

Mate Choice in Molluscs of the Genus Littorina (Gastropoda: Littorinidae) from White Sea

Abstract
:1. Introduction
2. Materials and Methods
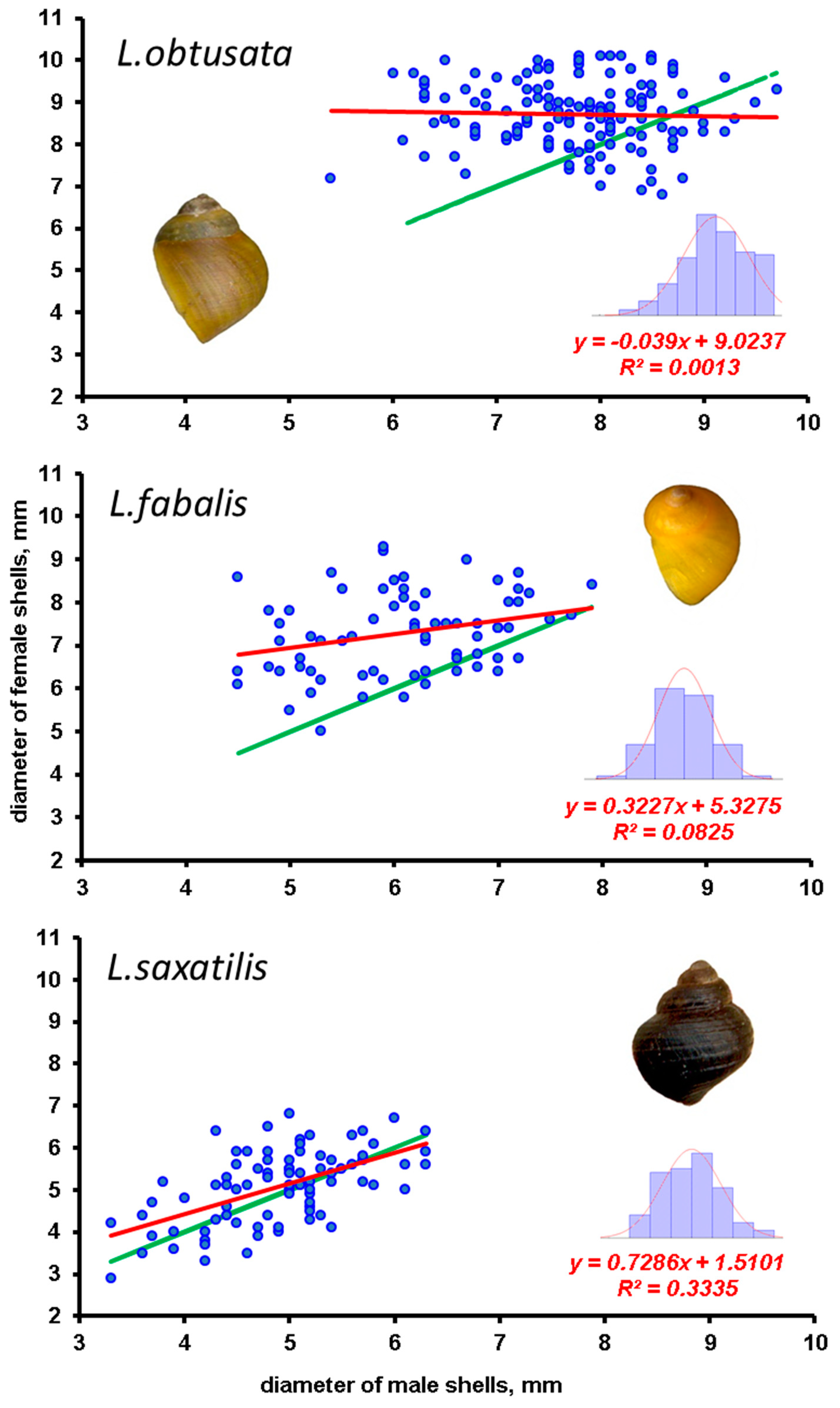
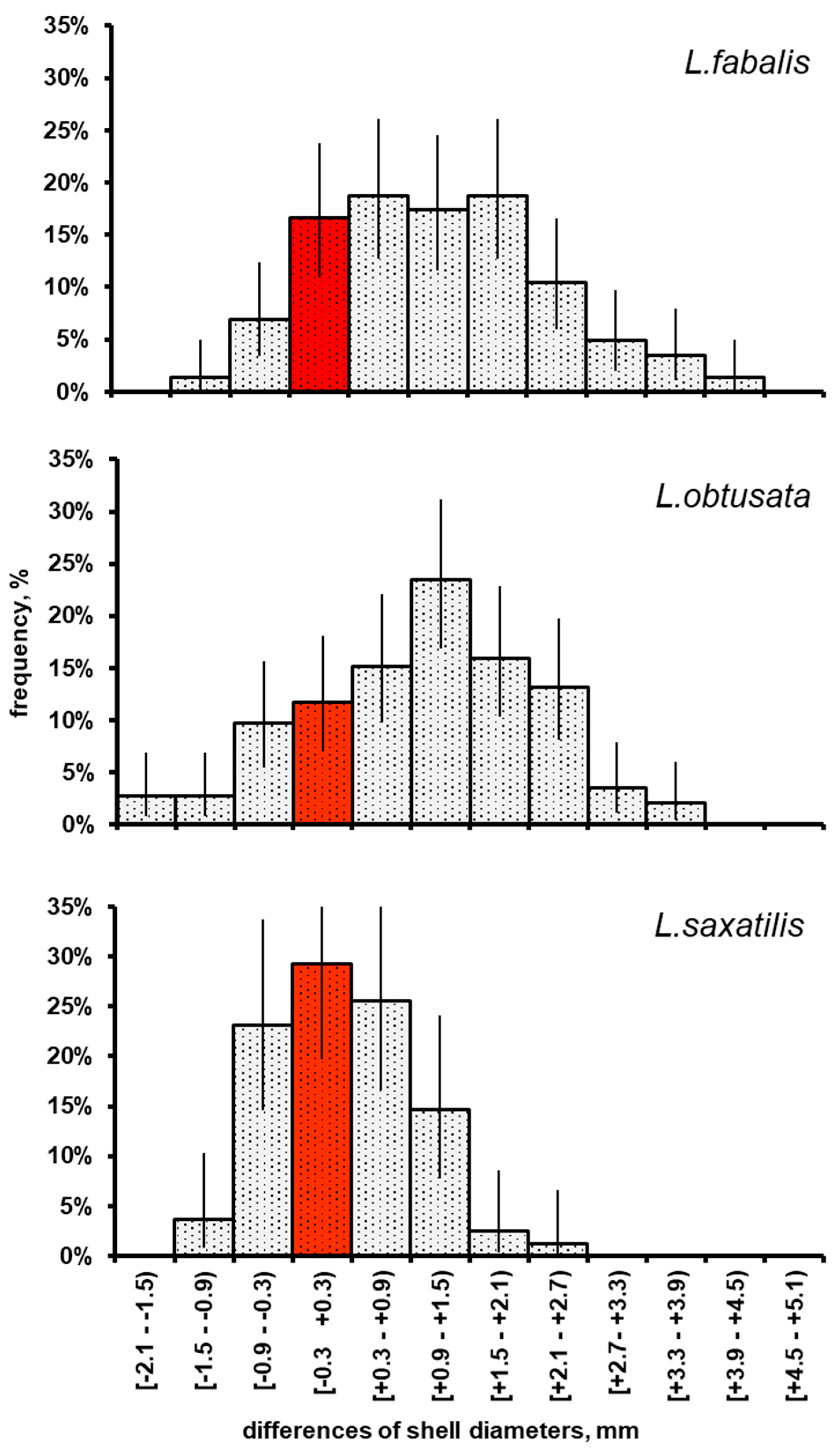
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Coyne, J.A. Genetics and speciation. Nature 1992, 355, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Erlandsson, J.; Rolán-Alvarez, E. Sexual selection and assortative mating by size and their roles in the maintenance of a polymorphism in Swedish Littorina saxatilis populations. Hydrobiologia 1998, 378, 59–69. [Google Scholar] [CrossRef]
- Ng, T.P.T.; Davies, M.S.; Stafford, R.; Williams, G.A. Mucus trail following as a mate-searching strategy in mangrove littorinid snails. Anim. Behav. 2011, 82, 459–465. [Google Scholar] [CrossRef]
- Nagel, L.; Schluter, D. Body size, natural selection, and speciation in sticklebacks. Evolution 1998, 52, 209–218. [Google Scholar] [CrossRef]
- Ratcliffe, L.M.; Grant, P.R. Species recognition in Darwin’s finches (Geospiza, Gould) I. Discrimination by morphological cues. Anim. Behav. 1983, 31, 1139–1153. [Google Scholar] [CrossRef]
- Richmond, J.Q.; Reeder, T.W. Evidence for parallel ecological speciation in scincid lizards of the Eumeces skiltonianus species group (squamata: Scincidae). Evolution 2002, 56, 1498–1513. [Google Scholar] [CrossRef]
- Jones, A.G.; Moore, G.I.; Kvarnemo, C.; Walker, W.D.; Avise, J.C. Sympatric speciation as a consequence of male pregnancy in seahorses. Proc. Nat. Acad. Sci. USA 2003, 100, 6598–6603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, T.P.T.; Williams, G.A. Size-dependent male mate preference and its association with size-assortative mating in a mangrove snail, Littoraria ardouiniana. Ethology 2014, 120, 995–1002. [Google Scholar] [CrossRef]
- Rolán-Alvarez, E.; Austin, C.J.; Boulding, E.G. The contribution of the genus Littorina to the field of evolutionary ecology. Oceanogr. Mar. Biol. Annu. Rev. 2015, 53, 157–214. [Google Scholar]
- Carvalho, J.; Sotelo, G.; Galindo, J.; Faria, R. Genetic characterization of flat periwinkles (Littorinidae) from the Iberian Peninsula reveals interspecific hybridization and different degrees of differentiation. Biol. J. Linn. Soc. 2016, 118, 503–519. [Google Scholar] [CrossRef] [Green Version]
- Johannesson, K.; Saltin, S.H.; Charrier, G.; Ring, A.-K.; Kvarnemo, C.; André, C.; Panova, M. Non-random paternity of offspring in a highly promiscuous marine snail suggests postcopulatory sexual selection. Behav. Ecol. Sociobiol. 2016, 70, 1357–1366. [Google Scholar] [CrossRef]
