V.; Gofarov, M.Y.; Kondakov, A.V.; Paltser, I.S.; Stefansson, A.; Travina, O.V.; Vinarski, M.V. Origin of a divergent mtDNA lineage of a freshwater snail species, Radix balthica, in Iceland: Cryptic glacial refugia or a postglacial founder event? Hydrobiologia 2017, 787, 73–98. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Wessel, P.; Smith, W.H.F. A global self-consistent, hierarchical, high-resolution shoreline database. J. Geophys. Res. 1996, 101, 8741–8743. [Google Scholar] [CrossRef]
- Lehner, B.; Grill, G. Global river hydrography and network routing: Baseline data and new approaches to study the world’s large river systems. Hydrol. Process. 2013, 27, 2171–2186. [Google Scholar] [CrossRef]
- Lehner, B.; Verdin, K.; Jarvis, A. New global hydrography derived from spaceborne elevation data. Eos 2008, 89, 93–94. [Google Scholar] [CrossRef]
- Elias, S.A. Encyclopedia of Quaternary Science; Elsevier: Amsterdam, The Netherlands, 2006; Volume 1, pp. 2785–2802. [Google Scholar]
- Ehlers, J.; Astakhov, V.; Gibbard, P.L.; Mangerud, J.; Svendsen, J.I. GLACIATIONS: Late Pleistocene in Eurasia. In Encyclopedia of Quaternary Science, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 224–235. [Google Scholar] [CrossRef]
- Markova, A.K.; Puzachenko, A.Y. European small mammal faunas during Dnieper (Saale) glaciation and transition to the Mikulino (Eem) interglacial. Quat. Int. 2021, 605, 254–276. [Google Scholar] [CrossRef]
- Vikhrev, I.V.; Ieshko, E.P.; Kondakov, A.V.; Mugue, N.S.; Bovykina, G.V.; Efremov, D.A.; Bulakhov, A.G.; Tomilova, A.A.; Yunitsyna, O.A.; Bolotov, I.N. Postglacial Expansion Routes and Mitochondrial Genetic Diversification of the Freshwater Pearl Mussel in Europe and North America. Diversity 2022, 14, 477. [Google Scholar] [CrossRef]
- Matoshko, A.V.; Gozhik, P.F.; Danukalova, G.A. Key late Cenozoic fluvial archives of eastern Europe: The Dniester, Dnieper, Don and Volga. Proc. Geol. Assoc. 2004, 115, 141–173. [Google Scholar] [CrossRef]
- Yanina, T.A. The Ponto-Caspian region: Environmental consequences of climate change during the Late Pleistocene. Quat. Int. 2014, 345, 88–99. [Google Scholar] [CrossRef]
- Krijgsman, W.; Tesakov, A.; Yanina, T.; Lazarev, S.; Danukalova, G.; Van Baak, C.G.; Agustí, J.; Alçiçek, M.C.; Aliyeva, E.; Bista, D.; et al. Quaternary time scales for the Pontocaspian domain: Interbasinal connectivity and faunal evolution. Earth-Sci. Rev. 2019, 188, 1–40. [Google Scholar] [CrossRef]
- Bij de Vaate, A.; Jazdzewski, K.; Ketelaars, H.A.; Gollasch, S.; Van der Velde, G. Geographical patterns in range extension of Ponto-Caspian macroinvertebrate species in Europe. Can. J. Fish. Aquat. Sci. 2002, 59, 1159–1174. [Google Scholar] [CrossRef]
- Hwang, S.Y.; Lin, T.P.; Ma, C.S.; Lin, C.L.; Chung, J.D.; Yang, J.C. Postglacial population growth of Cunninghamia konishii (Cupressaceae) inferred from phylogeographical and mismatch analysis of chloroplast DNA variation. Mol. Ecol. 2003, 12, 2689–2695. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Distance, % | ||||||