- Estévez, D.; Kozminsky, E.; Carvajal-Rodríguez, A.; Caballero, A.; Faria, R.; Galindo, J.; Rolán-Alvarez, E. Mate choice contributes to the maintenance of shell color polymorphism in a marine snail via frequency-dependent sexual selection. Front. Mar. Sci. 2020, 7, 614237. [Google Scholar] [CrossRef]
- Hull, S.L. Does size assortative mating maintain a direct polymorphism between two distinct populations of Littorina saxatilis (Olivi) on the north-east coast of England. Hydrobiologia 1998, 378, 79–88. [Google Scholar] [CrossRef]
- Johnson, L.J. Size assortative mating in the marine snail Littorina neglecta. J. Mar. Biol. Assoc. U. K. 1999, 79, 1131–1132. [Google Scholar] [CrossRef]
- Saltin, S.H.; Schade, H.; Johannesson, K. Preference of males for large females causes a partial mating barrier between a large and a small ecotype of Littorina fabalis (W. Turton, 1825). J. Molluscan Stud. 2013, 79, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Galindo, J.; Grahame, J.W. Ecological speciation and the intertidal snail Littorina saxatilis. Adv. Ecol. 2014, 2014, 239251. [Google Scholar] [CrossRef] [Green Version]
- Panova, M.; Hollander, J.; Johannesson, K. Site-specific genetic divergence in parallel hybrid zones suggests nonallopatric evolution of reproductive barriers. Mol. Ecol. 2006, 15, 4021–4031. [Google Scholar] [CrossRef]
- Johannesson, K.; Havenhand, J.N.; Jonsson, P.R.; Lindegarth, M.; Sundin, A.; Hollander, J. Male discrimination of female mucous trails permits assortative mating in a marine snail species. Evolution 2008, 62, 3178–3184. [Google Scholar] [CrossRef]
- Hughes, R.N.; Answer, P. Growth, spawning and trematode infection of Littorina littorea (L.) from an exposed shore in north Wales. J. Molluscan Stud. 1982, 48, 321–330. [Google Scholar] [CrossRef]
- Janson, K. Variation in the occurrence of abnormal embryos in females of the intertidal Gastropod Littorina saxatilis Olivi. J. Molluscan Stud. 1985, 51, 64–68. [Google Scholar] [CrossRef]
- Erlandsson, J.; Johannesson, K. Sexual selection on female size in a marine snail, Littorina littorea (L). J. Exp. Mar. Biol. Ecol. 1994, 181, 145–157. [Google Scholar] [CrossRef]
- Kaufman, Z.S. Osobennosti Polovykh Tsiklov Belomorskikh Bespozvonochnykh (Features of Sexual Cycles of White Sea Invertebrates); Nauka: Leningrad, Russia, 1977; pp. 1–264. (In Russian) [Google Scholar]