|---|---|---|---|---|---|---|
| Population pairwise FST | IBER | EUR1 | EUR2 | ITAL | AZOV | |
| IBER | X | 2.25 ± 0.45 | 2.73 ± 0.55 | 3.34 ± 0.63 | 2.56 ± 0.51 | |
| EUR1 | 0.633 | X | 1.39 ± 0.35 | 3.14 ± 0.65 | 2.29 ± 0.50 | |
| EUR2 | 0.833 | 0.735 | X | 3.68 ± 0.74 | 2.44 ± 0.53 | |
| ITAL | 0.765 | 0.833 | 0.924 | X | 3.21 ± 0.64 | |
| AZOV | 0.561 | 0.592 | 0.766 | 0.725 | X | |
| Lineage | N | h | Hd | π | Fu’s Fs | Tajima’s D | Mismatch Analysis (Spatial Expansion Model): Estimated τ |
|---|---|---|---|---|---|---|---|
| IBER | 68 | 24 | 0.912 ± 0.017 | 0.010 ± 0.005 | −5.389 | −0.533 | 6.337 |
| ITAL | 47 | 8 | 0.658 ± 0.066 | 0.004 ± 0.003 | 0.307 | 0.041 | 4.110 |
| AZOV | 80 | 25 | 0.907 ± 0.021 | 0.011 ± 0.006 | −4.064 | 0.017 | 5.939 |
| EUR | 305 | 62 | 0.810 ± 0.019 | 0.006 ± 0.003 | −25.872 * | −1.939 * | 5.452 |
| incl.: | |||||||
| EUR1 | 64 | 16 | 0.850 ± 0.024 | 0.005 ± 0.003 | −3.660 | −0.490 | 2.485 |
| EUR2 | 241 | 46 | 0.706 ± 0.027 | 0.002 ± 0.0015 | −28.793 * | −2.454 * | 0.934 |
| Neutrality Test | Criteria Value | Scenario | |||
|---|---|---|---|---|---|
| Growth | Decline | Bottleneck | Split/Admixture | ||
| Fu’s Fs | −65.184 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Tajima’s D | −2.480 | p < 0.001 | p < 0.001 | p < 0.001 | p < 0.001 |
| Ramos-Onsins and Rozas R2 statistic | 0.014 | p < 0.05 | p < 0.0005 | p < 0.0005 | p < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyubas, A.A.; Tomilova, A.A.; Kondakov, A.V.; Konopleva, E.S.; Vikhrev, I.V.; Gofarov, M.Y.; Eliseeva, T.A.; Aksenova, O.V.; Bovykina, G.V.; Kryuk, D.V.; et al. Phylogeography and Genetic Diversity of Duck Mussel Anodonta anatina (Bivalvia: Unionidae) in Eurasia. Diversity 2023, 15, 260. https://doi.org/10.3390/d15020260
Lyubas AA, Tomilova AA, Kondakov AV, Konopleva ES, Vikhrev IV, Gofarov MY, Eliseeva TA, Aksenova OV, Bovykina GV, Kryuk DV, et al. Phylogeography and Genetic Diversity of Duck Mussel Anodonta anatina (Bivalvia: Unionidae) in Eurasia. Diversity. 2023; 15(2):260. https://doi.org/10.3390/d15020260
Chicago/Turabian StyleLyubas, Artem A., Alena A. Tomilova, Alexander V. Kondakov, Ekaterina S. Konopleva, Ilya V. Vikhrev, Mikhail Yu. Gofarov, Tatyana A. Eliseeva, Olga V. Aksenova, Galina V. Bovykina, Darya V. Kryuk, and et al. 2023. "Phylogeography and Genetic Diversity of Duck Mussel Anodonta anatina (Bivalvia: Unionidae) in Eurasia" Diversity 15, no. 2: 260. https://doi.org/10.3390/d15020260
APA StyleLyubas, A. A., Tomilova, A. A., Kondakov, A. V., Konopleva, E. S., Vikhrev, I. V., Gofarov, M. Y., Eliseeva, T. A., Aksenova, O. V., Bovykina, G. V., Kryuk, D. V., Gorbunova, T. L., Munjiu, O., Pokrovsky, O. S., & Bolotov, I. N. (2023). Phylogeography and Genetic Diversity of Duck Mussel Anodonta anatina (Bivalvia: Unionidae) in Eurasia. Diversity, 15(2), 260. https://doi.org/10.3390/d15020260