- Sergievsky, S.O. Vybor Partnera Dlya Sparivaniya v Populyatsiakh Litoal’nogo Mollyuska Littorina obtusata (L.) (Gastropoda: Prosobranchia) (The Choice of a Partner for Mating in Populations of the Intertidal Mollusc Littorina obtusata (L.) (Gastropoda: Prosobranchia); Vestnik Leningrad State University: Saint Petersburg, Russia, 1983; Volume 21, pp. 22–27. (In Russian) [Google Scholar]
- Maltseva, A.L.; Varfolomeeva, M.A.; Gafarova, E.R.; Panova, M.A.; Mikhailova, N.A.; Granovitch, A.I. Divergence together with microbes: A comparative study of the associated microbiomes in the closely related Littorina species. PLoS ONE 2021, 16, e0260792. [Google Scholar] [CrossRef]
- Kozminsky, E.V. Opredelenie Vozrastsa u Littorina obtusata (Gastropoda, Prosobranchia) (Determination of age in Littorina obtusata (Gastropoda, Prosobranchia)). Zool. Zhurnal 2006, 85, 146–157. (In Russian) [Google Scholar]
- Kuznetzov, V.V. Beloe More I Biologicheskie Osobennosti Yego Flory I Fauny (White Sea and Biological Features of Its Flora and Fauna); AS USSR Publishing: Moscow, Russia, 1960. (In Russian) [Google Scholar]
- Beskupskaja, T.P. Pitanie Nekotoryh Obychnykh Litoral’nykh Bespozvonochnykh Belogo Moray (Nutrition of Some Common Littoral Invertebrates in the White Sea). Proc. Kandalaksha Reserve 1963, 4, 114–135. (In Russian) [Google Scholar]
- Matveeva, T.A. Ecologia I Zhiznennyye Tsikly Obychnykh Vidov Mokkyusk Barentseva I Belogo Morey (Ecology and Life Cycles of Common Gastropod Species in the Barenz and the White Seas). In Seasonal Phenomena in the Life of the White and the Barents Seas; Byhovskiy, B.E., Ed.; Nauka: Leningrad, Russia, 1974; pp. 65–190. (In Russian) [Google Scholar]
- Reid, D.G. Systematics and evolution of Littorina; The Ray Society: London, UK, 1996; 463p. [Google Scholar]
- Glantz, S. Medico-Biologicheskaya Statistica (Primers of Biostatistics); Praktika: Moscow, Russia, 1998; pp. 1–469. (In Russian) [Google Scholar]
- Arnqvist, G.; Rowe, L.; Krupa, J.J.; Sih, A. Assortative mating by size: A meta-analysis of mating pattern in water striders. Evol. Ecology. 1996, 10, 265–284. [Google Scholar] [CrossRef]
- Johannesson, K.; Rolán-Alvarez, E.; Ekendahl, A. Incipient reproductive isolation between two sympatric morphs of the intertidal snail Littorina saxatilis. Evolution 1995, 49, 1180–1190. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Meirama, M.; Estevez, D.; Ng, T.P.; Williams, G.A.; Carvajal-Rodriguez, A.; Rolan-Alvarez, E. A novel method for estimating the strength of positive mating preference by similarity in the wild. Ecol. Evol. 2017, 9, 2883–2893. [Google Scholar] [CrossRef]
- Lloyd, E.; Lederman, W. Spravochnikh po Prikladnoy Statistike (Handbook of Applicable Statistic); Finances and Statistics: Moscow, Russia, 1989; Volume 1, pp. 1–511. (In Russian) [Google Scholar]
- Saur, M. Mate discrimination in Littorina littorea (L.) and L. saxatilis (Olivi) (Mollusca: Prosobranchia). Hydrobiologia 1990, 193, 261–270. [Google Scholar] [CrossRef]
- Zahradnik, T.D.; Lemay, M.A.; Boulding, E.G. Choosy males in a littorinid gastropod: Male Littorina subrotundata prefer large and virgin females. J. Molluscan Stud. 2008, 74, 245–251. [Google Scholar] [CrossRef]
- Yu, T.L.; Wang, L.M. Mate choice and mechanical constraint on size-assortative paring success in a simultaneous hermaphroditic pond snail Radix lagotis (Gastropoda: Pulmonata) on the Tibetan Plateau. Ethology 2013, 119, 738–744. [Google Scholar] [CrossRef]
- Cardoso, R.S.; Costa, D.S.; Loureiro, V.F. Mating behaviour of the marine snail Littoraria flava (Mollusca: Caenogastropoda) on a boulder shore of south-east Brazil. J. Mar. Biol. Assoc. U. K. 2007, 87, 947–952. [Google Scholar] [CrossRef]
- Ng, T.P.T.; Rolan-Alvarez, E.; Dahlen, S.S.; Davies, M.S.; Estevez, D.; Stafford, R.; Williams, G.A. The causal relationship between sexual selection and sexual size dimorphism in marine gastropods. Anim. Behaviour. 2019, 148, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Grahame, J.W.; Wilding, C.S.; Butlin, R.K. Adaptation to a steep environmental gradient and an associated barrier to gene exchange in Littorina saxatilis. Evolution 2006, 60, 268–278. [Google Scholar] [CrossRef] [Green Version]
- Tatarenkov, A.; Johannesson, K. Evidence of a reproductive barrier between two forms of the marine periwinkle Littorina fabalis (Gastropoda). Biol. J. Linn. Soc. 1998, 63, 349–365. [Google Scholar] [CrossRef] [Green Version]
- Granovich, A.I.; Mikhailova, N.A.; Znamenskaya, O.; Petrova, Y.A. Species complex of mollusks of the genus Littorina (Gastropoda, Prosobranchia) from the eastern murman coast. Zool. Zhurnal 2004, 83, 1305–1317. (In Russian) [Google Scholar]
- Ito, A.; Wada, S. Intrasexual copulation and mate discrimination in a population of Nodilittorina radiata (Gastropoda: Littorinidae). J. Ethol. 2006, 4, 45–49. [Google Scholar] [CrossRef]
- Dewitt, T.J. Gender contests in a simultaneous hermaphrodite snail: A size advantage model for behaviour. Anim. Behav. 1996, 51, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Yusa, Y. The effects of body size on mating features in a field population of the hermaphroditic sea hare Aplysia kurodai Baba, 1937 (Gastropoda: Opisthobranchia). J. Molluscan Stud. 1996, 62, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Angeloni, L. Sexual selection in a simultaneous hermaphrodite with hypodermic insemination: Body size, allocation to sexual roles and paternity. Anim. Behav. 2003, 66, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.P.T.; Williams, G.A. Contrasting reproductive traits in two species of mangrove-dwelling littorinid snails in a seasonal tropical habitat. Invertebr. Biol. 2012, 131, 177–186. [Google Scholar] [CrossRef]
- Rolán-Alvarez, E.; Zapata, C.; Alvarez, G. Multilocus heterozygosity and sexual selection in a natural population of the marine snail Littorina mariae (Gastropoda: Prosobranchia). Heredity 1995, 75, 17–25. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Copulating Snails | Non-Copulating Snails | S.S.I. | S.L. | ||
|---|---|---|---|---|---|
| Littorina fabalis | |||||
| Mean | 6.1 | 5.8 | 0.34 | α = 0.0545 | |
| males | S.D. | 0.84 | 0.96 | ||
| N | 72 | 49 | |||
| Mean | 7.3 | 7.4 | −0.10 | do not differ | |
| females | S.D. | 0.94 | 0.94 | ||
| N | 72 | 28 | |||
| Littorina obtusata | |||||
| Mean | 7.8 | 7.8 | 0.00 | do not differ | |
| males | S.D. | 0.78 | 1.01 | ||
| N | 145 | 37 | |||
| Mean | 8.7 | 8.0 | 0.71 | α < 0.001 | |
| females | S.D. | 0.85 | 1.06 | ||
| N | 145 | 40 | |||
| Littorina saxatilis | |||||
| Mean | 4.9 | 4.8 | −0.07 | do not differ | |
| males | S.D. | 0.69 | 0.59 | ||
| N | 82 | 25 | |||
| Mean | 5.1 | 5.1 | −0.01 | do not differ | |
| females | S.D. | 0.87 | 0.71 | ||
| N | 82 | 40 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozminsky, E.V.; Serbina, E.A.; Smurov, A.O. Mate Choice in Molluscs of the Genus Littorina (Gastropoda: Littorinidae) from White Sea. Diversity 2023, 15, 297. https://doi.org/10.3390/d15020297
Kozminsky EV, Serbina EA, Smurov AO. Mate Choice in Molluscs of the Genus Littorina (Gastropoda: Littorinidae) from White Sea. Diversity. 2023; 15(2):297. https://doi.org/10.3390/d15020297
Chicago/Turabian StyleKozminsky, Eugene V., Elena A. Serbina, and Alexey O. Smurov. 2023. "Mate Choice in Molluscs of the Genus Littorina (Gastropoda: Littorinidae) from White Sea" Diversity 15, no. 2: 297. https://doi.org/10.3390/d15020297
APA StyleKozminsky, E. V., Serbina, E. A., & Smurov, A. O. (2023). Mate Choice in Molluscs of the Genus Littorina (Gastropoda: Littorinidae) from White Sea. Diversity, 15(2), 297. https://doi.org/10.3390/d15020297